 2023, Vol. 43
2023, Vol. 43文章信息
- 姚程, 王谦, 姜霞, 郭轶男, 王坤, 吴志皓, 车霏霏, 陈俊伊
- YAO Cheng, WANG Qian, JIANG Xia, GUO Yinan, WANG Kun, WU Zhihao, CHE Feifei, CHEN Junyi
- 湖泊生态系统碳汇特征及其潜在碳中和价值研究
- Review of lake ecosystem's characteristics of carbon sink and potential value on carbon neutrality
- 生态学报. 2023, 43(3): 893-909
- Acta Ecologica Sinica. 2023, 43(3): 893-909
- http://dx.doi.org/10.5846/stxb202112283680
-
文章历史
- 收稿日期: 2021-12-28
- 网络出版日期: 2022-10-10





2. 北京师范大学水科学研究院, 北京 100875;
3. 生态环境部土壤与农业农村生态环境监管技术中心, 北京 100012;
4. 沈阳农业大学水资源学院, 沈阳 110866
2. College of Water Sciences, Beijing Normal University, Beijing 100875, China;
3. Technical Centre for Soil, Agricultural and Rural Ecology and Environment, Ministry of Ecology and Environment, Beijing 100012, China;
4. College of Water Resources, Shenyang Agricultural University, Shenyang 110866, China
全球湖泊总面积约为270万平方公里, 占陆地总面积的1.8%, 发挥着涵养水源、保护生物多样性的生态价值, 拥有着独特的人文景观价值。全球湖泊每年向大气中排放约1100—5700亿吨的CO2, 因积极的呼吸和矿化, 湖泊常处于CO2过饱和状态[1—2], 被认为是典型的碳源[3—4]。湖泊的碳循环强度一般高于草原、森林等陆地单元, 在全球碳循环中扮演者重要角色[5]。图 1概括了碳在湖泊水体-大气、水体-沉积物、湖泊-河道之间的流动和形态变化。湖泊溶解性有机碳(Dissolved Organic Carbon, DOC)可以氧化形成CO2, 溶解性无机碳(Dissolved Inorganic Carbon, DIC)在不同碱度下的平衡决定了水体CO2分压pCO2[6—7]。湖泊沉积物是重要的碳累积场所, 也是湖泊生态系统重要的营养来源与活动场所。湖泊将流域内的碳一部分固定于沉积物中, 一部分以CO2、CH4的形式排放进入大气[8]。湖泊沉积物碳累积情况成为判断湖泊碳流向的重要依据, 长期历史数据反映了不同类型湖泊在气候变化、人类活动影响下碳源/汇角色的切换和变化趋势。

|
| 图 1 湖泊碳循环的概况图 Fig. 1 The overview of carbon cycling in lake DIC: 溶解性无机碳Dissolved Inorganic Carbon; DOC: 溶解性有机碳Dissolved Organic Carbon; TC: 总碳Total Carbon; TOC: 总有机碳Total Organic Carbon; UV: 紫外线Ultraviolet; Alk: 碱度Alkalinity |
根据生态系统对CO2综合吸收/排放情况, 大致可以分为异养型、自养型湖泊[9]。异养型湖泊的生态系统对有机碳的分解利用超过生产者光合作用, 湖泊常处于CO2过饱和状态, 自养型湖泊生态系统以光合作用作为主导, 湖泊处于CO2欠饱和状态, 大气中的CO2向湖泊汇集[10]。初级生产力成为湖泊生态系统发挥固碳功能的重要依据。湖泊富营养化过程中过度营养盐输入让少数几种浮游植物快速繁殖, 健康的湖泊生态系统被破坏, 湖泊生物多样性下降。失去生物多样性, 固碳能力也随之下降。在富营养湖泊出现的CO2欠饱和现象常与浮游植物活动显著相关[11]。有研究估计湖泊每固定1t CO2需要16 pg/m2的净生产力, 往往是富营化湖泊才能达到的净生产力水平[12]。但在富营养化程度严重的湖泊, 例如我国太湖, 初级生产力的提升并不能抵消其CO2排放的增加[13]。同时全球气候变化使得地区平均气温持续上升, 影响湖泊水量、水质的同时加速有机碳的分解。湖泊在具备碳汇潜力的同时面临逐渐严峻的碳排放压力。
随着《巴黎气候协定》的签署实施以及碳达峰、碳中和成为“十四五”乃至2035国家战略目标, 巩固提升自然资源的碳汇能力是践行碳达峰、碳中和的重要保障。目前森林、草原、湿地、海洋、土壤、冻土等地理单元的碳汇价值得到了肯定[14—22], 开展了山水林田湖草沙一体化保护和生态修复[23—33]。但对于湖泊生态系统如何发挥碳汇成效, 碳汇价值如何结算仍在讨论。尤其在全球气候变化、水体富营化这两大背景下现有生态资源碳汇基数存在下降的风险。为了实现湖泊碳汇贡献, 本文梳理对比了全球各类湖泊碳汇/源情况, 旨在提炼提出了未来的可行的方向和途径。
1 湖泊生态系统碳汇特征湖泊生态系统相比森林、草原等陆地生态系统具有固-液两相的复合生境, 相比海洋生态系统体量小, 变化快, 总体上是一类复杂多样的生态系统。湖泊生态系统各层级动植物、微生物都积极参与到湖泊碳循环, 其中以浮游植物为代表的初级生产者光合作用成为了全球不同地区、不同类型湖泊的诸多案例中最为突出且广泛的特征。光合作用虽然受到水体质量、气候温度、地形地貌等条件的影响, 但降低水体pCO2的效果是显著的。光合作用还可以通过影响湖泊碳酸盐体系的平衡来增强自身的碳汇效果。湖泊沉积物中有机碳的提高也是光合作用的重要成果。以鱼类、水生植物为代表的生态系统本身对碳的固定使其成为湖泊又一重要“碳库”。
1.1 水体CO2分压变化浮游植物光合作用降低了水体pCO2, 在部分富营养湖泊出现了与光照强度紧密相关的水体pCO2变化[34], 甚至在少数富营养湖泊中水体无机碳浓度成为限制浮游植物生物量的关键因素, 而不是氮、磷等营养物质[35—36]。水体 pCO2可以指示水体-大气间的CO2交换情况, 40 Pa pCO2被视作大气-水体之间实现了平衡[10, 37—38]。当浮游植物光合作用超过了大气对湖泊水体CO2补给速度, 湖泊处于一种CO2欠饱和状态。全球多个湖泊中出现随着富营养程度提高, CO2欠饱和状态也越频繁[11]。浮游植物生长旺盛期光合作用强度可以达到12.5—50 mmol L-1 h-1, 在数小时内就可以让湖泊内无机碳浓度显著下降[39]。光合作用带来的pCO2下降是有限的。例如在美国Acton湖[40—41]、日本霞浦湖[42—43], 在每年6—10月份 pCO2随着光照强度、Chl.a浓度的增加而持续下降或保持在较低的水平。浅水湖泊相比深水湖泊更易出现pCO2与光合作用的密切联系[44]。在光合作用强烈的富营养湖泊中CO2欠饱和状态是季节性的, 例如巴西Peri湖作为一个富营养浅水湖泊发现在夏季处于CO2欠饱和状态, 在春季处于CO2饱和状态(详见表 1)[45]。制约、影响浮游植物光合作用的因素还有很多, 包括营养水平、水体透明度、气候温度、地形地貌等环境条件。但从全球视角出发, 无论湖泊地处温带还是亚热带, 无论是富营养还是贫营养湖泊, 浮游植物光合作用对降低水体pCO2的积极作用都是不可忽视的, 即使在寒带的湖泊中浮游植物光合作用也可以显著降低水体pCO2, 只是弱于其他碳循环过程[46]。
| 湖泊名称 Lakes |
时间 Time |
平均水深 Depth/m |
营养程度 Eutrophic states |
CO2分压 pCO2/Pa |
初级生产力 Primary productivity/ (mgC m-2 d-1) |
水面碳通量 Carbon flux/ (mmol m-2 d-1) |
沉积物有机碳 Sediment organic carbon/% |
沉积物有机碳累计速率 Sediment organic carbon accumulation rate OCAR/ (gC m-2 a-1) |
碳汇/源 Carbon sink/ source |
湖泊类型 Lake type |
| 洱海Lake Erhai | 2014 | 10.5 | 富营养 | -0.4—0.9 | 3.0—6.0 | 8.0—87.0 | 碳源 | 高原湖泊 | ||
| 滇池Lake Dianchi | 2014 | 5.0 | 富营养 | 0.8 | 10.0—120.0 | 碳汇 | 高原湖泊 | |||
| 武汉东湖Lake Donghu | 2003—2004 | 2.5 | 富营养 | 254.0—2230.0 (NPP) | -0.5—4.2 | 3.53—3.83 | 241.5 | 碳汇 | 亚热带平原湖泊 | |
| 太湖Lake Taihu | 2000—2015 | 1.9 | 超富营养 | 66.3—100.0 | 1088.4(NPP) | 12.4—44.5 | 22.6—99.5 | 5.0—373.0 | 碳源 | 亚热带平原湖泊霞浦湖 |
| Lake Kasumigaura | 1996—2015 | 4.0 | 超富营养 | 0.02—1258.8 | 碳汇 | 温带平原湖泊(日本) | ||||
| Mendota | 2012—2018 | 12.8 | 富营养 | -53.0— -12.6 | 碳汇 | 温带平原湖泊(美国) | ||||
| El sol | 2000—2018 | 6.0 | 贫营养 | 4.6—7.9 | 4.0—110.0 | 碳汇 | 热带高山湖泊(墨西哥) | |||
| La luna | 2000—2018 | 5.0 | 贫营养 | 5.5—6.7 | 19.0—96.0 | 碳汇 | 亚热带高山湖泊(玻利维亚) | |||
| Peri | 2009—2010 | 4.0 | 贫营养 | 26.9(夏)— 47.3(秋) 47.6(冬)— 88.9(春) | -0.16(夏)— 0.11(秋) 0.36(冬)— 0.67(春) | 19.0—77.7 | 季节交替总体碳源 | 热带平原湖(巴西) | ||
| Acton | 2007—2008 | 3.9 | 超富营养 | 5.0—398.8 | 6.1—7.2 (GPP) | -0.1—0.2 | < 1 | 270.0—340.0 | 碳源 | 温带人工水库(美国) |
| Valkea-Kotinen | 1997—1998 | 6.0 | 中营养 | 52.0—76.7 (GPP) | 3.1—5.4 | 36.0 | 3.0 | 碳源 | 温带森林湖泊(芬兰) | |
| Alinen Rautjärvi | 10.0 | 5.5—30.1 (GPP) | 3.6—4.9 | 17.0 | 4.0 | 总体平衡 | ||||
| Ekojärvi | 7.0 | 16.4—54.8 (GPP) | 3.1—5.1 | 11.0 | 6.0 | 碳源 | ||||
| Kuohijärvi | 32.0 | 30.1—71.2 (GPP) | 1.2—1.7 | 4.8 | 1.0 | 碳源 | ||||
| Kukkia | 36.0 | 30.1— 71.2 (GPP) | 1.4—1.8 | 2.0 | 0.3 | 碳源 | ||||
| Geneva | 2000—2010 | 154.0 | 中营养 | 16.0—80.0 | 472.0—826.0 (NPP) | 0.6—2.2 | 13.8 | 碳源 | 温带高山湖泊(法国) | |
| Bourget | 85.0 | 中营养 | 35.2—160.0 | 1416.1—3078.5 (NPP) | 1.5—3.6 | 27.0 | 碳源 | |||
| Annecy | 41.0 | 贫营养 | 40.0—192.0 | 456.8—605.7 (NPP) | 2.8—3.8 | 7.2 | 碳源 | |||
| NPP: 净生产力Net primary productivity; GPP: 总生产力Gross primary productivity | ||||||||||
湖泊的pCO2受水体碱度直接影响。当湖泊存在大量溶解性有机酸、有机阴离子时, pCO2会随着水体碱度一起下降[47]。随着湖泊受纳大量碱性离子、碱性金属, 以及光合-呼吸过程产生的碱度, 水体碱度增加会提高pCO2[48]。微生物呼吸作用会加速HCO3-从湖泊沉积物中释放, 例如在反硝化的过程中会产生OH-, 与CO2反映进一步产生HCO3-[49—50]。湖泊水体碱度的提升有利于大气中的CO2向水体中汇集。初级生产者通过光合作用直接固定大气中CO2的同时, 提高湖泊碱度, 加强大气向湖泊的碳汇通量[12]。浮游植物光合作用过程中对氮、磷、硫等离子的还原, 消耗水中的质子, 水体碱度提升, 甚至改变沉积物中有机碳和CaCO3的比例, 导致沉积物中有机碳的净增量超过CaCO3沉淀的净增量[51]。光合作用引发的水体碱度提升显著加强了CO2向水体的溶解, 并通过浮游植物的同化作用留存在湖泊中。这一现象在初级生产力较高的湖泊中尤为明显, 是湖泊发挥碳汇效益的重要原因[52—54]。
碳酸盐岩的溶蚀也会引起湖泊水体碱度的升高。碳酸盐与Ca2+、Mg2+等阳离子形成的碳酸盐沉淀同时也会发生碳酸盐岩的溶蚀[55]。碳酸盐岩溶蚀涉及一系列链式反应, 是多相化学平衡的复杂溶解过程, 可以总结为以下两个反应[56]:
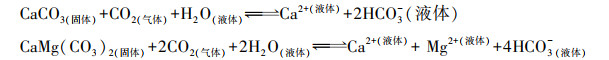
|
反应过程需要从周边环境中吸收消耗CO2。在河道中发现在底层水体发生的碳酸盐岩溶蚀可以促进上层水体中CO2的溶解, 增加大气-水体之间的碳汇通量[56—57]。在微生物作用下发生的碳酸盐沉淀溶蚀现象更有利于湖泊发挥碳汇效益[58]。湖泊中以衣藻、小球藻、蓝藻为代表的浮游植物对碳酸盐岩具有生物溶蚀作用, 通过浮游植物胞外酶以及细胞上的阴离子通道促进碳酸盐岩溶蚀, 产生HCO3-;另一方面阴离子通道会利用HCO3-, 促使沉积物释放更多的溶解性无机碳, 进一步增强湖泊碳汇[59]。碱度提高, pCO2上升, 带来碳汇增益, 这反映了CO2在水体中大量溶解, 积极参与到湖泊碳循环过程。虽然pCO2仍然是判断气-水界面CO2运动趋势的直接依据, 但简单地将pCO2高于40 Pa作为判断湖泊为碳源的依据可能忽视了水体碱度上升带来的碳汇效益。
1.2 沉积物有机碳增加沉积物有机碳是湖泊碳沉积的主要方式[60—61]。有学者估计每年全球湖泊、水库沉积物中要累积大约200兆吨的碳[62], 湖泊中有机碳的累积速率实际上要高于陆地、海洋[63—65]。自20世纪以来湖泊沉积物有机碳累积速率(Organic Carbon Accumulation Rate, OCAR)相比受到工业化影响前的湖泊要高出20%—30%[66—67]。外界营养物质的输入为湖泊带来大量有机碳的同时也促进了湖泊生态系统初级生产力的增长[68], 湖泊沉积物OCAR的加速主要归因于湖泊生态系统光合作用, 通过光合作用固定下的CO2有相当一部分以有机碳的形式留存在沉积物中[69]。浮游植物荚膜吸附裹挟的有机沉淀也会促进沉积物有机碳的累积[70]。湖泊富营养化加速了有机碳在沉积物中的累积。例如在我国云贵高原湖泊随着富营化程度的加剧, 水生植物、浮游浮游植物活动的增强伴随着OCAR的升高[71]。在我国西南地区富营养化程度最高的大型湖泊滇池, OCAR达到112 gC m-2 a-1, 远高于云贵高原大型湖泊平均水平[72]。我国西南大量的深水构造湖泊正由贫/中营养向富营养过度, 再结合深层静水区, 有机碳的累积速率已经高于全国。实际上我国云贵高原大型湖泊(如泸沽湖、洱海、黑海等)年均OCAR约24—27 gC m-2 a-1[72], 远高于太湖(15 gC m-2 a-1)[73]、全球湖泊平均水平(4—14 gC m-2 a-1)[4, 62]。北美明尼苏达州、爱荷华州农业区湖泊沉积物OCAR分别达到50 gC m-2 a-1、200 gC m-2 a-1左右, 远高于该地区的长期历史记录(12—60 gC m-2 a-1)[74]。
在我国云南腾冲青海湖发现沉积物中内源有机碳的比例不断升高, 其中主要来自浮游植物生长代谢[75]。在瑞士卢塞恩这样的贫营养湖泊中异养微生物群落仅能分解少部分(22%)被浮游植物固定下来的碳, 与毗邻的富营养湖泊艾比孔相比沉积物中的有机碳含量也少得多。净初级生产力决定了碳的累积[76]。浮游植物作为湖泊初级生产力的主要构成, 其光合作用是碳汇的第一步, 要实现沉积物有机碳累积的增加, 整个湖泊生态微生物群落的呼吸分解作用也至关重要。光合-呼吸作用之间的平衡、博弈决定了沉积物有机碳的累积情况。当异养微生物群落能及时分解沉降的衰亡组织、有机质时, 沉积物中有机碳不会大量累积;当湖泊经历呼吸对光合的相对滞后, 有机碳才会大量累积。
1.3 食物链对碳的固定在森林开发和保护过程中人们就注意到增加森林生物多样性更有利于碳的累积, 过渡砍伐、商业开发带来的生物多样性下降会引发碳排放的增加[77]。更复杂、更多层次的食物链为生态系统固碳能力打下基础。湖泊生态系统是一个囊括浮游、水生、底栖动植物以及各类细菌、真菌、原生动物的复杂生态系统, 物质、能量在各子系统之间并在子系统内都有着流动和交换, 形成一套复杂的食物链[78]。碳在湖泊生态系统食物链中的流动可以概括为浮游植物、水生植物等初级生产者流向原生动物、浮游动物等初级消费者, 再通向底栖动物、鱼虾等高级消费者, 细菌作为分解者全程参与并将一部分碳归还于大气。一个生物多样性高, 生产力旺盛的湖泊生态系统相比于森林、草原等陆地生态系统更为复杂。在斯堪纳维亚半岛的湖泊中发现初级生产者生产力的提升通过食物链传导给消费者, 初级生产力的提高带动了整个湖泊生态系统生产力的提升, 沉积物中碳的累积也随之提升[79]。在加拿大安大略湖研究者发现象鱼类这样的外来高级消费者入侵都可以引起湖泊生态系统食物链结构变化, 向下传导至浮游植物、浮游动物等初级生产者、消费者, 最终导致整个湖泊生态系统的生产力以及对碳的固定量下降[78]。在我国武汉东湖发现每年通过捕捞淡水鱼能从系统中带走993吨的碳, 而全年以CO2、CH4形式向大气中排放的碳大约在752吨左右[80]。食物链通过影响系统整体的生产力决定了对湖泊生态系统碳的固定能力, 同时也将大量的碳系留其中, 是湖泊发挥碳汇效益的重要“碳库”。
大型水生植物作为湖泊生态系统净生产力的重要组成, 发挥着固定CO2的积极作用[81]。水生植物由发达的根茎、叶子、花卉、果实组成, 通过光合作用将CO2以新增生物量的形式系留在植物组织中, 实际上水生植物的净初级生产力要低于浮游植物, 但其生物量较大, 导致水生植物对碳的累积从时间尺度来看与浮游植物相比缓慢而持续, 后者急促而短暂。研究者认为水生植物相比浮游植物能更有效固定CO2, 不会出现浮游植物死亡后碳的快速释放。植物组织大量纤维素是降解过程中碳缓慢释放的主要原因。纤维素是由葡萄糖组成的大分子多糖聚合物, 马来眼子菜(Potamogeton wrightii Morong)、芦苇(Phragmites australis)等水生植物纤维素在浅水湖泊沉积物中初步降解就需要200天左右, 降解产物也主要由腐殖质构成[82]。木质素是植物组织内含量仅此于纤维素的大分子有机物, 含有丰富的芳环结构以及复杂的碳链结构, 也是最难以分解的那部分有机质[83]。因此丰富的水生植物群落可以加速沉积物中碳的累积, 通过水生植物稳定地将大气CO2系留在湖泊沉积物中。在我国滇西高原纳帕海湿地中以花叶水葱(Scirpus validus)、茭白(Zizania latifolia)、荸荠(Heleocharis liouana)为代表的水生植物生长速率要远高于衰败后的分解速率, 大部以有机质的形式留存在沉积物中, 使纳帕海湿地发挥了“碳汇”的积极作用[84]。在芬兰Valkea—Kotinen、Alinen Rautjärvi、Ekojärvi、Kuohijärvi、Kukkia等湖泊中发现水生植物生物量越大沉积物有机碳总量、OCAR也越高[85]。但在上述湖泊沉积物新增有机碳中水生植物的贡献要低于浮游植物、自然沉降。湖泊水生植物对大气CO2浓度变化还表现出良好的响应, 例如在美国Adirondack山区, 湖泊中的沉水植物群落生物量就与大气CO2浓度呈正相关[86]。在湖泊生态系统中由水生植物固定下的CO2总量相比浮游植物并不大, 但在湖泊碳累积过程中发挥了“压舱石”般的积极作用, 即系统内最为稳定的CO2固定者, 并且一定程度上积极应对大气CO2浓度升高而不受其他因素的影响。
2 湖泊碳排放压力面对当下诸多环境问题, 湖泊碳排放压力陡然增加。例如森林砍伐、农田开垦、城镇发展等土地利用降低了陆上单元的固碳能力, 湖泊面对外部碳源的大幅增加。全球气候变暖加速了水体、沉积物中有机质的分解。富营养化破坏了健康的湖泊生态系统, 导致湖泊生物多样性下降, 浮游植物死亡后还会短时间内释放大量CO2、CH4等气体[87]。湖泊生物多样性下降对湖泊碳汇的负面影响是全面的, 综合生产力下降减少了对碳的同化能力, 食物链中的碳流动也变得简短、急促, 不利于碳的累积。
2.1 外部碳源增加人类活动导致有机碳源的剧增对湖泊碳平衡影响最为明显。例如在我国太湖受周边人口稠密的城镇和现代农业影响CO2释放通量存在显著的空间差异。太湖长年处在CO2饱和状态, pCO2保持在(77.8±16.9) Pa, 平均每年向大气排放(700±300)×106 t的碳[13]。太湖湖面开阔(2400 km2), 深度较浅(1.9—2.5 m), 是典型的大型浅水湖泊, 在富营养化程度较高的太湖西北区域(宜兴沿岸、梅梁湾), 虽然光合作用强度(Chl.a浓度)高, 但该区域年均的pCO2仍然高于全湖, 个别年份甚至超过200 Pa[13]。太湖不同月份的pCO2与Chl.a浓度之间也存在负相关性, 太湖沉积物中以内源型有机碳为主, 来自浮游植物、水生植物沉积的有机碳占据了最高的比重[88]。但对碳的固定速率赶不上生态系统的呼吸分解作用, 太湖并没有出现其他富营养湖泊中呼吸分解作用相对光合同化作用的滞后或落后。
持续汇入湖泊的陆地碳源通过微生物群落的呼吸影响着湖泊的碳平衡, 而碳源类型就成为制约湖泊碳汇功能的客观因素。DOC的输入是导致湖泊的CO2过饱和状态重要原因。DOC在光催化反应下被分解, 在水体中被细菌、浮游植物等微生物降解, 是整个湖泊系统中最易转化为CO2的部分[89—91]。颗粒态有机碳(Particle Organic Carbon, POC)是汇入湖泊的有机碳主要形式之一, 湖泊中包括细菌、昆虫、底栖动物在内的消费者可以直接对其分解利用, CO2作为分解产物进入水体[92]。在一些中小型贫营养湖泊来自陆地的有机碳甚至超过内源有机碳, 成为消费者主要营养来源[93—94]。来自农业用地的易降解有机质在进入沉积物后也更易降解, 进而升高水体pCO2[95]。例如在芬兰众多中小型浅水湖泊中即使低温减缓了水中DOC的分解, 但沉积物有机质的矿化分解作用超越了浮游植物的光合同化作用, 使当地部分湖泊处于CO2过饱和状态[96]。在我国洞庭湖沉积物中来自陆地植物组织的长有机碳链比例不断减少, 随着19世纪中叶至21世纪初包括农业开发在内人类活动的加剧, 短有机碳链比例不断提高, 更有利于有机碳的分解[97]。
2.2 全球气候变暖全球气候变暖对湖泊碳平衡的影响是显著的, 总体上低温有利于有机碳的沉积[98], 而温度升高会导致水体/沉积物中有机碳分解的加剧[99—101]。例如在我国部分深水构造湖中发现随着近十年气候变暖水体DOC的分解也随之加速[102], 虽然温暖的水体、丰富的营养盐让浮游植物大量繁殖, 但其碳的固定速度赶不上有机碳的分解速度[101]。沉积物有机碳的分解也随着温度升高而加强, 无论是有氧条件下发生的氧化分解, 还是缺氧/无氧条件下发生的反硝化、产甲烷反应[100]。例如位于法国阿尔卑斯省的Geneva、Annecy、Bourget湖是典型的构造型深水湖泊(深度82—309 m), 经历了19世纪末至20世纪70、80年代由现代农业引起的水体富营养化, 以及1980s至2000s年代恢复到贫营养的过程, 其水体CO2浓度变化情况也由1980s的平衡甚至欠饱和状态转变为2000s的过饱和状态[103]。1935—1960年3个湖泊均处于富营化早期阶段, 水体CO2浓度达到最高, 湖泊pCO2达到当时大气的2—3倍数;1970s—1980s时期3个湖泊均处于富营养程度最高状态, pCO2出现了短暂的下降, 浮游植物活动的增强对pCO2有着抑制作用。随着2000年后气候变暖加剧, 温度上升带来呼吸、矿化作用加速, 浮游植物光合作用不能抵消呼吸、矿化作用带来的CO2排放, 最终导致碳排放加速[104]。同时随着降雨量的增加入湖河流、雨水冲刷给Geneva、Annecy、Bourget湖泊带来大量DOC, 在微生物群落分解过程中提高了pCO2 [105]。
随着有机碳分解的加快, 湖泊碳平衡可能表现出季节性变化。例如美国威斯康辛州的Mendota湖是一个深受现代农业、城市化影响的大型(39.61平方公里)富营养深水湖泊, 平均深度12.8 m, 最大深度25.3 m, 近年(2012—2017年)监测数据显示Mendota湖发挥着碳汇的作用, 年净碳汇由2012年时的55 g/m2增加至2017年时的204 g/m2, 2014年时最高达到232 g/m2 [106]。实际上Mendota湖在春季至夏季期间是典型的碳源, 平均碳净排放速率可以达到140 g/m2, 进入秋冬季才逐渐转入碳的汇入。北半球湖泊在夏秋之交处于呼吸—光合作用最高时刻[36, 107—108], Mendota湖碳净排放速率在春季至秋季期间不断升高, 在秋季达到最高, 随后整个湖泊迅速转入碳汇状态。在我国洱海也存在夏秋季碳源/汇的交替, 但与Mendota湖不同的是洱海总体上表现为碳源[109]。
2.3 生物多样性下降生物多样性下降影响了生态系统生产力, 生物多样性与生产力的争议由来已久, 有学者认为两者正相关, 物种愈丰富系统的生产力愈高[110], 而有学者认为生物多样性下降会带来生产力升高[111—112]。当生物多样性、生产力还比较低的时候, 随着营养水平的提高, 生物多样性和生产力同时增加, 但当生产力超越一个界限后生物多样性开始下降, 即单峰曲线模式[113]。例如在我国洱海, 富营化带来水生植物生产力提高, 而水生植物群落的多样性却逐年下降, 以苦草(Vallisneria natans)、微齿眼子菜(Potamogeton maackianus)为代表的优势物种生物量主导了洱海水生植物总生物量[114]。在陆地、海洋、淡水生态系统中发挥净水、防灾、产粮等生态功能的主要由系统中2至4种植物或浮游植物承担[115]。在加拿大魁北克地区中营养及以下湖泊中浮游动物生物多样性减少会引起湖泊生产力下降, 但是在富营养湖泊中生物多样性的下降伴随着生产力的上升[116]。
湖泊生态系统物质循环快、生化反应强烈, 有关湖泊生物多样性与生产力之间的研究相对陆地生态系统还有待完善[117]。湖泊生物多样性是生产力提升的前提, 由少数优势物种带来的负相关是短暂而不可持续的[118], 并且在更广阔的空间尺度上生物多样性的增加才能带来丰富的生物量[119]。例如在全美1157淡水湖泊中浮游植物生物量与浮游植物物种多样性就保持了良好的正相关, 由于纬度变化带来的温度、光照差异以及营养状态差异虽然对浮游植物生物量都有影响, 但总体上呈正相关趋势[120—122]。少数几种浮游浮游植物、水生植物生物量快速增加造成整个湖泊生物多样性下降, 这些过度繁殖的植物在衰落死亡过程中伴随着生物降解, 破坏湖泊景观、造成恶劣理化环境的同时向大气中释放包括CO2、CH4在内的温室气体[123]。物种多样性下降会给湖泊碳汇带来全面的负面影响, 尤其由富营化引起的生物多样性下降不仅带来水生植物、浮游植物死亡后CO2、CH4温室气体排放的增加, 中长期对大气中CO2的固定效果也会下降。
3 湖泊碳中和潜力为了发挥湖泊生态系统碳汇潜力, 以提高生物多样性为目的的湖泊生态修复, 以正确认识沉积物碳累积贡献的评价方法改进, 以减排产能为目的的技术探索可能是未来的重点和难点。初级生产者对CO2的同化是湖泊生态系统发挥碳汇价值的第一步, 通过生态修复提高湖泊生物多样性, 延长食物链中的碳流动, 增强碳在湖泊中的系留效果。调查沉积物中碳的累积情况为我们提供一个判断湖泊碳平衡的高效手段, 综合反映了湖泊中碳的累积速率、来源, 有机碳的分解程度, 通过与陆地碳累积情况对比突出湖泊生态系统的碳汇贡献。在湖泊碳沉积资源化的探索中沉积物产CH4的技术路线取得了初步成果。
3.1 改善湖泊生态系统重建水生植物群落是湖泊生态修复的重要手段, 过程中在局部水域或沿岸人工培植了大量挺水、浮叶、沉水植物, 初步达成了净化水质、控制氮磷、恢复湖泊生物多样性的目的[124—126]。随着湖泊生态修复实践的增加, 研究者提出合理地构建水生植物群落可能有助于控制湖泊碳排放, 例如在白洋淀增加水生植物覆盖面积可以作为抵消引水扩大湖面、碳排放增加的有效手段[127];在大通湖恢复的沉水植物群落不但通过光合作用将CO2以有机质的形式固定在沉积物中, 还可以通过影响根区微生物加速有机质的累积[128]。在日本八郎湖大量培植的挺水植物群落虽然没能显著地提高沉积物有机碳含量, 却改善了沉积物有机质的构成, 抑制了有机碳在夏季的快速分解释放[129]。研究者对日本八郎湖挺水植物没能显著提高沉积物有机碳含量特意做了解释和分析, 主要原因包括分析方法的局限性可能会遗漏挺水衰亡过程中的细小有机组织, 挺水植物通过减少风浪为当地细小无机颗粒物大量沉积提供了有利的水力条件。水生植物修复为湖泊带来的直接碳汇效益也许是有限的, 但水生植物对湖泊生态系统和生境的改善是显著的, 从氮、磷营养盐控制[130—132]到抑制基底再悬浮[133], 从改变微生物群落结构[128]到增加沉积物中的自由氧[134]。这些改变可以提高水体透明度, 增强光合作用, 可以提供沉积物氧化环境, 有利于沉积物有机碳的稳定累积, 为湖泊碳汇效益“保驾护航”。
提高生物多样性是增强生态系统碳汇效益的可行途径。在巴西大西洋热带雨林保护项目中就将减少碳排放、增加森林碳汇同恢复植被覆盖、恢复雨林生物多样性一道设置为首要目标[135]。在芬兰南方开展了恢复泥炭湿地结构与功能的生态修复项目, 该泥炭湿地随着水土流失正在失去植被覆盖度、生物多样性以及固碳能力, 通过人工培植陆上植物首先克服了水土流失导致的生态退化, 恢复了泥炭湿地部分固碳能力。但对比未受人类影响的自然泥炭湿地, 人工修复湿地的固碳速率还远远落后, 人工修复生态系统的生物多样性还有所缺失, 导致碳汇效益的大打折扣[136]。在诸多湖泊生态修复案例中从浮游植物[137]到大型水生植物[138], 从浮游动物[139]到底栖动物[140], 从鱼类[141]到水鸟[142], 湖泊生物多样性得到了恢复或改善。当以浮游植物、水生植物为代表的湖泊初级生产者生物多样性提高时, 可以显著地改变食物链中的碳流动, 将更多的碳系留在湖泊中[143]。通过湖泊生态修复, 提高生物多样性, 虽然能将更多的碳固定下来, 但可能要面临食物链中碳流动简短, 整体效率不如原生态系统的缺点。
3.2 重视沉积物碳累积湖泊沉积物是浮游植物、水生植物衰亡后, 以及大量入湖有机物的最终去向[144—146]。相比水体中溶解性有机物的矿化分解, 沉积物中的反应速度普遍较慢[147—150]。单独依据pCO2来判断湖泊的碳汇/源可能会忽视湖泊生态系统的碳汇贡献。部分CO2过饱和湖泊同时拥有着较高的沉积物OCAR, CO2在向大气释放的同时有机碳也在沉积物中快速累积。位于武汉附近的东湖作为一个中/富营养湖泊常年处于CO2过饱和状态, 初级生产力约1440 mgC m-2 d-1, 沉积物碳累积率达到241.5 gC m-2 a-1, 通过对比大气-水体间碳通量、沉积物碳累积量, 发现东湖每年向大气碳排放量仅占沉积物碳累积量8%, 发达的湖泊生态系统让东湖发挥了碳汇的积极作用[80]。位于长江下游的太湖常年处于CO2饱和状态, 周期暴发的蓝藻在衰亡后还会堆积腐解以甲烷CH4的形式排入大气[151]。太湖沉积物OCAR可以达到373.0 gC m-2 a-1[73], 太湖沉积物有机质的来源在湖区之间各有不同, 以梅梁湾、竺山湾为代表的太湖西北湖区由于富营化导致沉积物中含有大量浮游植物有机质, 以太浦运河河口为代表的太湖东南湖区由于丰富的水生植物使得沉积物中也以水生植物有机质为主, 以贡湖湾为代表的太湖中部、东北部以陆地植物有机质为主, 三者有机质易分解程度由易到难[152]。虽然基于水体pCO2、沉积物OCAR, 研究者分别得出了太湖作为碳源、碳汇单元的相左结论, 但两者都客观反映了太湖碳循环的一部分, 即太湖在向大气排放CO2、CH4的同时也在受纳流域内陆地有机碳[13, 73]。要判断太湖的碳源/汇状况需要完整、全面的碳平衡结算。
湖泊液相环境比陆地单元能更快地分解溶解性有机质[153], 但还有一部分来自流域内陆地单元的有机质被固定在沉积物中[154—157]。这部分碳如果没有进入湖泊生态系统而是停留在陆地上是否会减少碳排放, 将深刻影响湖泊碳汇价值判断。通过研究木质素形态发现来自森林的陆地有机质在进入湖泊前已经高度降解, 进入湖泊后反而在沉积物中稳定了下来[158]。我国东部平原湖泊的平均碳累积速率、有机碳含量要高于其他区域的湖泊, 而东部地区土壤中有机碳含量要低于其他区域。这种差异主要归因于有机质在陆上的矿化分解速度比在湖泊沉积物中更剧烈。东部平原丰富的植被、暖湿的气候以及更多的人类活动使碳循环更加迅速, 有机质在土壤中的分解要强于在湖泊沉积物中的分解[159]。面对流域内的陆地碳源, 如果一个湖泊每年沉积物碳的累积速率超过流域内土壤碳累积速率, 该湖泊的整体碳汇应该得到肯定。全面的碳平衡结算是评价湖泊碳源/汇贡献的最可靠、最理想的方法, 但需要长期的、涉及多项理化、生物指标的全面调查, 成本高耗时长。湖泊沉积物除了可以直接反映碳累积速率变化趋势外还可以通过有机碳的溯源、分类反映湖泊长期的营养、生产力水平, 调查沉积物可以同时了解以上影响湖泊碳循环的关键因素。重视沉积物碳累积, 加强沉积物有机碳的溯源分类将为我们对湖泊的碳汇价值判断提高重要参考, 同时也给出了替代碳平衡全面调查的可靠方案。
3.3 采集湖泊甲烷资源在温室气体排放、能源需求的双重压力下采集利用环境CH4气体的方案被提出, 以期减少温室气体排放的同时用于燃烧发电, 尽可能减少能源供应对化石燃料的依赖。CH4作为一种温室效益数十倍(28—36)于CO2的温室气体在人类进入工业化的百年间迅速上升[160]。湖泊CH4气体主要源自底层水体、沉积物缺氧条件下产甲烷菌对有机质的分解, 全球湖泊、水库在2011年向大气排放约92×106—142×106 t的CH4, 大约占当年环境排放总量的20%[161]。湖泊、湿地、水库等内陆淡水水系所产CH4被选为潜在的替代能源除了目前可观的产量外, 易获取以及预期增长的产量成为重要的依据。易获取性指的是湖泊、水库往往靠近城镇基础设施, 开发成本低, 湖泊、水库的水力条件相对稳定, 利于采集运输[162]。预期增长的产量主要源于目前通过测算表面水体CH4通量低估了湖泊的CH4产量。有研究估计有相当一部分(30%—90%)来自底层水体、沉积物的CH4向上方释放过程中被氧化, 如果能减少氧化损失, 理论上可以获取约470×106 t的CH4[163]。另一方面CH4主要来自水生植物、浮游植物的衰亡腐解, 以及有机沉积的生物分解, 水体的富营养化以及入湖有机碳的增多势必引起CH4产出持续升高, 带来碳排放压力的同时却为开辟湖泊碳中和新局面提供了契机。
对于开发利用湖泊CH4气体的技术前提和难点是对其进行富集。早在2008年东非基伍湖(Lake Kivu)就已经开展湖泊CH4气体的采集, 并用于当地电力供应[164]。但该湖泊深达200米深水静置层高浓度CH4极为独特, 一般湖泊尤其遍布我国东部地区的浅水湖泊面临CH4总量大、浓度低的不利局面, 再加上CH4气体本身较低的能量密度, 进一步加剧了应用难度。常用的低温高压液化处理方式运行成本高, 尤其对于湖泊中低浓度的CH4气体该方法基本失去了应用价值[165]。疏水气-液膜反应器(Hydrophobic Gas-Liquid Membrane Contactors, GLMC)在废水处理领域已经实现了气-液两相的高效分离(>50%), 发展出微孔聚合膜(microporous polymer-based membranes)、中空纤维超滤膜(hollow-fiber ultrafiltration membranes)等成熟技术[166—167]。相关领域一方面改变膜的微观构型, 一方面不断利用高分子聚合材料增强膜的疏水性能以及结构稳定性, 以期可以富集更低环境浓度的CH4气体, 更长的使用寿命[168—169]。同时以酶膜反应器为代表的新技术尝试将CH4气体转化为甲醇CH3OH。该技术的核心在于将甲基球菌(Methylococcus capsulatus)的甲烷单氧化酶提取绑定在膜上, 当CH4气体经过这些镶嵌在膜上的颗粒态酶团簇时被转化为液态的CH3OH[170]。这些脱离出来的酶带来了无与伦比的富集效率, 但是这些酶的稳定性不佳, 使得该技术目前还不能投入到实际应用。这也反映出湖泊CH4资源化的主要矛盾, 即目前技术还不能很好地克服湖泊环境中低密度CH4与能源供应上对高密度的需求。但总体而言, CH4气—液分离技术的不断进步为拓宽湖泊碳汇价值打下了基础, 使湖泊在未来不仅能成为稳定的“碳库”, 还可能成为清洁能源的重要来源。
4 总结与展望碳中和需要在减排的同时充分发挥山、水、林、田、湖、草各生态功能区碳汇潜力, 扩大碳汇。目前对于湖泊生态系统的碳汇贡献仍然在讨论, 湖泊常被认定为碳源, 但众多研究结果显示湖泊生态系统以其独特的生境条件、发达的生产力可以实现对碳的汇入和累积。湖泊生态系统的光合-呼吸作用直接调整水体pCO2的同时增加水体碱度间接地加强CO2的汇入。沉积是湖泊碳累积的主要方式, 光合作用会带来的沉积物内源有机碳累积的加速, 结合呼吸分解的相对滞后实现总体的碳累积。生态系统生产力决定了固碳能力, 是湖泊发挥碳汇效益的重要“碳库”。由水生植物固定下的CO2总量不如浮游植物, 但在过程中发挥了“压舱石”般的稳定作用。
土地开发利用、全球气候变暖等问题正在破坏健康的湖泊生态系统, 导致湖泊生物多样性下降, 复杂的食物链体系被破坏, 给湖泊碳汇带来了负面影响。要发挥湖泊碳汇潜力, 利用好水生植物对碳的稳固吸收是当下总结出的可靠经验。在未来提高湖泊生物多样性, 增强碳在食物链中的系留可能才是真正发挥湖泊碳汇潜力的主要途径。富营养化湖泊应该作为研究的重点。富营养化湖泊虽然通过浮游藻类的光合作用实现短期水体CO2欠饱和状态, 但浮游藻类过度繁殖会破坏健康的湖泊生态系统, 被固定下的碳会很快再次释放回大气。近年来我国水环境质量已大幅改善, 但中/富营养化已经是当下许多湖泊的常态, 如何利用这些中/富营养化湖泊让其发挥碳汇效益成为一个现实问题。浮游植物既是湖泊实现碳汇的主要途径又可能破坏湖泊生态系统, 让湖泊丧失碳汇功能。所以关键就在于有效控制浮游植物的数量, 维持群落的多样性, 避免大规模同时衰亡、分解。为此未来需要开发更有效的浮游植物调控手段, 改善湖泊景观面貌的同时发挥中/富营化湖泊旺盛的初级生产力, 使其成为具有稳定碳汇价值的地理单元。
在一些CO2过饱和湖泊同时拥有着较高的沉积物OCAR, 单独依据pCO2来判断湖泊的碳汇/源可能会忽视湖泊生态系统的碳汇贡献。全面的碳平衡结算是评价湖泊碳源/汇的最可靠最理想方法, 但成本高耗时长。调查沉积物中碳的累积速率、来源以及分解程度, 以及与陆地单元碳累积速率的对比可能是一种更快捷的判定方法。未来对湖泊沉积物有机质分解代谢的研究会愈发得到关注, 湖泊碳汇价值还会因此得到扩展和认可。湖泊、水库沉积物微生物分解有机质产生的CH4能占全球每年产量的20%左右, 有学者就提出碳中和的关键可能就深藏于湖泊之下。将湖泊沉积物产CH4资源化被提出并进行了初步尝试, 可大规模应用的水体CH4分离技术也取得了进步[171]。为湖泊资源化, 拓宽湖泊碳汇价值打下了坚实基础。
| [1] |
Raymond P A, Hartmann J, Lauerwald R, Sobek S, McDonald C, Hoover M, Butman D, Striegl R, Mayorga E, Humborg C, Kortelainen P, Dürr H, Meybeck M, Ciais P, Guth P. Global carbon dioxide emissions from inland waters. Nature, 2013, 503(7476): 355-359. DOI:10.1038/nature12760 |
| [2] |
Lind O. Textbook of limnology. Limnology and Oceanography Bulletin, 2016, 25: 137-138. DOI:10.1002/lob.10142 |
| [3] |
Holgerson M A, Raymond P A. Large contribution to inland water CO2 and CH4 emissions from very small ponds. Nature Geoscience, 2016, 9(3): 222-226. DOI:10.1038/ngeo2654 |
| [4] |
Tranvik L J, Downing J A, Cotner J B, Loiselle S A, Striegl R G, Ballatore T J, Dillon P, Finlay K, Fortino K, Knoll L B, Kortelainen P L, Kutser T, Larsen S, Laurion I, Leech D M, McCallister S L, McKnight D M, Melack J M, Overholt E, Porter J A, Prairie Y, Renwick W H, Roland F, Sherman B S, Schindler D W, Sobek S, Tremblay A, Vanni M J, Verschoor A M, von Wachenfeldt E, Weyhenmeyer G A. Lakes and reservoirs as regulators of carbon cycling and climate. Limnology & Oceanography, 2009, 54(6): 2298-2314. |
| [5] |
Finlay K, Leavitt P R, Wissel B, Prairie Y T. Regulation of spatial and temporal variability of carbon flux in six hard-water lakes of the northern Great Plains. Limnol Oceanogr. Limnology and Oceanography, 2009, 54(6_part_2): 2553-2564. |
| [6] |
Valencia S, Marin J M, Rcstrepo G, Frimmel F H. Evaluations of the TiO2/simulated solar UV degradations of XAD fractions of natural organic matter from a bog lake using size-exclusion chromatography. Water Research, 2013, 47(14): 5130-5138. DOI:10.1016/j.watres.2013.05.053 |
| [7] |
Brinkmann T, Abbt-Braun G, Frimmel F H. Alkaline Degradation of Dissolved Organic Matter. Acta hydrochimica et hydrobiologica, 2003, 31(3): 213-224. DOI:10.1002/aheh.200300491 |
| [8] |
Sobek S, Tranvik L J, Prairie Y T. Patterns and Regulation of Dissolved Organic Carbon: An Analysis of 7, 500 Widely Distributed Lakes. Limnology & Oceanography, 2007, 52(3): 1208-1219. |
| [9] |
秦伯强, 高光, 朱广伟, 张运林, 宋玉芝, 汤祥明, 许海, 邓建明. 湖泊富营养化及其生态系统响应. 科学通报, 2013, 58(10): 855-864. |
| [10] |
Balmer M B, Downing J A. Carbon dioxide concentrations in eutrophic lakes: undersaturation implies atmospheric uptake. Inland Waters, 2011, 1(2): 125-132. DOI:10.5268/IW-1.2.366 |
| [11] |
Zagarese H E, de los Ángeles González Sagrario M, Wolf-Gladrow D, Nõges P, Nõges T, Kangur K, Matsuzaki S I S, Kohzu A, Vanni M J, Özkundakci D, Echaniz S A, Vignatti A, Grosman F, Sanzano P, Dam B V, Knoll L B. Patterns of CO2 concentration and inorganic carbon limitation of phytoplankton biomass in agriculturally eutrophic lakes. Water Research, 2021, 190: 116715. DOI:10.1016/j.watres.2020.116715 |
| [12] |
Lerman A, Stumm W. CO2 storage and alkalinity trends in lakes. Water Research, 1989, 23(2): 139-146. DOI:10.1016/0043-1354(89)90037-7 |
| [13] |
Xiao Q T, Xu X F, Duan H T, Qi T C, Qin B Q, Lee X H, Hu Z H, Wang W, Xiao W, Zhang M. Eutrophic Lake Taihu as a significant CO2 source during 2000-2015. Water Research, 2020, 170: 115331. DOI:10.1016/j.watres.2019.115331 |
| [14] |
王伯炜, 牟长城, 王彪. 长白山原始针叶林沼泽湿地生态系统碳储量. 生态学报, 2019(9): 3344-3354. |
| [15] |
李岩, 干珠扎布, 胡国铮, 万运帆, 李玉娥, 旦久罗布, 白玛玉珍, 高清竹. 增温对青藏高原高寒草原生态系统碳交换的影响. 生态学报, 2019(6): 2004-2012. |
| [16] |
池源, 石洪华, 孙景宽, 李捷, 杨帆, 付战勇. 近30年来黄河三角洲植被净初级生产力时空特征及主要影响因素. 生态学报, 2018, 38(08): 2683-2697. |
| [17] |
张富贵, 张舜尧, 唐瑞玲, 王惠艳, 杨志斌, 周亚龙, 孙忠军. 青藏高原湿地冻土区活动层甲烷排放特征. 物探与化探, 2017, 41(6): 1027-1036. |
| [18] |
陈吉龙, 何蕾, 温兆飞, 寥华军, 王炳亮, 崔林林, 李国胜. 辽河三角洲河口芦苇沼泽湿地植被固碳潜力. 生态学报, 2017, 37(16): 5402-5410. |
| [19] |
Abelmann A, Gersonde R, Knorr G, Zhang X, Chapligin B, Maier E, Esper O, Friedrichsen H, Lohmann G, Meyer H. The seasonal sea-ice zone in the glacial Southern Ocean as a carbon sink. Nature Communications, 2015, 6: 8136. DOI:10.1038/ncomms9136 |
| [20] |
潘竟虎, 文岩. 中国西北干旱区植被碳汇估算及其时空格局. 生态学报, 2015, 35(23): 7718-7728. |
| [21] |
Fang JY, Chen AP, Peng CH, Zhao SQ. Changes in forest biomass carbon storage in China between 1949 and 1998. SCIENCE, 2001, 292(5525): 2320-2322. DOI:10.1126/science.1058629 |
| [22] |
Sarmiento J L, Sundquist E T. Revised budget for the oceanic uptake of anthropogenic carbon dioxide. Nature, 1992, 356: 589-593. DOI:10.1038/356589a0 |
| [23] |
杨胜苏, 刘卫柏. 基于恢复生态学的洞庭湖区"山水林田湖草"生态修复研究. 生态学报, 2021, 41(16): 6430-6439. |
| [24] |
杨庆媛, 毕国华. 平行岭谷生态区生态保护修复的思路, 模式及配套措施研究——基于重庆市"两江四山"山水林田湖草生态保护修复工程试点. 生态学报, 2019, 39(23): 8939-8947. |
| [25] |
罗明, 周妍, 鞠正山, 魏洪斌, 张世文. 粤北南岭典型矿山生态修复工程技术模式与效益预评估——基于广东省山水林田湖草生态保护修复试点框架. 生态学报, 2019, 39(23): 8911-8919. |
| [26] |
刘世梁, 董玉红, 孙永秀, 史芳宁. 基于生态系统服务提升的山水林田湖草优先区分析——以贵州省为例. 生态学报, 2019, 39(23): 8957-8965. |
| [27] |
李涛, 唐涛, 邓红兵, 吴钢, 蔡庆华. 湖北省三峡地区山水林田湖草系统原理及生态保护修复研究. 生态学报, 2019, 39(23): 8896-8902. |
| [28] |
宋伟, 韩赜, 刘琳. 山水林田湖草生态问题系统诊断与保护修复综合分区研究——以陕西省为例. 生态学报, 2019, 39(23): 8975-8989. |
| [29] |
宇振荣, 杨新民, 陈雅杰. 河南省南太行地区山水林田湖草生态保护与修复. 生态学报, 2019, 39(23): 8886-8895. |
| [30] |
刘时栋, 刘琳, 张建军, 王柯, 郭义强. 基于生态系统服务能力提升的干旱区生态保护与修复研究——以额尔齐斯河流域生态保护与修复试点工程区为例. 生态学报, 2019, 39(23): 8998-9007. |
| [31] |
李潇, 吴克宁, 刘亚男, 冯喆, 谢家麟. 基于生态系统服务的山水林田湖草生态保护修复研究——以南太行地区鹤山区为例. 生态学报, 2019, 39(23): 8806-8816. |
| [32] |
田野, 冯启源, 唐明方, 郑拴宁, 柳彩霞, 吴迪, 王丽娜. 基于生态系统评价的山水林田湖草生态保护与修复体系构建研究——以乌梁素海流域为例. 生态学报, 2019, 39(23): 8826-8836. |
| [33] |
孔令桥, 郑华, 欧阳志云. 基于生态系统服务视角的山水林田湖草生态保护与修复——以洞庭湖流域为例. 生态学报, 2019, 39(23): 8903-8910. |
| [34] |
Visser P M, Verspagen J M H, Sandrini G, Stal L J, Matthijs H C P, Davis T W, Paerl H W, Huisman J. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae, 2016, 54(apr.): 145-159. |
| [35] |
Pacheco F, Landim P, Szocs T. Anthropogenic impacts on mineral weathering: A statistical perspective. Applied Geochemistry, 2013, 36(Complete): 34-48. |
| [36] |
Michelle B, John D. Carbon dioxide concentrations in eutrophic lakes: undersaturation implies atmospheric uptake. Inland Waters, 2011, 1(2): 125-132. DOI:10.5268/IW-1.2.366 |
| [37] |
Hesslein R H, Rudd J W M, Kelly C, Ramlal P, Hallard K. Carbon Dioxide Pressure in Surface Waters of Canadian Lakes. 1991.
|
| [38] |
Kratz T K, Schindler J, Hope D, Riera J L, Bowser C J. Average annual carbon dioxide concentrations in eight neighboring lakes in northern Wisconsin, USA. SIL Proceedings, 1922-2010, 1997, 26(2): 335-338.
|
| [39] |
Hein M. Inorganic carbon limitation of photosynthesis in lake phytoplankton. Freshwater Biology, 2010, 37(3): 545-552. |
| [40] |
Williamson T J, Vanni M J, Renwick W H. Spatial and Temporal Variability of Nutrient Dynamics and Ecosystem Metabolism in a Hyper-eutrophic Reservoir Differ Between a Wet and Dry Year. Ecosystems, 2021, 24(1): 68-88. DOI:10.1007/s10021-020-00505-8 |
| [41] |
Knoll L B, Vanni M J, Renwick W H, Dittman E K, Gephart J A. Temperate reservoirs are large carbon sinks and small CO2 sources: Results from high-resolution carbon budgets. Global Biogeochemical Cycles, 2013, 27(1): 52-64. DOI:10.1002/gbc.20020 |
| [42] |
Matsuzaki S I S, Suzuki K, Kadoya T, Nakagawa M, Takamura N. Bottom-up linkages between primary production, zooplankton, and fish in a shallow, hypereutrophic lake. Ecology, 2018, 99(9): 2025-2036. DOI:10.1002/ecy.2414 |
| [43] |
Tomioka N, Imai A, Komatsu K. Effect of light availability on Microcystis aeruginosa blooms in shallow hypereutrophic Lake Kasumigaura. Journal of Plankton Research, 2011, 33(8): 1263-1273. DOI:10.1093/plankt/fbr020 |
| [44] |
Verspagen J M H, van de Waal D B, Finke J F, Visser P M, van Donk E, Huisman J. Rising CO2 levels will intensify phytoplankton blooms in eutrophic and hypertrophic lakes. PLoS One, 2014, 9(8): e104325. DOI:10.1371/journal.pone.0104325 |
| [45] |
Tonetta D, Fontes M L S, Petrucio M M. Linking summer conditions to CO2 undersaturation and CO2 influx in a subtropical coastal lake. Limnology, 2015, 16(3): 193-201. DOI:10.1007/s10201-015-0460-9 |
| [46] |
Downing J A. Global limnology: up-scaling aquatic services and processes to planet Earth. SIL Proceedings, 1922-2010, 2009, 30(8): 1149-1166.
|
| [47] |
Abril G, Bouillon S, Darchambeau F, Teodoru C R, Marwick T R, Tamooh F, Ochieng Omengo F, Geeraert N, Deirmendjian L, Polsenaere P, Borges A V. Technical Note: Large overestimation of pCO2 calculated from pH and alkalinity in acidic, organic-rich freshwaters. Biogeosciences, 2015, 12(1): 67-78. DOI:10.5194/bg-12-67-2015 |
| [48] |
Sutcliffe D W. Acid precipitation and its effects on aquatic systems in the English Lake District. 1983.
|
| [49] |
Kilham P. Acid precipitation: Its role in the alkalization of a lake in Michiganl. Limnology and Oceanography, 1982, 27(5): 856-867. DOI:10.4319/lo.1982.27.5.0856 |
| [50] |
Kelly C A, Rudd J W M, Cook R B, Schindler D W. The potential importance of bacterial processes in regulating rate of lake acidification1, 2. Limnology and Oceanography, 1982, 27(5): 868-882. DOI:10.4319/lo.1982.27.5.0868 |
| [51] |
Niessen F, Sturm M. Die sedimente des baldeggersees (schweiz) \u2015 ablagerungsraum und eutrophierungsentwicklung während der letzten 100 jahre. Archiv Fur Hydrobiologie, 1987, 108: 365-383. DOI:10.1127/archiv-hydrobiol/108/1987/365 |
| [52] |
Reeder B C. Primary productivity limitations in relatively low alkalinity, high phosphorus, oligotrophic Kentucky reservoirs. Ecological Engineering, 2017, 108: 477-481. DOI:10.1016/j.ecoleng.2017.06.009 |
| [53] |
Narver, David W. Primary Productivity in the Babine Lake System, British Columbia. Journal of the Fisheries Board of Canada, 2011, 24(10): 2045-2052. |
| [54] |
Lerman A, Stumm W. CO2 storage and alkalinity trends in lakes. Water Research, 1989, 23(2): 139-146. DOI:10.1016/0043-1354(89)90037-7 |
| [55] |
The encyclopedia of sedimentology. Geological Magazine, 1979, 116(4): 330.
|
| [56] |
曹建华, 杨慧, 康志强. 区域碳酸盐岩溶蚀作用碳汇通量估算初探: 以珠江流域为例. 科学通报, 2011, 56(26): 2181-2187. |
| [57] |
唐伟. 外源水对碳酸盐岩溶蚀速率与碳汇效应影响研究——以毛村地下河流域为例[D]. 重庆: 西南大学, 2011.
|
| [58] |
Liu Z H. Role of carbonic anhydrase as an activator in carbonate rock dissolution and its implication for atmospheric CO2 sink. Acta Geologica Sinica-English Edition, 2001, 75(3): 275-278. |
| [59] |
谢腾祥. 微藻对碳酸盐矿物的生物溶蚀和沉淀作用及其碳汇效应[D]. 中国科学院大学, 2014.
|
| [60] |
Marotta H, Pinho L, Gudasz C, Bastviken D, Tranvik L J, Enrich-Prast A. Greenhouse gas production in low-latitude lake sediments responds strongly to warming. Nature Climate Change, 2014, 4(6): 467-470. DOI:10.1038/nclimate2222 |
| [61] |
Houghton R A. Balancing the global carbon budget. Annual Review of Earth and Planetary Sciences, 2007, 35: 313-347. DOI:10.1146/annurev.earth.35.031306.140057 |
| [62] |
Cole J J, Prairie Y T, Caraco N F, McDowell W H, Tranvik L J, Striegl R G, Duarte C M, Kortelainen P, Downing J A, Middelburg J J, Melack J. Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems, 2007, 10(1): 172-185. DOI:10.1007/s10021-006-9013-8 |
| [63] |
Tranvik L J, Downing J A, Cotner J B, Loiselle S A, Striegl R G, Ballatore T J, Dillon P, Finlay K, Fortino K, Knoll L B, Kortelainen P L, Kutser T, Larsen S, Laurion I, Leech D M, McCallister S L, McKnight D M, Melack J M, Overholt E, Porter J A, Prairie Y, Renwick W H, Roland F, Sherman B S, Schindler D W, Sobek S, Tremblay A, Vanni M J, Verschoor A M, von Wachenfeldt E, Weyhenmeyer G A. Lakes and reservoirs as regulators of carbon cycling and climate. Limnology and Oceanography, 2009, 54(6part2): 2298-2314. DOI:10.4319/lo.2009.54.6_part_2.2298 |
| [64] |
Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165-173. |
| [65] |
Dean W E, Gorham E. Magnitude and significance of carbon burial in lakes, reservoirs, and peatlands. Geology, 1998, 26(6): 535. |
| [66] |
Anderson N J, Engstrom D R, Leavitt P R, Flood S M, Heathcote A J. Changes in coupled carbon nitrogen dynamics in a tundra ecosystem predate post-1950 regional warming. Communications Earth & Environment, 2020, 1: 38. |
| [67] |
Wang S R, Zhuang Q L, Lähteenoja O, Draper F C, Cadillo-Quiroz H. Potential shift from a carbon sink to a source in Amazonian peatlands under a changing climate. PNAS, 2018, 115(49): 12407-12412. |
| [68] |
Dietz R D, Engstrom D R, Anderson N J. Patterns and drivers of change in organic carbon burial across a diverse landscape: Insights from 116 Minnesota lakes. Global Biogeochemical Cycles, 2015, 29(5): 708-727. |
| [69] |
Mulholland P J, Elwood J W. The role of lake and reservoir sediments as sinks in the perturbed global carbon cycle. Tellus, 1982, 34(5): 490-499. |
| [70] |
Ostrovsky I, Yacobi Y Z. Sedimentation flux in a large subtropical lake: Spatiotemporal variations and relation to primary productivity. Limnology and Oceanography, 2010, 55(5): 1918-1931. |
| [71] |
Lin Q, Liu E F, Zhang E L, Nath B, Bindler R, Liu J, Shen J. Organic carbon burial in a large, deep alpine lake (southwest China) in response to changes in climate, land use and nutrient supply over the past -100 years. CATENA, 2021, 202: 105240. |
| [72] |
Huang C C, Yao L, Zhang Y L, Huang T, Zhang M L, Zhu A X, Yang H. Spatial and temporal variation in autochthonous and allochthonous contributors to increased organic carbon and nitrogen burial in a plateau lake. Science of the Total Environment, 2017, 603/604: 390-400. |
| [73] |
Dong X H, Anderson N J, Yang X D, chen X, Shen J. Carbon burial by shallow lakes on the Yangtze floodplain and its relevance to regional carbon sequestration. Global Change Biology, 2012, 18(7): 2205-2217. |
| [74] |
Heathcote A J, Downing J A. Impacts of Landscape Change on Carbon Burial in Freshwater Lakes. 2010.
|
| [75] |
刘培. 腾冲青海湖近1700a来沉积物有机碳、氮同位素特征及古环境重建[D]. 昆明: 云南师范大学, 2018.
|
| [76] |
Lammers J M, Schubert C J, Middelburg J J, Reichart G J. Microbial carbon processing in oligotrophic Lake Lucerne (Switzerland): results of in situ 13C-labelling studies. Biogeochemistry, 2017, 136(2): 131-149. |
| [77] |
Forsius M, Kujala H, Minunno F, Holmberg M, Leikola N, Mikkonen N, Autio I, Paunu V V, Tanhuanpää T, Hurskainen P, Mäyraä J, Kivinen S, Keski-Saari S, Kosenius A K, Kuusela S, Virkkala R, Viinikka A, Vihervaara P, Heikkinen R K. Developing a spatially explicit modelling and evaluation framework for integrated carbon sequestration and biodiversity conservation: application in southern Finland. Science of the Total Environment, 2021, 775: 145847. |
| [78] |
Stewart T J, Sprules W G. Carbon-based balanced trophic structure and flows in the offshore Lake Ontario food web before (1987-1991) and after (2001-2005) invasion-induced ecosystem change. Ecological Modelling, 2011, 222(3): 692-708. |
| [79] |
Jensen T C, Zawiska I, Oksman M, Słowiński M, Woszczyk M, Luoto T P, Tylmann W, Nevalainen L, Obremska M, Schartau A K, Walseng B. Historical human impact on productivity and biodiversity in a subalpine oligotrophic lake in Scandinavia. Journal of Paleolimnology, 2020, 63(1): 1-20. |
| [80] |
Yang H, Xing Y P, Xie P, Ni L Y, Rong K W. Carbon source/sink function of a subtropical, eutrophic lake determined from an overall mass balance and a gas exchange and carbon burial balance. Environmental Pollution, 2008, 151(3): 559-568. |
| [81] |
Smith, S. V. Marine macrophytes as a global carbon sink. Science, 1981, 211(4484): 838-840. |
| [82] |
宋娜. 浅水湖泊沉积物中纤维素降解的过程特征及强化研究[D]. 中国科学院大学, 2013.
|
| [83] |
曹培培, 刘茂松, 唐金艳, 滕漱清, 徐驰. 几种水生植物腐解过程的比较研究. 生态学报, 2014, 34(14): 3848-3858. |
| [84] |
Guo X H, Xiao D R, Tian K, Yu H Z. Biomass production and litter decomposition of lakeshore plants in Napahai wetland, Northwestern Yunnan Plateau, China. Acta Ecologica Sinica, 2013, 33(5): 1425-1432. |
| [85] |
Einola E, Rantakari M, Kankaala P, Kortelainen P, Ojala A, Pajunen H, Mäkelä S, Arvola L. Carbon pools and fluxes in a chain of five boreal lakes: a dry and wet year comparison. Journal of Geophysical Research Atmospheres, 2011, 116(G3): G03009. |
| [86] |
Titus J E, Pagano A M. Carbon dioxide and submersed macrophytes in lakes: Linking functional ecology to community composition. Ecology, 2017, 98(12): 3096-3105. |
| [87] |
Tao F X. Air-water CO2 flux in an algae bloom year for Lake Hongfeng, Southwest China: implications for the carbon cycle of global inland waters. Acta Geochimica, 2017, 36(4): 658-666. |
| [88] |
倪兆奎, 李跃进, 王圣瑞, 金相灿, 储昭升. 太湖沉积物有机碳与氮的来源. 生态学报, 2011, 31(16): 4661-4670. |
| [89] |
Valle J, Gonsior M, Harir M, Enrich-Prast A, Schmitt-Kopplin P, Bastviken D, Conrad R, Hertkorn N. Extensive processing of sediment pore water dissolved organic matter during anoxic incubation as observed by high-field mass spectrometry (FTICR-MS). Water Research, 2018, 129: 252-263. |
| [90] |
Raymond J E, Fernandez I J, Ohno T, Simon K. Soil drainage class influences on soil carbon in a new England forested watershed. Soil Science Society of America Journal, 2013, 77(1): 307-317. |
| [91] |
Sobek A, Olli K, Gustafsson O. On the relative significance of bacteria for the distribution of polychlorinated biphenyls in Arctic Ocean surface waters. Environmental Science & Technology, 2006, 40(8): 2586-2593. |
| [92] |
Hessen D O, Nygaard K. Bacterial transfer of methane and detritus: implications for the pelagic carbon budget and gaseous release. Archiv fur Hydrobiologie, 1992(37): 139-148. |
| [93] |
Pace, Carpenter S R, Cole J J. Terrestrial subsidies of aquatic food webs: Results of 13C additions to lakes contrasting in dissolved organic matter and nutrients. 2005.
|
| [94] |
Kritzberg E S, Cole J J, Pace M L, Granéli W, Bade D L. Autochthonous versus allochthonous carbon sources of bacteria: results from whole-lake 13C addition experiments. Limnology and Oceanography, 2004, 49(2): 588-596. |
| [95] |
Wilkinson G M, Buelo C D, Cole J J, Pace M L. Exogenously produced CO2 doubles the CO2 efflux from three north temperate lakes. Geophysical Research Letters, 2016, 43(5): 1996-2003. |
| [96] |
Kortelainen P, Rantakari M, Huttunen J T, Mattsson T, Alm J, Juutinen S, Larmola T, Silvola J, Martikainen P J. Sediment respiration and lake trophic state are important predictors of large CO2 evasion from small boreal lakes. Global Change Biology, 2006, 12(8): 1554-1567. |
| [97] |
Wang Y H, Huang Y, Tian J M, Li C H, Yu K K, Zhang M L, Lang X L, Sun T. A sediment record of terrestrial organic matter inputs to Dongting Lake and its environmental significance from 1855 to 2019. Ecological Indicators, 2021, 130: 108090. |
| [98] |
Radbourne A D, Ryves D B, Anderson N J, Scott D R. The historical dependency of organic carbon burial efficiency. Limnology and Oceanography, 2017, 62(4): 1480-1497. |
| [99] |
Marotta H, Pinho L, Gudasz C, Bastviken D, Tranvik L J, Enrich-Prast A. Greenhouse gas production in low-latitude lake sediments responds strongly to warming. Nature Climate Change, 2014, 4(6): 467-470. |
| [100] |
Gudasz C, Bastviken D, Steger K, Premke K, Sobek S, Tranvik L J. Temperature-controlled organic carbon mineralization in lake sediments. Nature, 2010, 466(7305): 478-481. |
| [101] |
Sobek S, Durisch-Kaiser E, Zurbrügg R, Wongfun N, Wessels M, Pasche N, Wehrli B. Organic carbon burial efficiency in lake sediments controlled by oxygen exposure time and sediment source. Limnology and Oceanography, 2009, 54(6): 2243-2254. DOI:10.4319/lo.2009.54.6.2243 |
| [102] |
Wang H X, Jiao R Y, Wang F, Zhang L, Yan W J. Dissolved organic carbon content and characteristics in relation to carbon dioxide partial pressure across Poyang Lake wetlands and adjacent aquatic systems in the Changjiang Basin. Environmental Pollution, 2016, 219: 714-723. DOI:10.1016/j.envpol.2016.07.012 |
| [103] |
Perga M E, Maberly S C, Jenny J P, Alric B, Pignol C, Naffrechoux E. A century of human-driven changes in the carbon dioxide concentration of lakes. Global Biogeochemical Cycles, 2016, 30(2): 93-104. DOI:10.1002/2015GB005286 |
| [104] |
Yvon-Durocher G, Caffrey J M, Cescatti A, Dossena M, Giorgio P D, Gasol J M, Montoya J M, Pumpanen J, Staehr P A, Trimmer M, Woodward G, Allen A P. Reconciling the temperature dependence of respiration across timescales and ecosystem types. Nature, 2012, 487(7408): 472-476. DOI:10.1038/nature11205 |
| [105] |
Maberly S C, Barker P A, Stott A W, de Ville M M. Catchment productivity controls CO2 emissions from lakes. Nature Climate Change, 2013, 3(4): 391-394. DOI:10.1038/nclimate1748 |
| [106] |
Reed D E, Dugan H A, Flannery A L, Desai A R. Carbon sink and source dynamics of a eutrophic deep lake using multiple flux observations over multiple years. Limnology and Oceanography Letters, 2018, 3(3): 285-292. DOI:10.1002/lol2.10075 |
| [107] |
Dugan H A, Woolway R I, Santoso A B, Corman J R, Jaimes A, Nodine E R, Patil V P, Zwart J A, Brentrup J A, Hetherington A L, Oliver S K, Read J S, Winters K M, Hanson P C, Read E K, Winslow L A, Weathers K C. Consequences of gas flux model choice on the interpretation of metabolic balance across 15 lakes. Inland Waters, 2016, 6(4): 581-592. DOI:10.1080/IW-6.4.836 |
| [108] |
Solomon C T, Bruesewitz D A, Richardson D C, Rose K C, van de Bogert M C, Hanson P C, Kratz T K, Larget B, Adrian R, Babin B L, Chiu C Y, Hamilton D P, Gaiser E E, Hendricks S, Istvànovics V, Laas A, O' Donnell D M, Pace M L, Ryder E, Staehr P A, Torgersen T, Vanni M J, Weathers K C, Zhu G W. Ecosystem respiration: drivers of daily variability and background respiration in lakes around the globe. Limnology and Oceanography, 2013, 58(3): 849-866. DOI:10.4319/lo.2013.58.3.0849 |
| [109] |
Shao C L, Chen J Q, Stepien C A, Chu H S, Ouyang Z T, Bridgeman T B, Czajkowski K P, Becker R H, John R. Diurnal to annual changes in latent, sensible heat, and CO2 fluxes over a Laurentian Great Lake: a case study in Western Lake Erie. Journal of Geophysical Research: Biogeosciences, 2015, 120(8): 1587-1604. DOI:10.1002/2015JG003025 |
| [110] |
Grace J B, Anderson T M, Seabloom E W, Borer E T, Adler P B, Harpole W S, Hautier Y, Hillebrand H, Lind E M, Pärtel M, Bakker J D, Buckley Y M, Crawley M J, Damschen E I, Davies K F, Fay P A, Firn J, Gruner D S, Hector A, Knops J M H, MacDougall A S, Melbourne B A, Morgan J W, Orrock J L, Prober S M, Smith M D. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature, 2016, 529(7586): 390-393. DOI:10.1038/nature16524 |
| [111] |
Waide R B, Willig M R, Steiner C F, Mittelbach G, Gough L, Dodson S I, Juday G P, Parmenter R. The relationship between productivity and species richness. Annual Review of Ecology and Systematics, 1999, 30: 257-300. DOI:10.1146/annurev.ecolsys.30.1.257 |
| [112] |
Huston M A. A General Hypothesis of Species Diversity. The American Naturalist, 1979, 113(1): 81-101. DOI:10.1086/283366 |
| [113] |
Hodáňová D. Plant strategies and vegetation processes. Biologia Plantarum, 1981, 23(4): 254. DOI:10.1007/BF02895358 |
| [114] |
Wu A P, Ye S Y, Yuan J R, Qi L Y, Cai Z W, Ye B B, Yuan J, Chu Z S, Xie Y H, Liu L, Zhong W, Wang Y H. The relationship between diversity and productivity from a three-dimensional space view in a natural mesotrophic lake. Ecological Indicators, 2021, 121: 107069. DOI:10.1016/j.ecolind.2020.107069 |
| [115] |
Cardinale B J, Matulich K L, Hooper D U, Byranes J E, Duffy E, Gamfeldt L, Balvanera P, O' Connor M I, Gonzalez A. The functional role of producer diversity in ecosystems. American Journal of Botany, 2011, 98(3): 572-592. DOI:10.3732/ajb.1000364 |
| [116] |
Barnett A, Beisner B E. Zooplankton biodiversity and lake trophic state: explanations invoking resource abundance and distribution. Ecology, 2007, 88(7): 1675-1686. DOI:10.1890/06-1056.1 |
| [117] |
Gustafsson C, Norkko A. Quantifying the importance of functional traits for primary production in aquatic plant communities. Journal of Ecology, 2019, 107(1): 154-166. DOI:10.1111/1365-2745.13011 |
| [118] |
Naeem S, Thompson L J, Lawler S P, Lawton J H, Woodfin R M. Declining biodiversity can alter the performance of ecosystems. Nature, 1994, 368(6473): 734-737. DOI:10.1038/368734a0 |
| [119] |
Dimitrakopoulos P G, Schmid B. Biodiversity effects increase linearly with biotope space. Ecology Letters, 2004, 7(7): 574-583. DOI:10.1111/j.1461-0248.2004.00607.x |
| [120] |
Evans M A, Fahnenstiel G, Scavia D. Incidental oligotrophication of North American great lakes. Environmental Science & Technology, 2011, 45(8): 3297-3303. |
| [121] |
Cardinale B J, Matulich K L, Hooper D U, Byranes J E, Duffy E, Gamfeldt L, Balvanera P, O' Connor M I, Gonzalez A. The functional role of producer diversity in ecosystems. American Journal of Botany, 2011, 98(3): 572-592. DOI:10.3732/ajb.1000364 |
| [122] |
Stachowicz J J, Graham M, Bracken M E S, Szoboszlai A I. Diversity enhances cover and stability of seaweed assemblages: the role of heterogeneity and time. Ecology, 2008, 89(11): 3008-3019. DOI:10.1890/07-1873.1 |
| [123] |
Ansari A A, Gill S S, Khan F A. Eutrophication: Threat to Aquatic EcosystemsEutrophication: Causes. Consequences and Control, 2011, 89(11): 3008-3019. |
| [124] |
王志强, 崔爱花, 缪建群, 王海, 黄国勤. 淡水湖泊生态系统退化驱动因子及修复技术研究进展. 生态学报, 2017, 37(18): 6253-6264. |
| [125] |
秦伯强. 湖泊生态恢复的基本原理与实现. 生态学报, 2007, 27(11): 4848-4858. DOI:10.3321/j.issn:1000-0933.2007.11.055 |
| [126] |
秦伯强, 高光, 胡维平, 吴庆龙, 胡春华, 刘正文, 谷孝鸿, 朱广伟, 陈非洲. 浅水湖泊生态系统恢复的理论与实践思考. 湖泊科学, 2005, 17(1): 9-16. DOI:10.3321/j.issn:1003-5427.2005.01.002 |
| [127] |
Yuan X M, Liu Q, Cui B S, Xu X F, Liang L Q, Sun T, Yan S R, Wang X, Li C H, Li S Z, Li M. Effect of water-level fluctuations on methane and carbon dioxide dynamics in a shallow lake of Northern China: implications for wetland restoration. Journal of Hydrology, 2021, 597: 126169. DOI:10.1016/j.jhydrol.2021.126169 |
| [128] |
Chao C X, Wang L G, Li Y, Yan Z W, Liu H M, Yu D, Liu C H. Response of sediment and water microbial communities to submerged vegetations restoration in a shallow eutrophic lake. Science of the Total Environment, 2021, 801: 149701. DOI:10.1016/j.scitotenv.2021.149701 |
| [129] |
Fujibayashi M, Takakai F, Masuda S, Okano K, Miyata N. Effects of restoration of emergent macrophytes on the benthic environment of the littoral zone of a eutrophic lake. Ecological Engineering, 2020, 155: 105960. DOI:10.1016/j.ecoleng.2020.105960 |
| [130] |
姚程, 胡小贞, 姜霞, 陈俊伊, 王坤. 太湖贡湖湾人工湖滨带水生植物恢复及其富营养化控制. 湖泊科学, 2021, 33(6): 1626-1638. |
| [131] |
姚程, 胡小贞, 卢少勇, 耿荣妹. 几种水陆交错带植物对底质氮磷释放的抑制作用. 环境科学, 2017, 38(2): 589-599. DOI:10.13227/j.hjkx.201604237 |
| [132] |
Gao H L, Qian X, Wu H F, Li H M, Pan H, Han C M. Combined effects of submerged macrophytes and aquatic animals on the restoration of a eutrophic water body-A case study of Gonghu Bay, Lake Taihu. Ecological Engineering, 2017, 102: 15-23. DOI:10.1016/j.ecoleng.2017.01.013 |
| [133] |
姚程, 胡小贞, 耿荣妹, 薛彦君, 成小英. 几种水陆交错带植物对湖滨带底质的稳固作用. 湖泊科学, 2017, 29(1): 105-115. |
| [134] |
Bai G L, Zhang Y, Yan P, Yan W H, Kong L W, Wang L, Wang C, Liu Z S, Liu B Y, Ma J M, Zuo J C, Li J, Bao J, Xia S B, Zhou Q H, Xu D, He F, Wu Z B. Spatial and seasonal variation of water parameters, sediment properties, and submerged macrophytes after ecological restoration in a long-term (6 year) study in Hangzhou west lake in China: Submerged macrophyte distribution influenced by environmental variables. Water Research, 2020, 186: 116379. DOI:10.1016/j.watres.2020.116379 |
| [135] |
Ferretti A R, de Britez R M. Ecological restoration, carbon sequestration and biodiversity conservation: the experience of the Society for Wildlife Research and Environmental Education (SPVS) in the Atlantic Rain Forest of Southern Brazil. Journal for Nature Conservation, 2006, 14(3/4): 249-259. |
| [136] |
Kareksela S, Haapalehto T, Juutinen R, Matilainen R, Tahvanainen T, Kotiaho J S. Fighting carbon loss of degraded peatlands by jump-starting ecosystem functioning with ecological restoration. Science of the Total Environment, 2015, 537: 268-276. DOI:10.1016/j.scitotenv.2015.07.094 |
| [137] |
Zhang X J, Wang G Q, Tan Z X, Wang Y T, Li Q. Effects of ecological protection and restoration on phytoplankton diversity in impounded lakes along the eastern route of China' s South-to-North Water Diversion Project. Science of the Total Environment, 2021, 795: 148870. DOI:10.1016/j.scitotenv.2021.148870 |
| [138] |
Wang Y T, Wang W C, Zhou Z Z, Xia W, Zhang Y X. Effect of fast restoration of aquatic vegetation on phytoplankton community after removal of purse seine culture in Huayanghe Lakes. Science of the Total Environment, 2021, 768: 144024. DOI:10.1016/j.scitotenv.2020.144024 |
| [139] |
Chen F Z, Ye J L, Shu T T, Sun Y, Li J. Zooplankton response to the lake restoration in the drinking-water source in Meiliang Bay of subtropical eutrophic Lake Taihu, China. Limnologica, 2012, 42(3): 189-196. DOI:10.1016/j.limno.2011.11.001 |
| [140] |
Burlakova L E, Karatayev A Y, Pennuto C, Mayer C. Changes in Lake Erie benthos over the last 50 years: historical perspectives, current status, and main drivers. Journal of Great Lakes Research, 2014, 40(3): 560-573. DOI:10.1016/j.jglr.2014.02.008 |
| [141] |
Molina-Moctezuma A, Godby N, Kapuscinski K L, Roseman E F, Skubik K, Moerke A. Response of fish assemblages to restoration of rapids habitat in a Great Lakes connecting channel. Journal of Great Lakes Research, 2021, 47(4): 1182-1191. DOI:10.1016/j.jglr.2021.05.009 |
| [142] |
Zhang S Q, Zhang P Y, Pan B H, Zou Y A, Xie Y H, Zhu F, Chen X S, Li F, Deng Z M, Zhang H, Yang S. Wetland restoration in the East Dongting Lake effectively increased waterbird diversity by improving habitat quality. Global Ecology and Conservation, 2021, 27: e01535. DOI:10.1016/j.gecco.2021.e01535 |
| [143] |
Zhang X X, Yi Y J, Yang Z F. The long-term changes in food web structure and ecosystem functioning of a shallow lake: implications for the lake management. Journal of Environmental Management, 2022, 301: 113804. DOI:10.1016/j.jenvman.2021.113804 |
| [144] |
汪旋, 李致春, 李翔, 许晓光, 张思远, 王慧敏, 厉荣强, 王国祥. 蓝藻水华衰亡对沉积物碳库有机碳组分的影响. 湖泊科学, 2020, 32(2): 346-356. |
| [145] |
于志同, 王秀君, 赵成义, 兰海燕. 基于多指标分析的博斯腾湖表层沉积物有机碳来源. 湖泊科学, 2015, 27(5): 983-990. |
| [146] |
易文利, 王圣瑞, 杨苏文, 赵海超, 金相灿, 王国栋. 长江中下游浅水湖泊沉积物腐殖质组分赋存特征. 湖泊科学, 2011(1): 21-28. |
| [147] |
居琪, 章奇, 曹驰程, 白雷雷, 张晖. 湖泊沉积物中微生物群落对天然有机质输入的响应. 环境科学, 2021, 42(7): 3263-3271. |
| [148] |
闫兴成, 王明玥, 许晓光, 王国祥, 孙浩, 杨云皓, 石傲. 富营养化湖泊沉积物有机质矿化过程中碳, 氮, 磷的迁移特征. 湖泊科学, 2018, 30(2): 306-313. |
| [149] |
王鹏, 尚英男, 沈立成, 伍坤宇, 肖琼. 青藏高原淡水湖泊水化学组成特征及其演化. 环境科学, 2013, 34(3): 874-881. |
| [150] |
席北斗, 张亚丽, 许秋瑾. 矿化度作为蒙新高原湖泊营养物基准影响指标的可行性. 环境科学, 2012, 33(10): 3308-3313. |
| [151] |
汪旋, 李致春, 李翔, 许晓光, 张思远, 王慧敏, 厉荣强, 王国祥. 蓝藻水华衰亡对沉积物碳库有机碳组分的影响. 湖泊科学, 2020, 32(2): 346-356. |
| [152] |
Zhang Y D, Su Y L, Liu Z W, Yu J L, Jin M. Lipid biomarker evidence for determining the origin and distribution of organic matter in surface sediments of Lake Taihu, Eastern China. Ecological Indicators, 2017, 77: 397-408. DOI:10.1016/j.ecolind.2017.02.031 |
| [153] |
Gudasz C, Sobek S, Bastviken D, Koehler B, Tranvik L J. Temperature sensitivity of organic carbon mineralization in contrasting lake sediments. Journal of Geophysical Research: Biogeosciences, 2015, 120(7): 1215-1225. DOI:10.1002/2015JG002928 |
| [154] |
刘洋, 陈永娟, 王晓燕, 许康利. 人类活动对河流沉积物中反硝化厌氧甲烷氧化菌群落特征的影响. 环境科学, 2018, 39(8): 3677-3688. |
| [155] |
韩秀秀, 黄晓虎, 余丽燕, 杨浩, 黄昌春, 黄涛, 余艳红, 罗玉. 滇池沉积物有机质沉积特征与来源解析. 环境科学, 2017, 38(1): 78-86. |
| [156] |
邱祖凯, 胡小贞, 姚程, 张文慧, 许秋瑾, 黄天寅. 山美水库沉积物氮磷和有机质污染特征及评价. 环境科学, 2016(4): 1389-1396. |
| [157] |
李璐璐, 江韬, 闫金龙, 郭念, 魏世强, 王定勇, 高洁, 赵铮. 三峡库区典型消落带土壤及沉积物中溶解性有机质(DOM)的紫外-可见光谱特征. 环境科学, 2014, 35(3): 933-941. |
| [158] |
Yang B J, Ljung K, Nielsen A B, Fahlgren E, Hammarlund D. Impacts of long-term land use on terrestrial organic matter input to lakes based on lignin phenols in sediment records from a Swedish forest lake. Science of the Total Environment, 2021, 774: 145517. |
| [159] |
Xu L M, Li Y, Ye W T, Zhang X Z. Terrestrial organic carbon storage modes based on relationship between soil and lake carbon, China. Journal of Environmental Management, 2019, 250: 109483. |
| [160] |
Fletcher S E M, Schaefer H. Rising methane: a new climate challenge. Science, 2019, 364(6444): 932-933. |
| [161] |
Bastviken D, Tranvik L J, Downing J A, Crill P M, Enrich-Prast A. Freshwater methane emissions offset the continental carbon sink. Science, 2011, 331(6013): 50. |
| [162] |
Eyerer S, Schifflechner C, Hofbauer S, Bauer W, Wieland C, Spliethoff H. Combined heat and power from hydrothermal geothermal resources in Germany: an assessment of the potential. Renewable and Sustainable Energy Reviews, 2020, 120: 109661. |
| [163] |
Bastviken D, Cole J J, Pace M L, van de Bogert M C. Fates of methane from different lake habitats: connecting whole-lake budgets and CH4 emissions. Journal of Geophysical Research: Biogeosciences, 2008, 113(G2): G02024. |
| [164] |
Tofieldpasche N. Monitoring methane extraction in Lake Kivu. General Information, 2013.
|
| [165] |
Mason J A, Veenstra M, Long J R. Evaluating metal-organic frameworks for natural gas storage. Chem Sci, 2014, 5(1): 32-51. |
| [166] |
Cookney J, Mcleod A, Mathioudakis V, Ncube P, Soares A, Jefferson B, McAdam E J. Dissolved methane recovery from anaerobic effluents using hollow fibre membrane contactors. Journal of Membrane Science, 2016, 502: 141-150. |
| [167] |
Giménez J B, Martí N, Ferrer J, Seco A. Methane recovery efficiency in a submerged anaerobic membrane bioreactor (SAnMBR) treating sulphate-rich urban wastewater: evaluation of methane losses with the effluent. Bioresource Technology, 2012, 118: 67-72. |
| [168] |
Rezaei J, Bouteh E, Torabian A, Ghulami H. Evaluation of membrane bioreactor-hollow fiber (MBR-HF) pilot performance in the treatment of wastewater facing with different concentrations of amoxicillin (AMX) as shock loads. Journal of Environmental Chemical Engineering, 2020, 8(4): 103944. |
| [169] |
Sethunga G S M D P, Karahan H E, Wang R, Bae T H. Wetting- and fouling-resistant hollow fiber membranes for dissolved methane recovery from anaerobic wastewater treatment effluents. Journal of Membrane Science, 2021, 617: 118621. |
| [170] |
Blanchette C D, Knipe J M, Stolaroff J K, DeOtte J R, Oakdale J S, Maiti A, Lenhardt J M, Sirajuddin S, Rosenzweig A C, Baker S E. Printable enzyme-embedded materials for methane to methanol conversion. Nature Communications, 2016, 7: 11900. |
| [171] |
Bartosiewicz M, Rzepka P, Lehmann M F. Tapping freshwaters for methane and energy. Environmental Science & Technology, 2021, 55(8): 4183-4189. |