 2024, Vol. 44
2024, Vol. 44文章信息
- 郭子良, 张余广, 刘魏魏, 张曼胤, 刘振杰
- GUO Ziliang, ZHANG Yuguang, LIU Weiwei, ZHANG Manyin, LIU Zhenjie
- 河北衡水湖自然保护区水鸟多样性分布格局及其保护优先区
- Distribution pattern and conservation priority areas of waterbird diversity in Hengshui Lake Nature Reserve, Hebei Province
- 生态学报. 2024, 44(9): 4009-4019
- Acta Ecologica Sinica. 2024, 44(9): 4009-4019
- http://dx.doi.org/10.20103/j.stxb.202307131509
-
文章历史
- 收稿日期: 2023-07-13
- 网络出版日期: 2024-02-01

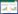




2. 中国林业科学研究院生态保护与修复研究所, 北京 100091;
3. 河北衡水湖湿地生态系统国家定位观测研究站, 衡水 053000;
4. 河北衡水湖国家级自然保护区管理委员会, 衡水 053000
2. Institute of Ecological Protection and Restoration Research, Chinese Academy of Forestry, Beijing 100091, China;
3. National Ecosystem Research Station of Hengshui Wetland, Hengshui 053000, China;
4. Administrative Committee of Hebei Hengshui Lake National Nature Reserve, Hengshui 053000, China
全球超过三分之一的湿地已经消失, 湿地及其生物多样性保护面临巨大挑战[1]。为了遏制野生动植物生境丧失、生物多样性减少, 建设了大量的自然保护地[2—3]。识别生物多样性空间分布格局及保护优先区, 是提升自然保护地管理成效、实现生物多样性保护的最有效途径之一[4—5]。而水鸟对环境变化具有极高的敏感性, 能指示生态系统健康水平、保护价值等[6—7]。将水鸟作为指示物种开展湿地保护优先区研究, 已广泛应用于自然保护地优化布局和建设管理[6, 8]。
生境破碎化导致的生境适宜性、人为干扰等变化干扰了鸟类栖息地选择, 改变了鸟类多样性[9—11]。水鸟多样性与湿地面积大小、食物量和生境质量等呈正相关关系, 与湿地围垦等人为干扰呈负相关关系[12—13]。目前人类活动增强、栖息地质量下降等成为影响水鸟群落结构和多样性的重要因素[14—15]。此外, 水位过度上升会导致生境类型趋于单一化, 水鸟群落从浅水类群为主转换为深水类群为主[16]。河北衡水湖是东亚-澳大利西亚水鸟迁徙路线的重要迁徙驿站, 面临着自然生境减少、多样性下降等问题[17—18]。生态保护和人们生产生活对土地需求之间的矛盾在衡水湖依然存在[17]。水鸟多样性时空格局及其保护优先区识别对自然保护区功能区划定、水鸟栖息地保护修复具有重要意义[19]。本研究使用GIS10.2软件和结构方程模型等对衡水湖自然保护区水鸟多样性分布格局及其保护优先区进行分析, 以期为水鸟及其栖息地保护管理提供科学支撑。
1 研究区概况河北衡水湖自然保护区(37°32′14″—37°41′25″N, 115°28′59″—115°41′40″E)位于河北省中南部衡水市境内, 总面积16365 hm2。属暖温带半湿润大陆性季风气候, 四季分明, 气温、降雨量和蒸发量等季节变化趋势明显[20], 生境类型多样, 水鸟种类丰富、季节性变化明显。自然保护区内鱼塘和农田面积较大, 主要位于中湖大道西侧, 即衡水湖西湖区域[17]; 自然保护区东部长期保持相对稳定的湖泊水面, 即衡水湖东湖, 一般也称为衡水湖(包括大湖和冀州小湖)。滏阳新河和滏东排河从自然保护区的北部穿过, 滏阳新河两侧为泄洪区。按照自然保护区内湿地空间分布和生境差异, 划分了5处湿地生境区域, 包括滏阳新河、滏东排河、衡水湖东湖、衡水湖西湖、冀码渠等, 见图 1。其中, 滏阳新河以河流湿地及周边泄洪区农田生境为主; 滏东排河以河流湿地及周边湿地芦苇沼泽生境为主, 包括北侧湿地恢复区; 衡水湖东湖以中湖大道东侧的湖泊水面、芦苇和香蒲沼泽生境为主; 衡水湖西湖以中湖大道西侧的鱼塘及周边农田生境为主; 冀码渠以河流湿地及周边农村坑塘生境为主。

|
| 图 1 研究范围及水鸟调查点示意图 Fig. 1 Schematic diagram of the research scope and survey sample sites of waterbirds |
2020年5月至2021年4月期间, 分别在2020年5月、7月、9月、10月、11月、12月, 以及2021年3月和4月进行衡水湖自然保护区水鸟群落的野外调查; 每次调查5—7d。在前期调查基础上, 划定公里网格, 将湿地区域作为主要调查样点, 根据自然保护区内湖泊、河流、沼泽和鱼塘等湿地, 以及桥梁和道路等分布, 布设调查样点开展水鸟调查, 记录不同调查区域距道路、居民点等干扰源的距离。按照《中国鸟类分类与分布名录(第四版)》[21]进行水鸟多样性编目, 确定水鸟的目、科、属、种, 及其在中国的分布情况。根据世界自然保护联盟(The International Union for Conservation of Nature, IUCN)的红色名录(https://www.iucnredlist.org/)、《国家重点保护野生动物名录(2021版)》和《河北省重点保护陆生动物名录》, 确定物种的濒危和保护等级。
2.2 数据处理将水鸟调查数据按照滏阳新河、滏东排河、衡水湖东湖、衡水湖西湖、冀码渠等5个区域进行整理, 分析不同区域水鸟多样性的差异。使用Shannon多样性指数、Jaccard相似系数分析水鸟多样性及其群落特征等[14]; 并使用生物多样性保护价值指数和不可替代性指数等开展保护优先区分析[8, 22]; 使用地理信息系统软件GIS10.2中的插值分析(反距离权重法)等分析自然保护区内水鸟多样性及其保护价值指数、不可替代性指数的地理空间格局。将水鸟多样性保护价值指数和不可替代性指数分别大于其最高值50%的范围的合并区域作为水鸟多样性保护优先区。
(1) Shannon多样性指数(H)计算公式如下:
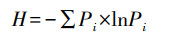
|
式中, Pi为第i种鸟类种群数量占鸟类种群总数量的比率。
(2) 水鸟群落组成相似性分析选择Jaccard相似系数(Cj)来度量, 比较不同区域水鸟组成的相似程度[14]。
(3) 根据不同区域水鸟的保护等级、濒危性和分布特征, 及其种群数量, 计算水鸟多样性保护价值指数(VA)[22—23]。
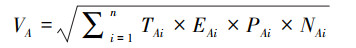
|
式中, n代表调查区域水鸟种类数量, TAi代表水鸟i的濒危性赋值, EAi代表水鸟i的特有性赋值, PAi代表水鸟i的保护等级赋值, NAi代表水鸟i的种群数量。VA值越高, 表示调查区域水鸟多样性保护价值越高。
(4) 通过不可替代性指数(I)分析评估区域的水鸟保护优先性[8]。不可替代性指数(I)计算公式如下:
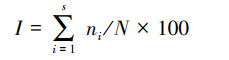
|
式中, ni代表调查区域第i个物种的种群数量, N代表衡水湖自然保护区第i个物种的总数量, S为衡水湖自然保护区调查记录的水鸟总种数。I值越高, 表示调查区域的保护优先级越高。
2.3 结构方程模型建立结构方程模型(Structural Equation Model)是基于变量的协方差矩阵分析变量之间关系的一种统计方法[24], 能有效描述潜变量及其外显指标之间的关系。该模型由测量方程和结构方程两部分组成。人为干扰、生境状况是影响水鸟食源、干扰强度的直接因素, 对水鸟栖息地选择起决定性作用[15]。调查区均位于衡水湖自然保护区内, 调查周期内衡水湖自然保护区的湿地范围、景观格局、道路和居民点等人为干扰基本稳定, 不同区域栖息地质量变化小[17]。基于以上考虑和数据的可获得性, 建立河北衡水湖湿地水鸟多样性与环境因子响应关系的概念模型。模型包括2个潜变量: 生境状况和人为干扰, 5个可测变量: 湿地占比、自然生境占比、距干扰源距离、人为干扰度、水鸟多样性。运用Amos 26.0进行结构方程模型分析, 采用x2/df、RMSEA、NFI、TFI、CFI、IFI、AIC、ECVI指征验证结果是否具有统计学意义。基于公里网格计算不同公里网格内的湿地占比、自然生境占比、人为干扰度[17], 反映调查区域及周边的栖息地状况、人为干扰, 并将其与调查样点的全年水鸟Shannon多样性指数进行匹配, 开展驱动因素分析。
其中, 距干扰源距离为调查样点距道路和居民点等干扰源的最近距离(m)。湿地占比和自然生境占比根据2016年8月的高分一号卫星(GF-1)遥感影像数据解译后的景观类型分布数据计算得到, 分别计算每个公里网格内湿地和自然生境所占比例[17]。人为干扰度使用人为干扰度指数(HDI)计算, 公式如下:
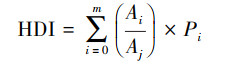
|
式中, m为景观类型的数量, Ai为采样网格中第i种景观类型的面积(hm2), Aj为采样网格总面积(hm2), Pi为第i种景观类型所反映的干扰度系数[25]。通过Delphi法咨询衡水湖工作的专家确定不同景观类型的干扰度系数, 即芦苇0.14、香蒲0.1、开阔水面0.33、莲0.4、鱼塘0.69、林地0.33、草地0.43、裸地0.56、果园0.7、苗圃0.56、耕地0.75、公园0.69、道路0.86、村庄0.86、城镇0.93。
3 结果 3.1 不同区域的水鸟群落结构差异滏东排河、衡水湖东湖是衡水湖自然保护区水鸟分布较为集中的区域, 水鸟多样性高、种群规模大, 见图 2。衡水湖东湖水鸟的种群数量明显高于其它区域, 累计水鸟数量达41104只。但衡水湖西湖和滏东排河水鸟的物种丰富度和Shannon多样性指数最高, 分别为56种和2.49。不同区域水鸟群落的优势类群多以雁鸭类游禽为主, 一般为中等相似程度(0.5<Cj<0.75), 见表 1。衡水湖东湖与滏阳新河、冀码渠的水鸟群落结构相似度低, 分别为0.46和0.49。衡水湖东湖的雁鸭类和鸥类等游禽种数占明显优势, 占总种数的60%左右; 而滏阳新河、冀码渠的鸻鹬类占其总种数的比例最高, 但丰富度不高。在种群数量上, 衡水湖东湖、衡水湖西湖、滏东排河和滏阳新河的雁鸭类种群数量较高, 均占其水鸟总数量的65%以上, 而冀码渠的鸊鷉目(Podicipediformes)等其它游禽种群数量最高, 占其水鸟总数量的43.38%。此外, 滏阳新河鸻鹬类种群数量明显高于其他区域, 为该区域种群数量第二高的类群。

|
| 图 2 不同区域的水鸟丰富度、多样性和种群数量 Fig. 2 Richness, diversity and individual number of waterbirds in different regions |
| 区域Region | 滏阳新河 | 滏东排河 | 冀码渠 | 衡水湖西湖 |
| 滏东排河Fudongpai river | 0.57 | |||
| 冀码渠Jima Qu | 0.51 | 0.62 | ||
| 衡水湖西湖West lake of Hengshui Lake | 0.56 | 0.62 | 0.54 | |
| 衡水湖东湖East lake of Hengshui Lake | 0.46 | 0.55 | 0.49 | 0.68 |
衡水湖自然保护区不同区域的游禽种数均表现为3月、11月、12月等月份较高, 5—9月份较低, 见图 3。除滏阳新河外, 其它区域在4—10月的游禽种数都十分接近, 而且滏阳新河游禽种数在不同时间均明显低于其它区域。滏东排河的游禽种数在11月最高, 随后12月衡水湖东湖游禽种数达到最高, 并持续到次年3月。此外, 滏东排河和衡水湖西湖的游禽种群数量在11月突然增加达到峰值; 而衡水湖东湖游禽种群数量则在12月达到峰值, 占全域游禽总数量的70%以上。

|
| 图 3 不同区域水鸟群落的季节性变化 Fig. 3 Seasonal variation of waterbird communities in different regions |
涉禽的种数和种群数量与该区域游禽的变化趋势正好相反, 3—5月不同区域的涉禽种数均逐渐增长, 并在4月或5月达到峰值。滏东排河、衡水湖西湖和东湖在4—5月的涉禽种数达到峰值后逐渐下降, 但滏阳新河的涉禽种数在5月和9月出现了两个峰值。但3—7月, 衡水湖不同区域的涉禽种群数量普遍呈增长趋势, 其中衡水湖东湖的增长最明显。9月之后, 涉禽种数和种群数量快速下降。
不同区域水鸟群落随时间不断改变, 4月开始各区域游禽种数和种群数量占比不断下降, 并在5—9月形成了以鹭科、鹬科等涉禽在种类组成上占优势的水鸟群落, 但滏东排河和冀码渠的鸭科等游禽种群数量占比仍很高。其中5月滏阳新河的游禽种数仅占其总种数的11.11%、种群数量仅占其总数量的27.30%;7月衡水湖东湖的游禽种群数量占其总数量的43.26%。9月之后各区域游禽种数和种群数量占比明显上升, 逐渐形成了以雁鸭类游禽占绝对优势的水鸟群落, 其种群数量普遍占各区域总数量的90%以上。
3.3 水鸟多样性分布格局及其驱动因素 3.3.1 水鸟多样性分布格局滏东排河及其北侧的湿地恢复区、衡水湖西湖鱼塘、衡水湖东湖部分岛屿的水鸟物种丰富度较高, 见图 4。但不同类群水鸟物种丰富度的空间分布格局存在较大差异。其中, 鸻鹬类和鹤鹳类物种丰富度较高的范围较小, 主要包括自然保护区北部的滏阳新河和滏东排河及周边区域。而鸥类和鹭类物种丰富度在不同区域较为接近, 空间聚集度低, 覆盖了衡水湖东湖和滏东排河的大部分区域, 特别是鹭类。

|
| 图 4 衡水湖自然保护区水鸟物种丰富度空间分布格局 Fig. 4 Spatial distribution pattern of waterbird species richness in Hengshui Lake Nature Reserve |
湿地恢复区及南侧的滏东排河、小湖隔堤两侧、衡水湖西湖鱼塘等地水鸟种群数量较大、聚集程度高, 见图 5。但雁鸭类、鸥类、鸻鹬类、鹭类和鹤鹳类等水鸟在衡水湖自然保护区的集群分布区域各异。雁鸭类主要聚集在小湖隔堤两侧及衡水湖西湖鱼塘和农田; 鸥类主要聚集在衡水湖东湖东北角和西南角; 鸻鹬类主要聚集在北部滏阳新河和滏东排河的部分河段; 鹭类则主要聚集在衡水湖东湖东北角以及中湖大道两侧; 鹤鹳类空间分布最集中, 主要分布在中湖大道西侧农田等区域。

|
| 图 5 衡水湖自然保护区水鸟种群数量空间分布格局 Fig. 5 Spatial distribution pattern of waterbird individual quantity in Hengshui Lake Nature Reserve |
河北衡水湖湿地水鸟多样性与环境因子的标准化系数修正模型见图 6;拟合指数总体表现较好, 均达到拟合指数要求, 见表 2。栖息地状况和人为干扰对水鸟多样性的直接影响分别为0.63和0.44, 共同改变了衡水湖湿地水鸟多样性。而且栖息地状况和人为干扰的相关系数为-1.05, 人为干扰显著改变了衡水湖湿地栖息地状况。此外, 自然生境占比、湿地占比两个可测变量对栖息地状况潜变量的影响路径系数分别为0.98和0.97, 影响较大。

|
| 图 6 河北衡水湖湿地水鸟多样性与环境因子响应关系修正模型 Fig. 6 Correction model for the relationship between waterbird diversity and environmental factors in Hengshui Lake wetland, Hebei e1—e7:残差Errors |
| 指数名称 Indices name |
参考标准 Evaluation criterion |
评价结果 Results |
|
| 绝对拟合指数Absolute fit index | x2/df | <3 | 1.623 |
| GFI | >0.9, 越接近1越好 | 0.980 | |
| RMSEA | <0.08 | 0.079 | |
| 相对拟合指数Relative fit index | NFI | >0.9, 越接近1越好 | 0.991 |
| TLI | >0.9, 越接近1越好 | 0.989 | |
| CFI | >0.9, 越接近1越好 | 0.997 | |
| 精简指数Compact index | AIC | 越小越好 | 28.870 |
| ECVI | 越小越好 | 0.295 | |
| x2/df: 卡方自由度比, GFI: 拟合优度指数Goodness of fit index, RMSEA: 近似误差均方根Root mean square error of approximation, NFI: 赋范拟合指数Normed fit index, TLI: 塔克-刘易斯指数Tucker-lewis index, CFI: 比较拟合指数Comparative fit index, AIC: 讯息效标系数Akaike information criterion, ECVI: 预期交叉验证指数Expected cross-validation index | |||
衡水湖自然保护区水鸟多样性保护价值指数和不可替代性指数分别在6.63—212.60和0.39—689.69, 不同区域差异较大, 见图 7。水鸟多样性保护价值较高的区域较为分散, 主要包括东北部的滏东排河和滏阳新河, 西湖的良心庄鱼塘和南李庄鱼塘, 东湖的小湖隔堤两侧。而水鸟不可替代性指数较高的区域范围较小且集中, 主要位于自然保护区北部滏东排河及两侧, 包括南李庄鱼塘、良心庄鱼塘、湿地恢复区及周边区域、东湖的姊妹东岛等地。综合水鸟多样性保护价值指数和不可替代性指数, 衡水湖自然保护区的水鸟保护优先区包括湿地恢复区及周边区域、南李庄鱼塘、良心庄鱼塘及周边区域、东湖的姊妹东岛、小湖隔堤两侧等5个区域, 总面积约1462.90hm2, 占保护区总面积的8.94%, 见图 8。

|
| 图 7 衡水湖自然保护区不同区域的水鸟多样性保护价值指数和不可替代性指数 Fig. 7 Conservation value index and irreplaceable index of waterbird diversity in different areas of Hengshui Lake Nature Reserve |

|
| 图 8 衡水湖自然保护区的水鸟保护优先区 Fig. 8 Conservation priority areas for waterbird in Hengshui Lake Nature Reserve |
衡水湖自然保护区表现为以雁鸭类等深水类群为主的水鸟群落特征。鸟类群落结构及其多样性受景观类型、隐蔽场所、人为干扰等多种因素的影响[26—27]。自然保护区内不同区域水鸟群落结构及其多样性表现出一定的差异。其中, 衡水湖西湖以鱼塘和农田生境为主, 形成以雁鸭类等游禽在数量上占绝对优势的水鸟群落, 但鱼塘水位的季节性波动使其鸻鹬类种数最高但种群数量少[17]。而衡水湖东湖长期保持相对稳定的高水位, 为雁形目(Anseriformes)、鸊鷉目、鸥科(Laridae)等各类游禽提供了适宜生境, 其种类多、种群数量大; 同时苍鹭(Ardea cinerea)、白鹭(Egretta garzetta)、池鹭(Ardeola bacchus)等鹭科鸟类种群数量大。但衡水湖东湖浅滩、浅水区等湿地生境较少, 鸻鹬类种群数量占比很小, 记录到的鸻鹬类仅少量出现在姊妹东岛等岛屿浅滩。滏阳新河和滏东排河不同月份水位波动大, 形成了不同水深条件的湿地生境, 除雁鸭类外仍有较多鸻鹬类等涉禽栖息, 涉禽种数和种群数量占比相对较高。但与历史数据相比, 衡水湖自然保护区内鸻鹬类等涉禽种数和种群数量明显降低[18, 28]。
衡水湖自然保护区的水鸟群落结构具有明显的季节性变化规律, 不同区域的涉禽和游禽物种丰富度、种群数量高峰期在时间上不交叉。3—7月苍鹭、白鹭、池鹭、夜鹭(Nycticorax nycticorax)等鹭类陆续在衡水湖东湖的岛屿和蒲草区繁殖, 同时大量鸻鹬类在滏东排河和滏阳新河停歇觅食。衡水湖自然保护区的涉禽种类和种群数量持续增加并形成优势种群, 特别是在东湖。而在秋冬季, 绿头鸭(Anas platyrhynchos)、绿翅鸭(Anas crecca)、豆雁(Anser fabalis)等雁鸭类在衡水湖集群停歇或越冬, 种类和种群数量逐渐达到峰值, 不同区域均形成了以雁鸭类为优势种群的水鸟群落; 而且在迁徙季衡水湖东湖白骨顶(Fulica atra)集群数量达千只以上, 这与华北很多区域的水鸟迁徙规律相似[29]。12月大量豆雁在衡水湖集群越冬或迁徙停歇, 成为优势种群, 衡水湖东湖和西湖的游禽数量明显增加。在衡水湖自然保护区内深秋至冬季越冬期豆雁主要在衡水湖西湖农田区域觅食, 并在湿地区域栖息和饮水。
游禽和涉禽虽然都依赖于湿地生存, 但不同类群水鸟对主要环境变量的响应存在明显差异[30]。生态补水、农业生产等人为干扰直接改变了不同区域湿地的水深、生境状况等, 一定程度上决定了衡水湖水鸟多样性时空格局[31—32]。衡水湖水鸟物种丰富度较高的区域集中在湿地恢复区及周边、衡水湖西湖部分鱼塘、衡水湖东北角的岛屿。而湿地恢复区、小湖隔堤两侧、良心庄鱼塘水鸟种群规模大, 形成了水鸟聚集分布中心。但不同类群水鸟分布格局有很大差异, 其中鸻鹬类、鹤鹳类等涉禽主要位于衡水湖北部滏东排河、滏阳新河及周边, 雁鸭类主要位于小湖隔堤两侧、衡水湖西湖鱼塘及周边。鸟类多样性是众多可测变量和潜变量之间相互作用的结果, 结构方程模型可以定量描述水鸟多样性与环境因子间的复杂关系[12]。人为干扰、栖息地状况对水鸟多样性都产生了直接影响, 其中栖息地状况的直接影响更大。而人为干扰通过改变栖息地状况对其产生间接影响, 具有显著的负向作用。强化自然保护区建设管理, 控制人类活动, 增加自然生境和湿地占比, 能维持并提升水鸟多样性。本文结构方程模型根据可获得现状变量数据构建, 反映研究期内水鸟多样性与已选变量的关系。对不同时期水鸟多样性与环境因子之间的关系仍需补充完善湿地水深、食物密度、人类活动等相关变量数据, 从而提高结论的科学性和准确性。
在衡水湖自然保护区北部的滏东排河及两侧、南李庄鱼塘、良心庄鱼塘等区域水鸟多样性保护价值和不可替代性均较高。衡水湖水鸟多样性保护价值和不可替代性较高的区域具有一定的空间重叠, 而且其与水鸟的主要繁殖区和越冬区较为吻合。其中, 南李庄鱼塘是衡水湖青头潜鸭(Aythya baeri)的主要越冬区和迁徙停歇区之一、灰翅浮鸥(Chlidonias hybrida)繁殖区之一; 良心庄鱼塘是衡水湖鸻鹬类的主要繁殖区和雁鸭类的主要越冬区之一。衡水湖水鸟栖息地的保护优先区主要包括湿地恢复区及周边区域、南李庄鱼塘、良心庄鱼塘及周边区域、东湖的姊妹东岛、小湖隔堤两侧等5个区域, 其保护管理对维护衡水湖水鸟繁殖和越冬的栖息地安全、提升水鸟多样性具有关键作用。建议基于此优化自然保护区的功能分区、制定保护管理计划, 开展衡水湖湿地水鸟栖息地保护修复。本研究调查范围大, 且受新冠疫情等影响全年调查次数受限, 水鸟调查数据积累可能存在不足。
| [1] |
Hu S J, Niu Z G, Chen Y F, Li L F, Zhang H Y. Global wetlands: potential distribution, wetland loss, and status. Science of the Total Environment, 2017, 586: 319-327. DOI:10.1016/j.scitotenv.2017.02.001 |
| [2] |
Mcdonald R I, Boucher T M. Global development and the future of the protected area strategy. Biological Conservation, 2011, 144(1): 383-392. DOI:10.1016/j.biocon.2010.09.016 |
| [3] |
Guo Z L, Liu W W, Zhang M Y, Zhang Y G, Li X Y. Transforming the wetland conservation system in China. Marine and Freshwater Research, 2020, 71(11): 1469-1477. DOI:10.1071/MF19383 |
| [4] |
Brooks T M, Mittermeier R A, da Fonseca G A B, Gerlach J, Hoffmann M, Lamoreux J F, Mittermeier C G, Pilgrim J D, Rodrigues A S L. Global biodiversity conservation priorities. Science, 2006, 313(5783): 58-61. DOI:10.1126/science.1127609 |
| [5] |
马星, 王浩, 余蔚, 杜勇, 梁健超, 胡慧建, 邱胜荣, 刘璐. 基于MaxEnt模型分析广东省鸟类多样性热点分布及保护空缺. 生物多样性, 2021, 29(8): 1097-1107. |
| [6] |
孙锐, 崔国发, 雷霆, 郑姚闽. 湿地自然保护区保护价值评价方法. 生态学报, 2013, 33(6): 1952-1963. |
| [7] |
Padoa-Schioppa E, Baietto M, Massa R, Bottoni L. Bird communities as bioindicators: the focal species concept in agricultural landscapes. Ecological Indicators, 2006, 6(1): 83-93. DOI:10.1016/j.ecolind.2005.08.006 |
| [8] |
Xia S X, Yu X B, Millington S, Liu Y, Jia Y F, Wang L Z, Hou X Y, Jiang L G. Identifying priority sites and gaps for the conservation of migratory waterbirds in China's coastal wetlands. Biological Conservation, 2017, 210: 72-82. |
| [9] |
Luo K, Wu Z L, Bai H T, Wang Z J. Bird diversity and waterbird habitat preferences in relation to wetland restoration at Dianchi Lake, south-west China. 鸟类学研究: 英文版, 2019, 10(3): 374-385. |
| [10] |
邓文洪. 栖息地破碎化与鸟类生存. 生态学报, 2009, 29(6): 3181-3187. |
| [11] |
赵坤明, 陈圣宾, 杨锡福. 基于红外相机技术调查四川都江堰破碎化森林鸟兽多样性及优势种活动节律. 生物多样性, 2023, 31(6): 72-84. |
| [12] |
张永, 施慧, 刘璐婷, 沈伟, 赵梓羲. 江苏沿江湖泊湿地越冬水鸟多样性及其影响因素. 湖泊科学, 2022, 34(6): 2005-2019. |
| [13] |
付梦娣, 贾强, 任月恒, 周汉昌, 李俊生, 张渊媛, 白加德, 陈克林. 环渤海滨海湿地鸻鹬类水鸟多样性及其环境影响因子. 生态学报, 2021, 41(22): 8882-8891. |
| [14] |
洪咏怡, 卢训令, 赵海鹏. 黄淮平原农业景观不同生境鸟类多样性特征及年际动态. 生态学报, 2021, 41(5): 2045-2055. |
| [15] |
Rico-Silva J F, Cruz-Trujillo E J, Colorado Z G J. Influence of environmental factors on bird diversity in greenspaces in an Amazonian city. Urban Ecosystems, 2021, 24(2): 365-374. DOI:10.1007/s11252-020-01042-x |
| [16] |
Liang W E, Lei J L, Ren B S, Cao R X, Yang Z X, Wu N R, Jia Y F. The impacts of a large water transfer project on a waterbird community in the receiving dam: a case study of Miyun Reservoir, China. Remote Sensing, 2022, 14(2): 417. DOI:10.3390/rs14020417 |
| [17] |
郭子良, 张曼胤, 刘魏魏, 刘振杰, 张余广. 三个时期河北衡水湖国家级自然保护区景观格局和保护成效分析. 湿地科学, 2021, 19(2): 170-177. |
| [18] |
郭子良, 张余广, 刘魏魏, 蔡乐涵, 张曼胤, 刘振杰, 郭华兵. 河北衡水湖国家级自然保护区水鸟群落特征及其季节性变化. 生态学杂志, 2022, 41(4): 732-740. |
| [19] |
段后浪, 于秀波. 黄河流域水鸟保护优先区及空缺. 生态学报, 2023, 43(20): 8663-8670. |
| [20] |
吴景峰. 衡水市蒸发特性及干旱指数分析. 水科学与工程技术, 2018(5): 17-19. |
| [21] |
郑光美. 中国鸟类分类与分布名录. 4版. 北京: 科学出版社, 2023.
|
| [22] |
郭子良, 邢韶华, 崔国发. 自然保护区物种多样性保护价值评价方法. 生物多样性, 2017, 25(3): 312-324. |
| [23] |
国家林业局. 自然保护区生物多样性保护价值评估技术规程. 北京: 中国标准出版社, 2017.
|
| [24] |
李慧, 汪景宽, 裴久渤, 李双异. 基于结构方程模型的东北地区主要旱田土壤有机碳平衡关系研究. 生态学报, 2015, 35(2): 517-525. |
| [25] |
孙永光, 赵冬至, 吴涛, 卫宝泉, 高树刚, 李媛, 曹芳芳. 河口湿地人为干扰度时空动态及景观响应——以大洋河口为例. 生态学报, 2012, 32(12): 3645-3655. |
| [26] |
Shwartz A, Turbé A, Simon L, Julliard R. Enhancing urban biodiversity and its influence on city-dwellers: an experiment. Biological Conservation, 2014, 171: 82-90. DOI:10.1016/j.biocon.2014.01.009 |
| [27] |
梁凤, 周立志. 环巢湖湿地公园鸟类多样性的空间变化. 生态学杂志, 2023, 42(7): 1687-1698. |
| [28] |
蒋志刚. 衡水湖国家级自然保护区生物多样性. 北京: 中国林业出版社, 2009: 63-91.
|
| [29] |
柴硕, 曹子祯, 孙浩, 吴渊, 刘亚东, 王圣杰, 吕姝皇, 洪剑明. 河北怀来官厅水库国家湿地公园的水鸟多样性. 湿地科学, 2019, 17(5): 544-552. |
| [30] |
王成, 刘红玉, 李玉凤, 王刚, 董斌, 陈浩, 张亚楠, 赵永强. 盐城滨海湿地水鸟类群生境适宜性及生态阈值研究: 对栖息地景观结构恢复的启示. 生态与农村环境学报, 2022, 38(7): 897-908. |
| [31] |
Chastant J E, Gawlik D E. Water level fluctuations influence wading bird prey availability and nesting in a subtropical lake ecosystem. Waterbirds, 2018, 41(1): 35-45. |
| [32] |
Zhu Y W, Wang H X, Guo W X. The impacts of water level fluctuations of East Dongting Lake on habitat suitability of migratory birds. Ecological Indicators, 2021, 132: 108277. |