 2022, Vol. 42
2022, Vol. 42文章信息
- 王秋生, 温璐, 苏旭坤
- WANG Qiusheng, WEN Lu, SU Xukun
- 气候变化背景下青藏高原藏羚羊生境时空格局演变
- Spatial-temporal pattern changes of Tibetan antelope's habitats on the Qinghai-Tibetan Plateau under climate change scenarios
- 生态学报. 2022, 42(22): 8985-8993
- Acta Ecologica Sinica. 2022, 42(22): 8985-8993
- http://dx.doi.org/10.5846/stxb202205271503
-
文章历史
- 收稿日期: 2022-05-27





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;
3. 中国科学院大学, 北京 100049
2. State Key Laboratory Urban and Regional Ecology Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
根据《IPCC第六次评估报告》的评估结果显示, 在未来几十年里, 所有地区的气候变化都将加剧且仍得不到有效遏制, 而不可遏制的气候变化仍被认为是21世纪全球生物多样性面临的最主要威胁之一[1]。人为原因引起的气候变化正以前所未有的速度改变着地球上物种的分布格局, 最明显的表现就是物种分布范围向高纬度或高海拔移动[2—3]。被誉为“世界屋脊”“地球第三极”的青藏高原[4], 是珍稀野生动物的天然栖息地和高原物种基因库, 是中国乃至亚洲重要的生态安全屏障, 是中国生态文明建设的重点地区之一。在青藏高原的气候环境不断变化的背景下, 地表环境和生态系统变化对气候变化的响应与反馈过程将产生一系列影响[5—6]。地表环境和生态系统变化也直接影响着物种的分布格局。青藏高原的物种丰富度在气候变化的影响下, 从东到西逐渐下降;虽然东南山区拥有较高的物种丰富度, 却是生物多样性损失最严重的地区。21世纪很有可能看到在青藏高原北部有蹄类物种增加的现象, 青藏高原野生动物的分布区域减少, 物种丰富度格局也将发生显著变化[7]。只根据当前的分布情况建立自然保护区和国家公园来保护目标物种是远远不够的[8—9]。将自然气候因素和人为气候因素结合以获得更准确的分布预测结果, 此研究使用的气候数据采用CMIP6模式采用的共享社会经济路径(Shared Socioeconomic Pathways, SSPs)包括SSP126、SSP245、SSP370和SSP585四种共享社会经济路径情景, 能够反映辐射强迫和社会经济发展间的关联, 体现气候因素与社会经济因素间的综合作用, 更有利气候变化预估与影响的研究[10]。因此, 采用最新的气候数据, 针对不同的气候情景来评估物种分布, 将保护行动的规模和自然保护地的优化和管理与气候变化预测的结果相匹配便显得极为重要[11—12]。
藏羚羊(Pantholops hodgsonii)被誉为“高原精灵”, 其已有百万年的演化历史, 是青藏高原的特有种与旗舰种[13]。藏羚羊是《国家重点保护野生动物名录》中的国家一级保护野生动物, 《濒危野生动植物种国际贸易公约》中被列为濒危野生动物[14], 2016年被世界自然保护联盟(IUCN)濒危野生动物保护红色目录《Red List of Threatened Species》列为近危(NT, Near Threatened)物种(IUCN, 2016)[15]。随着保护政策实施和保护力度加强, 藏羚羊的种群数量逐渐恢复。粗略估计, 目前青藏高原藏羚羊数量大约为30万只[16], 但近年来, 青藏高原的气候变化、基础设施建设以及放牧等人为干扰日益加剧, 藏羚羊赖以生存的草场和栖息地被破坏, 并导致生境破碎化和丧失, 直接威胁着藏羚羊等有蹄类野生动物的生存和种群延续[17]。对生活在生态脆弱地区的藏羚羊来说, 活动范围或生活习惯都受到限制, 极小的气候事件都有可能对藏羚羊的栖息地产生不成比例的影响[18]。另外, 我国青藏高原建立的自然保护区大多存在保护与经济发展的冲突, 缺乏统筹规划, 藏羚羊等珍稀濒危野生动物存在保护空缺等问题[4]。最大熵模型(MaxEnt)相比其他物种分布模型, 结果更精准, 在时空外推性方面也表现更好, 被广泛应用于珍稀濒危物种的地理分布及气候变化背景下的生境时空格局演变的研究[19—23], 如Luo等利用物种分布模型评估未来气候变化对青藏高原32种有蹄类动物的潜在影响[7];晏婷婷利用物种分布模型评估未来气候变化对我国特有物种大熊猫的潜在影响[24]。
因此, 本研究中应用物种分布模型, 在当前和未来气候变化情景下模拟藏羚羊生境时空格局演变并确定影响其分布的最主要因素, 明确百年尺度藏羚羊保护空缺变化。研究藏羚羊生境时空变化, 对青藏高原生物多样性保护与维持具有重要意义, 对青藏高原自然保护地体系优化提供科学支撑并助力青藏高原生态安全屏障建设。
1 材料与方法 1.1 研究区概况青藏高原平均海拔4000米以上, 地势复杂, 气候独特,常与南极和北极作比, 形象地被称为“世界第三极”[25]。青藏高原地理位置为73°19′—104°47′E, 26°00′—39°47′N, 总面积约为2.5×107 km2。青藏高原特殊的地理环境和地表特征形成了极其复杂的气候条件, 拥有大量的藏羚羊适宜生境[13, 16]。本文中的研究区边界(图 1)依照中国科学院生态环境研究中心的中国生态功能区划并结合保护区及青藏高原边界。研究区内已建立1个国家公园、3个国家级自然保护区和1个省级自然保护区用于保护藏羚羊等有蹄类野生动物及其生境, 分别是: 三江源国家公园(面积1.5×105 km2)、新疆阿尔金山国家级自然保护区(面积4.68×104 km2)、可可西里国家级自然保护区(面积5.03×104 km2)、羌塘国家级自然保护区(面积3.02×105 km2)和新疆中昆仑自治区级保护区(面积3.2×104 km2)。

|
| 图 1 研究区位置示意图 Fig. 1 Location of study area |
藏羚羊在不同季节的大致生境范围主要是通过查阅文献和报告获取, 并于2012年12月、2013年6月和2021年8—9月, 开展野外样线调查, 野外调查获取阿尔金山国家级保护区和羌塘国家级保护区的藏羚羊出现点位信息。根据提前布设的样线, 沿样线以10—20 km/h车速, 通过肉眼观察并辅以蔡斯8倍双筒望远镜, 用手持GPS记录观测出现点坐标, 并记录距藏羚羊距离、雌雄亚幼、相对角度、地形和距水源地距离等指标, 通过分析计算得到藏羚羊实际出现点位信息。通过样线调查法共得到420个藏羚羊点位数据, 另外从相关文献中获取部分藏羚羊出现点位坐标并进行数字化, 通过生境信息和ArcGIS的地理配准对比校正, 获取了70个藏羚羊点位数据作为补充[26]。将所有样点通过ArcGIS缓冲区分析法筛选, 以0.5 km作为缓冲半径, 若1 km范围出现多点则仅保留1个。最终获得青藏高原藏羚羊有效点位386个。
1.2.2 气象数据和地形数据获取本研究涉及9个环境因子, 包括6个气候因子、3个地形因子(海拔、坡度和坡向)。利用R语言的dismo安装包, 对Wordcilm2.1(http://www.worldclim.org/)网站下载的当前气候数据(2000—2018年)进行处理。根据每月最高气温、最低气温、降水量三个数据项, 计算出2000—2018年的19个生物气候变量(Bio1—Bio19)。利用SPSS对19个生物气候因子进行相关性分析, 剔除相关性大于0.8的变量, 最终确定温度季节性变化(Bio4), 温度年变幅(Bio7), 最干季均温(Bio9), 最干月降雨(Bio14), 最湿季降雨(Bio16)和最干季降雨(Bio17)作为气候变量, 分辨率为30″(面积约0.83km2)[27—29]。海拔数据来源于国家地球系统科学数据中心, 基于GIS技术生成坡度(slope)及坡向(aspect), 分辨率统一为30″(面积约0.83km2)。
未来生物气候变量(Bio1—Bio19)数据采用国际耦合模式计划第六阶段(CMIP6)的MIROC6模式模拟结果(https://esgf.node.llnl.gov/projects/cmip6/), 模拟时段为2021—2100年。本文选取2021—2040年, 2041—2061年, 2061—2080年, 2081—2100年四个时段。气候压迫情景选择SSP126、SSP585两种极端的路径情景和SSP245中间路径情景。空间分辨率与当前气候条件下的气候因子数据保持一致。
1.2.3 潜在生境识别藏羚羊的潜在生境识别应用最大熵模型(MaxEnt)完成, 将已知的藏羚羊点位数据和利用ArcGIS 10.6软件转化工具生成的ASCII格式的气候图层导入MaxEnt软件进行运算[29]。运算过程中, 设置训练数据集为75%, 测试验证数据集为25%, 迭代次数为10000, 设置调控倍频为1, 重复训练数为10, 进行数据模拟。在MaxEnt软件中, 检验各生物气候变量对青藏高原藏羚羊地理分布的限制强度的生物气候变量贡献率是由Jackknife模块计算得到[30]。
进行气候变化的影响分析时, 通过最大化卡帕值(maximum Kappa value)的阈值选取方法, 将MaxEnt模型预测结果在ArcGIS 10.6软件中通过重分类转化成适宜生境(0)和适宜生境(1)的栅格数据[31—33]。通过ArcGIS栅格计算器统计基准气候和未来各种气候条件下藏羚羊潜在适宜生境栅格数量得出对应气候条件下的藏羚羊潜在适宜生境面积。对基准气候条件下藏羚羊潜在适宜生境与未来不同气候条件下的藏羚羊潜在适宜生境进行叠加分析, 公式为:
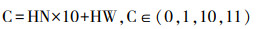
|
式中, HN为基准气候条件下的预测结果;HW为未来某一气候路径情景。X=10表示中高适宜生境→低适宜生境;X=1表示低适宜生境→中高适宜生境;X=0表示不变低适宜生境;X=11不变中高适宜生境。
MaxEnt模型预测的结果是0到1之间的ASCII数据, 而且适生值越高, 代表物种存在的概率就越高[34]。本研究中, 我们选择模型敏感度和特异性数值之和最大MSS (maximum sensitivity plus specificity)的阈值作为重分类的阈值, 阈值为0.195和0.380。对模型结果中的13个MSS和ESS值进行均值化处理, 结合其他学者的阈值判定方法将预测结果划分为三个等级[21, 22, 27]: 适生值< 0.2为低适生区;0.2 ≤适生值< 0.4为中适生区;0.4 ≤适生值< 1为高适生区。中适生区和高适生区所在的区域是藏羚羊的适宜生境的概率更高。因此, 本研究主要关注“中高适生区”的中适生区和高适生区。
2 结果与分析 2.1 模型精度验证由图 2可知, 预测分布结果的ROC曲线下面积(AUC)的训练集和测试集的均值为0.898, 这表明MaxEnt模型预测得到的青藏高原藏羚羊的潜在分布区具有很好的精度, 其预测结果可靠度高, 不具有随机性。

|
| 图 2 MaxEnt模型的受试者操作特性曲线 Fig. 2 Receiver operating handling characteristic curves in MaxEnt model |
通过Jackknife模块对各气候因子的贡献率进行评估可知, 温度季节性变化(Bio4)训练得分值为最高, 贡献率为37.2%, 其次是海拔, 贡献率为17.9%, 随后是最干月降雨(Bio14), 贡献率是15%。坡度, 最干季降雨(Bio17), 最湿季降雨(Bio16)贡献率分别是10.7%, 8.9%, 8.3%。而温度年变幅(Bio7), 最干季均温(Bio9), 坡向贡献率较小。这表明最干月降雨(Bio4)和海拔是影响藏羚羊分布的主导环境因子(图 3)。

|
| 图 3 刀切法检测环境因子对藏羚羊在青藏高原分布的重要程度 Fig. 3 Jackknife test for evaluating the relative importance of environmental variables for Tibetan antelope on the QTP |
藏羚羊在青藏高原的中高适生面积为6.11×105 km2, 占青藏高原总面积的24.44%, 其中适生区面积为2.70×105 km2, 占10.80%, 高适生区面积为3.41×105 km2, 占13.64%。中高适生区主要是在30.38°N—37.30°N、74.38°E—82.31°E之间, 主要分布在羌塘高原的东部, 可可西里, 阿尔金山南侧且一直延伸到三江源国家公园附近, 在昆仑山中部也有部分分布。中高适生区几乎涵盖整个可可西里国家级自然保护区, 也分布在羌塘国家级自然保护区的东南部分, 新疆阿尔金山国家级自然保护区和三江源国家公园的北部也有大面积的分布;中高适生区主要分布在西藏的北部, 青海的西部和新疆的东南部。中适生区在新疆中昆仑自然保护区有部分点状分布, 祁连山西侧也有少部分的中适生区分布(图 4)。青藏高原的国家级和省级保护区保护了青藏高原藏羚羊中高适生区的56.92%, 而剩下43.08%的中高适生区约2.70×105 km2在保护区外, 未受到有效保护。未受到保护的中高适生区主要围绕在羌塘国家级自然保护区、可可西里国家级自然保护区与三江源国家公园的周围区域。另外, 在青海湖周围区域也存在部分保护空缺区。

|
| 图 4 当前气候情景下藏羚羊适宜生境分布 Fig. 4 Suitable habitat distributions of Tibetan antelope under the current climatic scenario |
通过叠加分析可知, 在SSP126, SSP245, SSP585三种压迫情景下, 中高适宜区向高纬度和高海拔转移趋势的明显。栖息地的受影响程度较小。藏羚羊的核心分布区域(中高适生区)仍然以西藏的中北部, 青海的西部和新疆的东南部为主。从时间尺度上看, 在三种不同压迫情景下, 中高适生区的变化趋势基本一致, 在整体上呈现出西南部分减少, 东北部分增加的趋势, 特别是在未来的T4和T5中高适生区的面积变化明显。
与当前气候条件相比, T2阶段的(SSP126, SSP245, SSP585)的情境下, 三种情景下藏羚羊的中高适生区新增分别是6.72×104 km2, 6.75×104 km2, 6.53×104 km2;丧失的中高适生区有6.20×104 km2, 6.19×104 km2, 5.69×104 km2。丧失的中高适生区主要主要分布在羌塘保护区西南边界处, 新疆中昆仑自然保护区的西部和中部, 和三江源国家公园西南缘与羌塘保护区交界处。新增的中高适生区主要在阿尔金山国家级自然保护区的北部, 三江源国家公园的中北部在保护区外围也有大量零星分布。青海湖附近区域也有少量新增的藏羚羊中高适宜区。
与当前气候条件相比, T4阶段的(SSP126, SSP245, SSP585)的情境中, 三种情景下藏羚羊的中高适生区丧失的面积分别是7.68×104 km2, 6.16×104 km2, 7.99×104 km2;新增的中高适生区有6.59×104 km2, 7.82×104 km2, 7.29×104 km2。丧失的中高适生区主要是在丧失的中高适生区也呈现出点状零星分布。随着强迫程度的增加, 中高适生区丧失的面积也会增加。T5阶段的SSP585丧失中高适宜区面积最多, 达到9.22×104 km2, 丧失的中高适生区呈现出部分面状分布, 如新疆中昆仑自然保护区中部及南部, 羌塘高原的南部边缘, 新疆的南部的边缘, 西藏的北部, 祁连山的西南部, 青海湖西北部和三江源国家公园的中北部。羌塘自然保护区的西北部出现较多低适宜区转变为中高适宜区的区域而可可西里国家级自然保护区中高适生区对于气候变化的响应程度略低。

|
| 图 5 不同时期不同气候变化情景下藏羚羊适宜生境变化 Fig. 5 Suitable habitat changes of Tibetan antelope under future climatic scenarios in the T2, T3, T4 and T5 periods SSP85、SSP245、SSP126代表气候变化情景 |
在三种气候情景下, 保护空缺面积整体减小幅度不明显。青藏高原西南缘的保护空缺面积会随气候变化减少, 青藏高原东北部分出现新的适宜栖息地。新疆中昆仑自治区级保护区在SSP585的压迫情景下栖息地面积明显减少(图 6)。羌塘国家级自然保护区的南缘部分保护空缺将向更高纬度扩展。SSP3245情景下, 保护空缺的面积受气候变化的影响最小。

|
| 图 6 藏羚羊生境潜在分布与保护空缺分布 Fig. 6 Potential distributions of habitats and protection gaps of Tibetan antelope |
本研究发现, 气温和海拔是对藏羚羊适生区分布的贡献率最高的两个生物气候因子。其中, 温度季节性变化(Bio4)和海拔总贡献率达到55.1%。过去的研究发现藏羚羊喜好生活在全年3/4以上的月份平均气温不足0℃, 海拔介于3250—5500 m的高寒荒漠地区, 本次研究很好地验证了这一点。气温上升会影响野生动物的生理行为, 例如, 显著的温度升高和气温的变化可能会引起热应激并影响适应高山地区物种的生命率[35]。温度升高可能会限制物种的分布, 因为它会限制觅食等重要活动的可用时间, 或者限制基因交流所需的时间[36—37]。藏羚羊的适宜生境的变化也同多数前人的研究一致, 随着气候变暖, 其分布范围将逐渐缩小, 并向高纬度和高海拔转移[38]。本研究结果同样表明青藏高原西南部部分地区的海拔和气候条件已经不满足藏羚羊生存, 藏羚羊可能会为了躲避低海拔及低纬度生境的湿热化, 往高海拔和高纬度地方迁移。藏羚羊损失的适宜生境的主要出现在青藏高原西南部, 和中北部的低海拔地区;新出现的适宜区域整体上集中于青藏高原面的东部和东北部地区, 整体表现出向青藏高原东部和东北部迁移的趋势。另外, 藏羚羊适宜生境还表现出向核心地带缩减的趋势[23](图 7)。而在同一时期, 藏羚羊适宜生境的面积并没有随强迫程度的增加, 而呈现出一定的规律性。本研究针对此问题做出了两种假设: (1)文中所用的基于CMIP6模式的未来气候数据的准确性和完整性有待于提高, CMIP6模式虽然拥有更精确的空间分辨率、改进的云微物理过程参数化方案和更复杂的地球系统过程并在未来气候模拟采用新的情景框架, 但是CMIP6模式对青藏高原降水和气温的模拟能力仍然有限, 对于青藏高原这样的复杂地形区域, 气候模式的水平分辨率仍较为粗糙, 冷偏差与湿偏差依然存在, 并且偏差会随着时间的推移而变大[39];(2)目前关于气候变化下高海拔的有蹄类动物的潜在范围变化研究还很少, 这可能限制了我们对气候变化有蹄类野生动物潜在影响的认识和对制定保护策略的指导。

|
| 图 7 未来80年藏羚羊适宜生境转换 Fig. 7 Suitable habitat conversion of Tibetan antelope in the next 80 years |
三江源国家公园、可可西里国家级自然保护区、羌塘国家级自然保护区、新疆阿尔金山国家级自然保护区和新疆中昆仑自治区级自然保护区虽保护了56.92%藏羚羊的中高适生区, 但仍有43.18%的保护空缺区在保护区外, 未受到有效保护。且在低适生区内仍有大量零星的藏羚羊中高适宜生境。尽管藏羚羊在世界自然保护联盟(IUCN)红色名录中的保护等级已经下降, 但在气候变化的影响下, 藏羚羊的灭绝率仍将有上升的风险[24]。对于藏羚羊适宜生境丧失严重性、藏羚羊的分布的复杂性以及当前保护空缺的急迫性。建议相关部门在以国家公园为主体的自然保护地体系优化的大背景下, 综合考虑藏羚羊及其他有蹄类野生动物的生物学特性, 调整保护边界, 并在保护地之间布设生态廊道, 保障藏羚羊等濒危有蹄类野生动物栖息地完整性和连续性。
4 结论本文利用386个藏羚羊实际分布点位数据和9个环境变量数据, 采用最大熵模型(MaxEnt)模拟青藏高原地区藏羚羊当前分布格局以及气候变化背景下青藏高原藏羚羊生境时空格局演变, 并得到影响其分布的主导环境变量主要是温度季节性变化(Bio4), 海拔, 最干月降雨(Bio14)。通过空间叠加分析, 得到不同情景下的中高适生区和保护空缺分布。在三种气候情境下藏羚羊的适宜生境都具有向高纬度、高海拔迁移趋势。在五个自然保护地外仍然有近44%的中高适生区未受到保护。将不同气候情景下的适宜生境有效联系, 综合考虑藏羚羊迁徙廊道, 采取相应的保护措施, 如扩建和增建自然保护地、实施长期监测以获得藏羚羊转移和分布趋势的有效信息、加强畜牧储备平衡等。建议在今后的自然保护地管理中应对藏羚羊保护空缺区域加强关注。
| [1] |
Lynn J and Peeva N, Peeva N. Communications in the IPCC's Sixth Assessment Report cycle. Climatic Change, 2021, 169(1/2): 18. |
| [2] |
Parmesan C, Yohe G. A globally coherent fingerprint of climate change impacts across natural systems. Nature, 2003, 421(6918): 37-42. DOI:10.1038/nature01286 |
| [3] |
Walther G R, Post E, Convey P, Menzel A, Parmesan C, Beebee T J C, Fromentin J M, Hoegh-Guldberg O, Bairlein F. Ecological responses to recent climate change. Nature, 2002, 416(6879): 389-395. DOI:10.1038/416389a |
| [4] |
黄萍, 徐卫华, 范馨悦, 韩梅. 青藏高原珍稀濒危哺乳动物保护优先区评价与空缺分析. 环境保护科学, 2022, 48(3): 1-6. |
| [5] |
马耀明, 胡泽勇, 田立德, 张凡, 段安民, 阳坤, 张镱锂, 杨永平. 青藏高原气候系统变化及其对东亚区域的影响与机制研究进展. 地球科学进展, 2014, 29(2): 207-215. |
| [6] |
朴世龙, 张宪洲, 汪涛, 梁尔源, 汪诗平, 朱军涛, 牛犇. 青藏高原生态系统对气候变化的响应及其反馈. 科学通报, 2019, 64(27): 2842-2855. |
| [7] |
Luo Z H, Jiang Z G, Tang S H. Impacts of climate change on distributions and diversity of ungulates on the Tibetan Plateau. Ecological Applications, 2015, 25(1): 24-38. DOI:10.1890/13-1499.1 |
| [8] |
Hole D G, Huntley B, Arinaitwe J, Butchart S H M, Collingham Y C, Fishpool L D C, Pain D J, Willis S G. Toward a management framework for networks of protected areas in the face of climate change. Conservation Biology, 2011, 25(2): 305-315. |
| [9] |
吴建国, 王亮, 杨永伟, 代拴发, 刘建泉, 朱高. 自然保护区还需面对气候变化挑战. 环境保护, 2011(4): 30-32. |
| [10] |
张佳怡, 伦玉蕊, 刘浏, 刘宇轩, 李鑫, 徐宗学. CMIP6多模式在青藏高原的适应性评估及未来气候变化预估. 北京师范大学学报: 自然科学版, 2022, 58(1): 77-89. |
| [11] |
Wilcove D S, Wikelski M. Going, going, gone: is animal migration disappearing. PLoS Biology, 2008, 6(7): e188. DOI:10.1371/journal.pbio.0060188 |
| [12] |
Sun C, Xu X D, Wang P J, Yang D, Zhang S J, Wang C Z, Cai W Y. The warming and wetting ecological environment changes over the Qinghai-Tibetan Plateau and the driving effect of the Asian Summer Monsoon. Journal of Tropical Meteorology, 2022, 28(1): 95-108. DOI:10.46267/j.1006-8775.2022.008 |
| [13] |
马燕, 格日力. 藏羚羊的研究现状. 中国高原医学与生物学杂志, 2017, 38(3): 206-212. |
| [14] |
An L, Shu W. International plenipotentiary conference to conclude an international convention on trade in certain species of wildlife: convention on international trade in endangered species of wild Fauna and Flora. American Journal of International Law, 1974, 68(1): 197-211. DOI:10.2307/2198862 |
| [15] |
Zhou H, Li D Q, Zhang Y G, Yang T, Liu Y. Genetic diversity of microsatellite DNA loci of Tibetan antelope (Chiru, Pantholops hodgsonii) in Hoh Xil National Nature Reserve, Qinghai, China. Journal of Genetics and Genomics, 2007, 34(7): 600-607. DOI:10.1016/S1673-8527(07)60068-X |
| [16] |
赵丹平. 中国故事|可可西里: 为藏羚羊"母亲之旅"护航. 新华社. (2021-09-08)[2002-03-06]. http://www.news.cn/video/2021-09/08/c_1211361497.htm.
|
| [17] |
诸葛海锦, 李晓文, 张翔, 高峰, 许东华. 青藏高原高寒荒漠区藏羚适宜生境识别及其保护状况评估. 应用生态学报, 2014, 25(12): 3483-3490. |
| [18] |
Lu S L, Chen F, Zhou J F, Hughes A C, Ma X Q, Gao W W. Cascading implications of a single climate change event for fragile ecosystems on the Qinghai-Tibetan Plateau. Ecosphere, 2020, 11(9): e03243. |
| [19] |
Phillips S J, Dudík M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161-175. |
| [20] |
Qin A L, Liu B, Guo Q S, Bussmann R W, Ma F Q, Jian Z J, Xu G X, Pei S X. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Global Ecology and Conservation, 2017, 10: 139-146. |
| [21] |
伊拉木江·托合塔洪, 阿迪力·艾合麦提, 单文娟, 布威海丽其姆·阿巴拜科日, 塔依尔江·麦麦提, 马合木提·哈力克. 新疆兔属三物种潜在生境分布及未来气候变化的影响. 野生动物学报, 2020, 41(1): 70-79. |
| [22] |
柳晓燕, 李秀山, 赵彩云, 李飞飞, 朱金方, 纪文婧. 气候变化背景下三尾凤蝶潜在适生区模拟及保护空缺分析. 环境昆虫学报, 2021, 43(5): 1168-1177. |
| [23] |
魏子谦, 徐增让. 羌塘高原藏羚羊栖息地分布及影响因素. 生态学报, 2020, 40(23): 8763-8772. |
| [24] |
晏婷婷, 冉江洪, 赵晨皓, 钟雪, 梁敏仪. 气候变化对邛崃山系大熊猫主食竹和栖息地分布的影响. 生态学报, 2017, 37(7): 2360-2367. |
| [25] |
Qiu J. China: the third pole. Nature, 2008, 454(7203): 393-396. |
| [26] |
Zhang J J, Jiang F, Li G Y, Qin W, Wu T, Xu F, Hou Y S, Song P F, Cai Z Y, Zhang T Z. The four antelope species on the Qinghai-Tibet plateau face habitat loss and redistribution to higher latitudes under climate change. Ecological Indicators, 2021, 123: 107337. |
| [27] |
高文强, 王小菲, 江泽平, 刘建锋. 气候变化下栓皮栎潜在地理分布格局及其主导气候因子. 生态学报, 2016, 36(14): 4475-4484. |
| [28] |
张微, 姜哲, 巩虎忠, 栾晓峰. 气候变化对东北濒危动物驼鹿潜在生境的影响. 生态学报, 2016, 36(7): 1815-1823. |
| [29] |
李宁宁, 张爱平, 张林, 王克清, 罗红燕, 潘开文. 气候变化下青藏高原两种云杉植物的潜在适生区预测. 植物研究, 2019, 39(3): 395-406. |
| [30] |
沈涛, 张霁, 申仕康, 赵艳丽, 王元忠. 西南地区红花龙胆分布格局模拟与气候变化影响评价. 应用生态学报, 2017, 28(8): 2499-2508. |
| [31] |
曹雪萍, 王婧如, 鲁松松, 张晓玮. 气候变化情景下基于最大熵模型的青海云杉潜在分布格局模拟. 生态学报, 2019, 39(14): 5232-5240. |
| [32] |
雷军成, 徐海根, 吴军, 关庆伟. 基于IPCC AR5的我国常绿阔叶林潜在适宜生境变化分析. 生态与农村环境学报, 2015, 31(1): 69-76. |
| [33] |
Guo Q H, Liu Y. ModEco: an integrated software package for ecological niche modeling. Ecography, 2010, 33(4): 637-642. |
| [34] |
Hansen P J. Effects of heat stress on mammalian reproduction. Philosophical Transactions of the Royal Society B: Biological Sciences, 2009, 364(1534): 3341-3350. |
| [35] |
Vasseur D A, DeLong J P, Gilbert B, Greig H S, Harley C D G, McCann K S, Savage V, Tunney T D, O'Connor M I. Increased temperature variation poses a greater risk to species than climate warming. Proceedings of the Royal Society B: Biological Sciences, 2014, 281(1779): 20132612. |
| [36] |
Conley K E, Porter W P. Heat loss from deer mice (Peromyscus): evaluation of seasonal limits to thermoregulation. Journal of Experimental Biology, 1986, 126(1): 249-269. |
| [37] |
Sinervo B, Miles D B, Martínez-Méndez N, Lara-Resendiz R, la Cruz F R M. Response to comment on "erosion of lizard diversity by climate change and altered thermal niches". Science, 2011, 332(6029): 537. |
| [38] |
雷军成, 徐海根, 吴军, 关庆伟, 丁晖, 崔鹏. 气候变化情景下物种适宜生境预测研究进展. 四川动物, 2015, 34(5): 794-800. |
| [39] |
陈炜, 姜大膀, 王晓欣. CMIP6模式对青藏高原气候的模拟能力评估与预估研究. 高原气象, 2021, 40(6): 1455-1469. |