 2024, Vol. 44
2024, Vol. 44文章信息
- 杜志勇, 丛楠
- DU Zhiyong, CONG Nan
- 植被与土壤特征对青藏高原不同程度退化草地的响应
- Responses of vegetation and soil characterisitics to degraded grassland under different degrees on the Qinghai-Tibet Plateau
- 生态学报. 2024, 44(6): 2504-2516
- Acta Ecologica Sinica. 2024, 44(6): 2504-2516
- http://dx.doi.org/10.20103/j.stxb.202303310628
-
文章历史
- 收稿日期: 2023-03-31
- 网络出版日期: 2023-11-02





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
草地是陆地面积最大、分布最广的生态系统之一, 约占地球陆地面积的24%[1]。21世纪以来, 我国有90%的天然草地处于不同程度的退化之中[2], 导致生态环境遭到了严重的破坏。青藏高原是世界上最大的高寒草地分布区, 面积约为1.5×106km2, 占我国天然草地面积的30%—40%[3], 而高寒草地面积为1.27×108km2, 约占青藏高原总面积的一半。由于其独特的地理环境, 青藏高原草地在气候调节、水源涵养、生物多样性保护、碳汇和水土保持等方面发挥着重要的作用[4]。该地区气候严酷寒冷, 是一个典型的气候变化敏感区和生态脆弱区[5]。而草地对气候变化的响应尤为突出, 其具体变现为牧草生长季短, 植物群落结构简单, 抗逆性弱、恢复力差[6]。近年来, 受全球气候变化与人类活动的持续影响[7], 加之草畜的不当匹配和不合理的放牧管理方式[6], 导致高寒草地严重超载, 大面积的优良草地退化成为裸土, 其中退化草地面积已达到4.5×107km2[8]。伴随着草地退化日趋严重, 植被生产力随之逐渐降低, 土壤趋于贫瘠化与干旱化。生物多样性剧变, 草地生产力水平急剧下降, 群落物种组成改变, 优良牧草竞争、更新能力减弱, 毒杂草比例增加;草畜矛盾加重, 畜牧业经济效益下降, 鼠虫害严重, 景观破碎化加剧, 致使高寒草地生态系统的结构和功能严重受损并退化[9—12]。总的来看, 随着草地退化的加剧, 草地生态系统服务功能下降, 植物群落发生逆向演替, 朝着不利于恢复的方向进行。
草地退化是多种主导因素共同作用的结果[13], 许多学者认为高寒草地退化的主要原因是过度放牧和气候变化。在过去30多年间, 有关学者认为青藏高原高寒草地呈现整体向好、局部退化的态势[14—15]。以气候变化为主要驱动因子影响着青藏高原植被生产力, 呈现总体增加、局部下降的趋势[16];同时, 脆弱的草地生态系统内部驱动因素与过度放牧等外部干扰的整合也会造成草地退化[17]。此外, 通过分析遥感数据, 发现过度放牧对青藏高原高寒草地退化的贡献比19.90%, 远低于气候变化对草地退化的贡献比56.74%[7]。与此相反, 气候变化对草地退化的贡献率远低于人类活动以及两者共同主导的贡献率[18]。除气候变化外, 过度放牧也是进一步导致高寒草地退化的重要原因。过度放牧减少优良牧草的种类, 增加毒害草的种类, 缩小植物的叶片面积, 显著降低了植物的光合效率, 牧草养分严重匮乏, 抑制了植物的生长发育, 退化草地的开阔条件使得鼠、虫害加剧, 加快草地生境进一步趋于恶化的速率, 植物群落进行逆向演替[19—20]。因此, 气候变化通过改变青藏高原的水热条件, 进而加剧高寒草地生态系统退化;过度放牧降低土壤养分含量并破坏土壤质地, 降低植被生产力, 改变植被群落组成, 植被-土壤系统协同退化, 使草地退化陷入了恶性循环。
草地退化具体表现在植被退化和土壤退化两个方面, 植被退化表现为植被生产力、牧草质量、覆盖度和高度下降;土壤退化表现为土壤理化性质和微生物特性的减弱不利于植被的生长[21]。在植被退化方面, 禾本科、莎草科植物逐渐变少, 原生植被的群落优势种逐渐演变为杂类草[22—24];植被覆盖度、生产力和牧草质量逐渐下降, 生物多样性改变[25]。在土壤退化方面, 草地土壤退化滞后于植被退化[26], 其退化恢复时间要远远长于草地植被的恢复时间[27—28]。在草地退化的演替过程中, 植被生产指标和土壤养分含量明显降低[29], 土壤愈发贫瘠[25], 表层土壤理化性质对草地退化的响应程度比深层土壤更加敏感[30]。综上, 草地退化不仅表现为直观的植被退化, 还表现为潜在的土壤退化, 二者相互作用, 相互影响。植被动态变化及分布特征可以用来评估自然因素和人为干扰引起的土壤变化, 而土壤退化导致营养元素不足以提供植物正常的生长发育, 植被生产力和覆盖度等逐步下降, 土壤表层腐殖质的来源减少, 回归的土壤养分含量降低, 植被-土壤协同作用减弱, 这些都将导致土壤进一步恶化。
针对青藏高原草地退化状况, 我国学者已经开展了大量研究, 主要集中于土壤退化现状及其驱动力、植物群落生物量变化、土壤碳库变化等[31—32]。但是, 已有的青藏高原草地研究, 由于数据及方法的不同, 且研究草地类型的不同, 加上各自不同的时空分布特征, 导致结论差异较大, 甚至一些研究结果出现相互矛盾。邵建翔等[33]研究发现, 随着退化程度的加剧, 土壤pH在高寒草甸逐渐上升, 而在高寒草原却逐步下降;张骞等[21]总结青藏高寒区各生态类型分区草地退化现状, 描述各生态区的草地类型、植物群落组成、退化原因及对应恢复措施。同时, 研究范围大多局限于单一区域, 对于多区域、多类型的对比研究相对较少, 导致对高寒退化草地的植被和土壤变化特征仍缺乏统一的认识[34]。Meta分析是用于比较和综合针对同一科学问题研究结果的统计学方法, 通过整合相关研究的数据, 得出这一科学问题的普遍规律。因此, 本研究采用meta分析方法, 收集了2004—2022年间取得的研究结果, 对青藏高原不同退化程度高寒草地植被与土壤特征的变化进行系统分析, 旨在揭示草地退化过程中植被与土壤特征的变化规律, 为高寒退化草地恢复重建, 以及为建立稳定的高寒草地生态系统提供科学依据。
1 材料与方法 1.1 数据搜集本文通过中国知网、百度学术和Web of Science等中英文专业学术数据库, 以“青藏高原”、“退化草地”和“Qinghai-Tibet Plateau”、“Grassland degradation”为关键词搜集2004—2022年的相关文献, 为降低文献偏倚, 按照如下标准进行文章筛选和数据提取:(1)研究对象为青藏高原草地, 试验均在生长季开展, 试验样点生态环境相近;(2)试验包括对照组和至少一个退化处理组;(3)不同退化程度按照论文设定标准, 提取数据, 且文献图表数据清晰;(4)土壤深度不低于5cm, 当文献出现不同土壤深度处理组时, 选取最表层的数据;最终共纳入64篇文献的1403组数据。为比较草地退化对土壤理化性质及植被属性的影响, 本研究共选择了16个评价指标作为研究对象(表 1)。同时, 区分整理不同退化处理的高寒草地生态系统环境因素、土壤理化性质和植被属性。对应研究样地, 记录试验样点植被盖度(Vegetation coverage)、经度(Longitude)、纬度(Latitude)、海拔(Altitude)、年平均降水量(Mean annual precipitation)(图 1)以及年平均温度(Mean annual temperature)(图 2), 绘制研究区域分布图(高程数据来源于中国青藏高原数据中心http://data.tpdc.ac.cn)[35]。由图可见, 研究区域内的温度和降水趋于同频变化, 青藏高原的气候总体呈现暖湿化趋势, 局部地区受暖干化影响, 主要表现为东部地区成暖湿化分布, 而西北部地区成暖干化分布, 年均温自西北向东南逐渐递增, 年均降水量自东向西逐渐递减。
| 类别 Category |
指标 Indicators | 类别 Category |
指标 Indicators |
|
| 植被属性 | 地上生物量(AGB) | 土壤性质 | 土壤有机碳(SOC) | |
| Vegetation attributes | 地下生物量(BGB) | Soil properties | 土壤全氮(TN) | |
| 植被高度(H) | 土壤全磷(TP) | |||
| Shannon多样性指数(D) | 土壤全钾(TK) | |||
| Pielou均匀度指数(E) | 土壤有效氮(AN) | |||
| Margalef丰富度指数(R) | 土壤有效磷(AP) | |||
| 土壤有效钾(AK) | ||||
| 土壤容重(BD) | ||||
| 土壤酸碱度(pH) | ||||
| 土壤含水量(SWC) | ||||
| AGB:地上生物量Above-ground biomass;BGB:地下生物量Below ground biomass;H:植被高度Vegetation height;D:Shannon多样性指数Shannon diversity index;E:Pielou均匀度指数Pielou evenness index;R:Margalef丰富度指数Margalef richness index;SOC:土壤有机碳Soil organic carbon;TN:土壤全氮Soil total nitrogen;TP:土壤全磷Soil total phosphorus;TK:土壤全钾Soil total potassium;AN:土壤有效氮Soil available nitrogen;AP:土壤有效磷Soil available phosphorus;AK:土壤有效钾Soil available potassium;BD:土壤容重Soil bulk density;pH:土壤酸碱度Soil pH;SWC:土壤含水量Soil water content | ||||

|
| 图 1 研究样点年均降水量分布图 Fig. 1 The mean annual precipitation distribution map of study sites |

|
| 图 2 研究样点年均温度分布图 Fig. 2 The mean annual temperature distribution map of study sites |
以高寒退化草地为研究对象, 综合任继周[36]、赵新全等[37]、马玉寿等[38]对高寒草地退化程度划分的方法, 将草地退化程度分为4个梯度, 分别是未退化草地(Non-degraded grassland, ND)、轻度退化草地(Lightly degraded grassland, LD)、中度退化草地(Moderately degraded grassland, MD)和重度退化草地(Heavily degraded grassland, HD)(表 2)。针对少数植被盖度分类不同或没有提及植被盖度的文献, 本研究依据所纳入文献对草地退化梯度的分类标准, 进行相同的分类处理。
| 退化程度 Degradation degree |
植被覆盖度 Vegetation cover/% |
退化程度 Degradation degree |
植被覆盖度 Vegetation cover/% |
|
| 未退化草地(ND) Non-degraded grassland | >90 | 中度退化草地(MD) Moderately degraded grassland | 30—60 | |
| 轻度退化草地(LD) Lightly degraded grassland | 60—90 | 重度退化草地(HD) Heavily degraded grassland | <30 | |
| ND:未退化草地Non-degraded grassland;LD:轻度退化草地Lightly degraded grassland;MD:中度退化草地Moderately degraded grassland;HD:重度退化草地Heavily degraded grassland | ||||
对于论文发表的图片数据, 采用WebPlotDigitizer (Version 4.3)软件进行数据提取, 表格数据直接提取, 并按照试验对照组和退化组的平均值(X)、标准差(SD)和样本量(n)形式进行整合。若文献研究结果为土壤有机质含量的, 按照土壤有机质(g/kg)=土壤有机碳(g/kg)×1.724换算为土壤有机碳含量数据[39]。对于没有标准差的文献, 把平均值的10%作为标准差[40], 公式如下:
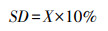
|
(1) |
当文献中出现标准误SE和样本量n时, 根据下式对标准差SD进行转换[41]:
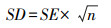
|
(2) |
本研究采用Hedges等[42]在1999年提出的响应比的自然对数RR, 在MetaWin 2.1(Sinauer Associates, Sunderland, USA)软件中, 输入各研究中处理组(退化草地)和对照组(未退化草地)的平均值, 及其对应的标准差和样本量, 得到结果RR和lnR, 计算公式如下:
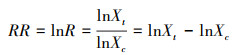
|
(3) |
公式中的RR、R、Xt、Xc和分别为效应值、响应比、处理组的平均值和对照组的平均值。响应比的对数RR的方差v计算公式如下:
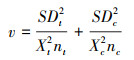
|
(4) |
公式中的nt和nc分别为处理组和对照组的样本量, SDt和SDc分别为处理组和对照组的标准差。
经过非参数权重因子w对所有结果的效应值进行加权, 权重因子w是方差v的倒数:

|
(5) |
lnR++、SDlnR++为非参数权重加权之后的平均效应值和方差, 计算公式如下:
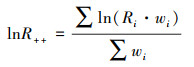
|
(6) |
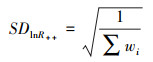
|
(7) |
本文采用随机效应模型来计算效应值95%的置信区间, 提取各指标在不同退化草地下的R++和Bookstrap CI。当权重效应值lnR++的95%CI跨越0时, 则认为草地退化对效应值没有显著影响, 反之有显著影响(P<0.05), 而当95%CI的最大值小于0, 认为草地退化对效应值有显著的负效应, 如果95%CI的最小值大于0, 则认为有显著的正效应。同时, 为了便于直观解释, 通过下面公式把平均效应值转化为百分数C:
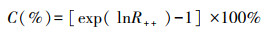
|
(8) |
本文使用MetaWin 2.1对采集的数据进行meta分析, 采用SPSS 26.0对数据进行处理分析, 研究区域概况图使用ArcGIS10.8绘制, 使用SigmaPlot 14.0对结果进行绘图。
2 结果与分析 2.1 土壤理化性质对草地退化的响应从轻度退化草地到中度和重度退化草地的演替过程中, 土壤有机碳、全氮、全磷、有效氮和土壤含水量均显著降低(图 3, P<0.05), 其中土壤有机碳下降幅度最大, 全磷下降幅度最小;土壤容重显著上升, 有效磷、有效钾在中、重度退化阶段显著下降(图 3, P<0.05);全钾、pH变化差异则不显著, pH整体呈现小幅度上升趋势(图 3, P>0.05), 全钾整体呈现小幅度下降趋势(图 3, P>0.05)。与未退化草地相比, 仅土壤容重上升了11.42%;其余理化因子均呈不同程度的下降, 其中, 土壤有机碳含量在重度退化草地降低了54.20%, 全氮含量减少了43.99%, 全磷含量下降了14.90%, 有效氮含量降低了32.51%, 有效磷含量减少了15.07%, 有效钾含量降低了10.28%, 土壤含水量下降了35.79%。

|
| 图 3 草地土壤指标对不同退化程度的响应 Fig. 3 Response of soil feature to grassland degradation SOC:土壤有机碳Soil organic carbon;TN:土壤全氮Soil total nitrogen;TP:土壤全磷Soil total phosphorus;TK:土壤全钾Soil total potassium;AN:土壤有效氮Soil available nitrogen;AP:土壤有效磷Soil available phosphorus;AK:土壤有效钾Soil available potassium;BD:土壤容重Soil bulk density;pH:土壤酸碱度Soil pH;SWC:土壤含水量Soil water content;LD:轻度退化草地Lightly degraded grassland;MD:中度退化草地Moderately degraded grassland;HD:重度退化草地Heavily degraded grassland |
对比不同退化程度的草地, 地上生物量、植被高度表现为一致地显著下降;地下生物量在中、重度退化草地显著降低(图 4, P<0.05)。地上生物量在轻度退化阶段显著降低, 而地下生物量在此阶段下降不显著, 说明地上生物量对于草地退化的响应早于地下生物量, 且在各个退化阶段中地下生物量的效应值低于地上生物量。Shannon多样性指数、Pielou均匀度指数在轻、中度退化草地升高, 在重度退化草地降低;Margalef丰富度指数在轻度退化阶段升高, 在中度和重度退化阶段降低, 其中, 重度退化阶段表现为显著降低(图 4, P<0.05)。在重度退化草地中的地上生物量降低了47.60%;地下生物量降低幅度更大, 下降了68.99%;植被高度在重度退化阶段降低了48.52%;Shannon多样性指数、Pielou均匀度指数、Margalef丰富度指数先升后降, 在重度退化草地分别降低了7.83%、3.84%、19.02%。

|
| 图 4 草地植被指标对不同退化程度的响应 Fig. 4 Response of grassland vegetation indicators to different degree of degradation AGB:地上生物量Above-ground biomass;BGB:地下生物量Below ground biomass;H:植被高度Vegetation height;D:Shannon多样性指数Shannon diversity index;E:Pielou均匀度指数Pielou evenness index;R:Margalef丰富度指数Margalef richness index |
草地退化演替过程中土壤理化性质与植物生物量紧密相关, 对重度退化阶段多个土壤理化性质响应比与植物生物量响应比做线性回归分析。研究结果表明:土壤有机碳、全氮、全磷与地上生物量、地下生物量存在显著的正相关(图 5, P<0.05);有效氮、有效磷、有效钾与地上生物量、地下生物量呈显著的正相关(图 6, P<0.05);土壤含水量与地上生物量、地下生物量存在显著的正相关(图 7, P<0.05);而土壤容重与地上生物量、地下生物量呈显著的负相关(图 7, P<0.05);pH与地上生物量、地下生物量呈负相关(图 7, P>0.05);土壤理化性质与地上生物量的相关系数整体上比与地下生物量的相关系数小;地下生物量与土壤容重、土壤含水量呈极显著相关(图 7, P<0.01), 其中, 地下生物量随着土壤容重的增加而下降, 而随着土壤含水量的增加而上升。

|
| 图 5 土壤有机碳、土壤总氮、土壤总磷响应比与地上生物量、地下生物量响应比的关系 Fig. 5 Relationships between response ratio of soil organic carbon、soil total nitrogen、soil total phosphorus and response ratio of above-ground biomass、below ground biomass |

|
| 图 6 土壤有效氮、土壤有效磷、土壤有效钾响应比与地上生物量、地下生物量响应比的关系 Fig. 6 Relationships between response ratio of soil available nitrogen、soil available phosphorus、soil available potassium and response ratio of above-ground biomass、below ground biomas |

|
| 图 7 土壤容重、土壤酸碱度、土壤含水量响应比与地上生物量、地下生物量响应比的关系 Fig. 7 Relationships between response ratio of soil bulk density、soil pH、soil water content and response ratio of above-ground biomass、below ground biomass |
土壤理化性质的改变与植物群落结构特征密切相关。在本研究中, 退化草地中的土壤含水量和土壤养分在整体上与植被覆盖度和生物量的变化趋势一致, 这与多数学者的研究结果一致[43—44]。随着高寒草地退化程度的加剧, 土壤含水量降低, 而土壤容重、pH增大(图 3)。土壤容重随着草地退化加剧而增大, 这与高寒草地过度放牧有关[45], 由于牲畜对地表植被的过度践踏, 使得土壤紧密度增加, 同时相对松散的土壤细颗粒因风蚀、流水作用容易被吹走和流失[46], 出现大量的砂砾石[47], 导致土壤容重增大。土壤容重的增加使土壤团粒结构丧失、孔隙度减小、渗透阻力增大, 加上草地植被覆盖度降低造成土壤裸露面积增大, 引起地表蒸发量增加和出现少量碳酸钙等盐分聚集到土壤表面, 造成土壤含水量降低和pH值升高[48];此外, 植物生物量降低引起土壤腐殖酸的减少以及太阳辐射对裸土中有机酸类物质的破坏[49], 最终都会使草地趋于盐渍化[50]。
在草地退化演替过程中, 由于持续的全球气候变化以及频繁的人类活动, 使得土壤养分发生改变, 进而影响着草地生态系统的结构与功能。本研究中, 结果表明, 土壤有机碳、全氮和全磷均随退化程度的加剧而显著下降(图 3)。土壤中有机碳、全氮和全磷主要来源于凋落物和植物根系的分解[51]。由于草地退化后植物生物量降低, 植被枯落物也相应减少, 使得地上与地下生产力和土壤有机质的输入不断下降, 同时较高的土壤温度能增强土壤呼吸作用, 促进土壤微生物和根系等分解土壤有机质, 加快土壤有机碳的损失。土壤全氮和全磷含量的减少, 一是由于家畜过度啃食, 舔食茎叶时将富集氮磷的植物根系一同拔出土壤而导致养分流失[51]。二是为了保证地上植物有充足的养分, 土壤微生物活动增强, 加快了土壤中全量养分的分解。在各个退化阶段中全钾的分布均无明显变化(图 3), 这与孙磊等[19]研究的结果一致, 由于土壤中全钾多以矿物态钾的形式存在, 而只有经过矿化后释放出来的游离态钾离子才能被植物所吸收利用, 因此全钾在不同退化阶段的变化不明显。
相关研究表明[52], 土壤的腐殖层随退化程度加大而逐渐变薄, 结构趋向单一, 由于腐殖层含有大量的土壤养分, 所以土壤养分含量因腐殖层变薄而逐渐降低(图 3)。有效氮、有效磷含量的减少, 一方面可能是植被覆盖度减少, 地表裸露面积增大, 更易受到风化和雨水侵蚀的影响, 带走部分土壤养分。另一方面, 随着草地退化程度加剧, 地表植物枯落层变薄, 所截留的降水量减少, 抑制了微生物的活动, 导致矿化作用减弱, 进一步减少对有效氮和有效磷的转化。我们的研究结果与杨军等[49]的发现相似, 有效钾含量在轻、中度退化阶段没有明显变化, 而在重度退化阶段开始下降, 主要原因是轻、中度退化阶段家畜排泄的粪尿中含有大量钾, 钾的输入量与钾的输出量相对平衡, 而重度退化阶段植被和家畜变得更少, 有效钾的回归量随之下降。在高寒草地退化演替过程中, 土壤养分减少会加快植被退化, 在一定程度上阻碍了退化草地的生态恢复进程。由此可见, 土壤养分的变化过程可以作为一种响应机制, 用来判断草地退化的程度。
3.2 草地退化对植被生产力和物种多样性的影响植被退化是草地退化的直观表现。随着退化程度的加剧, 生物量、植被覆盖度和植被高度均呈下降趋势, 这与以往的研究结果一致[51], 三者都是反映高寒草地退化的主要指标[53]。在放牧干扰下, 草地地表被牲畜严重踩踏, 牲畜迅速消耗大量的优良牧草, 植物群落结构发生变化, 植被开始退化[54]。草地退化对生物量的影响有两种不同的结果, 一是地上和地下生物量下降(图 4), 主要是优势种及禾草生物量降低引起的[55], 这是由于在未退化草地中禾草类占据着地上生产力的主要地位, 并产生发达的根系, 但是伴随着退化程度的加剧, 禾草类的生物量减少, 导致群落整体的地上生物量和地下生物量降低, 这以结果印证了仁青吉等[56]、张帆等[57]在青藏高原的研究结果。二是地上和地下生物量随退化程度加剧先增加后减少, 与郝爱华等[46]、周华坤等[58]的研究结果相似, 生物量的这种动态变化符合“中度干扰假说”, 在中度干扰、中海拔和中度退化的草地上, 会影响优势种植物对资源的利用, 促进生长能力较弱的物种吸收与利用资源, 使植物群落的物种趋于多样化, 生物量逐渐增加[59—61];而在过度的干扰下, 植被呈现单一化, 生物量急剧减少[62—63]。除此之外, 草原上前一年被牲畜践踏的凋落物堆积在地表形成覆盖层, 可以起到一定的遮光作用, 进而影响次年植被的生长, 同时牲畜践踏也有利于减缓遮光作用, 给下冠层植物带来更多的光照[64—65]。相关研究表明, 轻度踩踏促进植物的光合作用, 有利于植物的生长;而重度踩踏不仅抑制植物的光合作用, 还不利于植物的根系生长, 最终导致生物量的下降[66—67]。本研究发现, 地下生物量的变化滞后于地上生物量, 且地下生物量对草地退化的响应大于地上生物量, 这可能是因为植被退化与土壤退化存在着时间差。首先植物通常会优先将有限的能量分配到地上部分, 以满足光合作用和生长所需;其次在草地退化过程中, 一旦植被退化, 土壤质量、水分和养分等开始下降, 导致地下生物量降低和进一步影响地下生物的生存和繁殖;最后植物和土壤之间的相互作用也会导致地下生物量的滞后, 健康的植被可以通过根系分泌物和残留物质来促进土壤生物的生长活动, 从而增加地下生物量, 以上这些过程都需要一定的时间来发展和显现, 因此地下生物量的变化会滞后于地上生物量。且土壤退化受凋落物分解、牲畜排泄物回归和土壤微生物活动等多种因素的综合影响, 导致土壤退化存在滞后效应[51]。植被高度逐渐降低(图 4), 主要受到牲畜过度践踏草地、家畜喜食高大禾草和高原鼠兔的猖獗活动等因素的影响[68]。
植物群落的物种多样性一直是草地生态系统的研究重点[69]。本研究中Shannon多样性指数、Pielou均匀度指数和Margalef丰富度指数均呈单峰变化, 整体呈下降趋势, 这与前人在黄河源区的研究结果一致[70], 峰值都出现在轻度退化阶段(图 4), 这种变化趋势符合“中度干扰假说”, 即中等程度的干扰能维持高的生物多样性, 超过或低于这个阈值, 生物多样性都将会降低[62]。物种多样性主要取决于功能群之间的相互竞争[71]、生态位互补效应[49]、动物的偏食性采食策略[62]等因素。具体原因如下:在退化初期阶段, 过度放牧导致草场长期超载, 牲畜适口性好的草本植物(禾本科、莎草科)逐渐减少[70, 72], 降低了群落的高度, 使部分低矮的杂类草植物获得足够的光照, 为杂类草植物的入侵提供良好的机会[73], 群落各物种个体数目分配较为均匀, 群落物种数目处于相对平衡状态, 导致物种多样性、物种均匀度和物种丰富度在此阶段逐渐增加;而在退化末期阶段, 由于原生优势种逐渐退出群落, 对其他物种的抑制作用明显减弱, 而不受牲畜采食的杂类草却占据大量的生态位, 草地植被逐渐呈现出异种单一化的趋势, 群落中物种分配失衡, 植物群落的物种多样性、物种均匀度和物种丰富度严重下降。
3.3 草地植物生物量与土壤因子的相互关系诸多研究表明[19, 74], 土壤养分与植物生物量二者相互作用, 相互影响, 形成一种正反馈。伴随着草地生态系统的不断退化, 土壤肥力持续下降, 植被生产力明显降低, 说明草地退化同时对植物群落特征和土壤特征产生不利的影响。本研究土壤养分与植物群落地上生物量、地下生物量存在显著的正相关(图 5、图 6), 这是由于草地植物的生长发育需要大量的土壤养分, 而土壤养分主要来源于土壤有机质, 有机质又从植物的凋落物和动物的排泄物中获得补给。一是植物生物量减少, 造成土壤养分的回归量下降;二是土壤养分输入量减少, 则不足以提供植物生长所需的营养元素, 导致植物生长缓慢, 发育不良, 植物生物量和植物凋落物因此也变得稀少, 最终陷入反复的恶性循环。在草地退化演替过程中, 土壤养分和植物生物量伴随着草地退化程度的加剧逐渐降低, 表现出同步恶化的趋势, 因此土壤养分的改变与植物生物量的变化紧密相关。同时, 草地植物生物量的积累, 除了土壤养分, 土壤水分也是植物生长的一个重要因素, 土壤含水量在草地退化过程中急剧下降, 与生物量的变化趋势相同, 存在显著的正相关[49]。此外, 土壤容重、pH随着退化程度的加深而逐渐上升, 与草地植物生物量的变化呈现相反的态势(图 7)。土壤容重与植物生物量呈显著的负相关, 与前人的研究结果一致[46], 这是由于家畜的践踏使土壤紧实度增加, 土壤通透性和水源涵养的能力降低, 限制了土壤养分与水分的运转, 直接影响了植物对养分的吸收, 不利于植物根系发育, 而植物根系的减少反过来使土壤变得更加紧实, 导致土壤容重增大[75]。pH与植物生物量的关联性较弱, 与魏晶晶等[76]的研究结果相同, 这可能与pH上升的幅度比植物生物量下降的幅度小有关。研究发现, 土壤氮、磷含量与植物生物量的相关系数较高, 这说明植物在生长发育过程中, 氮、磷元素是植物生长发育所需的限制性元素[29]。同时还发现各项土壤养分与地下生物量的相关系数总体上比与地上生物量的相关系数高, 说明地下生物量对土壤养分的变化响应更为敏感[57]。综上所述, 退化草地的土壤养分、土壤含水量与植物生物量逐渐下降, 变化趋势基本相同, 而土壤容重、pH逐渐上升, 与草地退化的演替规律相符。
4 结论与展望认识草地退化过程中植被与土壤变化特征, 对青藏高原的草地恢复和生态保护有着重要意义。高寒草地退化是一个复杂且多因素综合调控的过程, 在此过程中, 植被与土壤特征对高寒草地的响应具有协同作用, 16个指标随着退化程度加剧整体呈负面效应, 对植被生长发育、土壤养分与结构维持、生物繁殖产生不利影响。随着高寒草地退化程度的加剧, 土壤有机碳、氮、磷、钾、含水量等总体呈下降趋势, 土壤容重、pH总体呈上升趋势。高寒草地退化导致生物量、植被高度、物种多样性等逐渐下降, 地下生物量下降幅度比地上生物量大, 物种多样性指数随着退化演替先增后减, 整体呈下降趋势, 峰值均出现在轻度退化阶段。土壤养分对草地退化的响应程度为土壤有机碳>全氮>全磷>全钾, 氮、磷含量与植物生物量的相关系数较高, 可以考虑修复退化草地时添加N、P元素, 缓解土壤N、P含量流失速度, 防止草地退化态势进一步加剧。本研究以青藏高原高寒草地为研究对象, 选取16个指标, 运用meta分析评估草地退化对土壤与植被特征的影响, 但未对土壤深度和草地类型进行分类以及纳入的指标不够全面。因此, 在今后应补充更多的指标和进行不同土壤深度和草地类型的亚组分析, 以期在更广的维度揭示高寒退化草地植被与土壤特征变化的一般规律, 进一步提高对草地退化机理的认识, 科学制定修复措施, 有利于加快退化草地的恢复重建, 为研究和治理高寒草地退化问题提供科学依据。
| [1] |
Janišová M, Michalcová D, Bacaro G, Ghisla A. Landscape effects on diversity of semi-natural grasslands. Agriculture, Ecosystems & Environment, 2014, 182: 47-58. |
| [2] |
杨汝荣. 我国西部草地退化原因及可持续发展分析. 草业科学, 2002, 19(1): 23-27. |
| [3] |
丁明军, 张镱锂, 刘林山, 王兆锋. 1982—2009年青藏高原草地覆盖度时空变化特征. 自然资源学报, 2010, 25(12): 2114-2122. DOI:10.11849/zrzyxb.2010.12.012 |
| [4] |
刘兴元, 龙瑞军, 尚占环. 青藏高原高寒草地生态系统服务功能的互作机制. 生态学报, 2012, 32(24): 7688-7697. |
| [5] |
姚檀栋, 陈发虎, 崔鹏, 马耀明, 徐柏青, 朱立平, 张凡, 王伟财, 艾丽坤, 杨晓新. 从青藏高原到第三极和泛第三极. 中国科学院院刊, 2017, 32(9): 924-931. |
| [6] |
尹亚丽. 三江源区退化高寒草甸土壤-微生物互作研究[D]. 西宁: 青海大学, 2020.
|
| [7] |
Xu H J, Wang X P, Zhang X X. Impacts of climate change and human activities on the aboveground production in alpine grasslands: a case study of the source region of the Yellow River, China. Arabian Journal of Geosciences, 2017, 10(1): 17. DOI:10.1007/s12517-016-2801-3 |
| [8] |
孙建, 张振超, 董世魁. 青藏高原高寒草地生态系统的适应性管理. 草业科学, 2019, 36(4): 933-938, 915. |
| [9] |
尚占环, 董世魁, 周华坤, 董全民, 龙瑞军. 退化草地生态恢复研究案例综合分析: 年限、效果和方法. 生态学报, 2017, 37(24): 8148-8160. |
| [10] |
Squires V, Hua L M. North-west China's rangelands and peoples: facts, figures, challenges and responses. Squires V, Hua L, Li G, Zhang D. Towards Sustainable Use of Rangelands in North-West China. Dordrecht: Springer, 2010: 3-18.
|
| [11] |
Miehe G, Schleuss P M, Seeber E, Babel W, Biermann T, Braendle M, Chen F H, Coners H, Foken T, Gerken T, Graf H F, Guggenberger G, Hafner S, Holzapfel M, Ingrisch J, Kuzyakov Y, Lai Z P, Lehnert L, Leuschner C, Li X G, Liu J Q, Liu S B, Ma Y M, Miehe S, Mosbrugger V, Noltie H J, Schmidt J, Spielvogel S, Unteregelsbacher S, Wang Y, Willinghöfer S, Xu X L, Yang Y P, Zhang S R, Opgenoorth L, Wesche K. The Kobresia pygmaea ecosystem of the Tibetan highlands-Origin, functioning and degradation of the world's largest pastoral alpine ecosystem. Science of the Total Environment, 2019, 648: 754-771. DOI:10.1016/j.scitotenv.2018.08.164 |
| [12] |
李秋年. 高寒草甸退化草地植物群落结构特征及物种多样性的初步分析. 青海环境, 2004, 14(1): 30-33. |
| [13] |
刘安榕, 杨腾, 徐炜, 上官子健, 王金洲, 刘慧颖, 时玉, 褚海燕, 贺金生. 青藏高原高寒草地地下生物多样性: 进展、问题与展望. 生物多样性, 2018, 26(9): 972-987. |
| [14] |
付建新, 曹广超, 郭文炯. 1980—2018年祁连山南坡土地利用变化及其驱动力. 应用生态学报, 2020, 31(8): 2699-2709. |
| [15] |
吴雪, 刘峰贵, 刘林山, 刘飞, 才项措毛. 青藏高原牲畜数量变化及其空间特征. 生态科学, 2021, 40(6): 38-47. |
| [16] |
朴世龙, 张宪洲, 汪涛, 梁尔源, 汪诗平, 朱军涛, 牛犇. 青藏高原生态系统对气候变化的响应及其反馈. 科学通报, 2019, 64(27): 2842-2855. |
| [17] |
Dong S K, Shang Z H, Gao J X, Boone R B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agriculture, Ecosystems & Environment, 2020, 287: 106684. |
| [18] |
李玲, 张福平, 冯起, 王虎威, 魏永芬, 李肖娟, 聂硕, 刘洁遥. 环青海湖地区草地对气候变化和人类活动的响应. 生态学杂志, 2019, 38(4): 1157-1165. |
| [19] |
孙磊, 刘玉, 武高林, 魏学红. 藏北退化草地群落生物量与土壤养分的关系. 草业科学, 2016, 33(6): 1062-1069. |
| [20] |
高巧静, 朱文琰, 侯将将, 王娅琳, 赵新全, 徐世晓, 孙平. 放牧强度对高寒草甸植物叶片生态化学计量特征的影响. 中国草地学报, 2019, 41(3): 45-50. |
| [21] |
张骞, 马丽, 张中华, 徐文华, 周秉荣, 宋明华, 乔安海, 王芳, 佘延娣, 杨晓渊, 郭婧, 周华坤. 青藏高寒区退化草地生态恢复: 退化现状、恢复措施、效应与展望. 生态学报, 2019, 39(20): 7441-7451. |
| [22] |
赵贯锋, 余成群, 武俊喜, 罗黎鸣, 苗彦军. 青藏高原退化高寒草地的恢复与治理研究进展. 贵州农业科学, 2013, 41(5): 125-129. DOI:10.3969/j.issn.1001-3601.2013.05.036 |
| [23] |
曹广民, 林丽, 张法伟, 李以康, 韩道瑞, 龙瑞军. 青藏高原高寒矮嵩草草甸稳定性的维持、丧失与恢复. 草业科学, 2010, 27(8): 34-38. DOI:10.3969/j.issn.1001-0629.2010.08.007 |
| [24] |
肖玉. 青藏高原高寒草原不同退化程度植物群落特征与土壤养分的关系[D]. 兰州: 兰州大学, 2016.
|
| [25] |
周华坤, 赵新全, 周立, 刘伟, 李英年, 唐艳鸿. 青藏高原高寒草甸的植被退化与土壤退化特征研究. 草业学报, 2005, 14(3): 31-40. |
| [26] |
张建贵, 王理德, 姚拓, 李海云, 高亚敏, 胡彦婷, 兰晓君, 周彤. 东祁连山不同退化草地植物群落特征与土壤养分特性. 水土保持学报, 2019, 33(1): 227-233. |
| [27] |
汪诗平. 青海省"三江源"地区植被退化原因及其保护策略. 草业学报, 2003, 12(6): 1-9. |
| [28] |
任继周, 林慧龙. 江河源区草地生态建设构想. 草业学报, 2005, 14(2): 1-8. |
| [29] |
彭艳, 孙晶远, 马素洁, 王向涛, 魏学红, 孙磊. 藏北不同退化阶段高寒草甸植物群落特征与土壤养分特性. 草业学报, 2022, 31(8): 49-60. |
| [30] |
伍星, 李辉霞, 傅伯杰, 靳甜甜, 刘国华. 三江源地区高寒草地不同退化程度土壤特征研究. 中国草地学报, 2013, 35(3): 77-84. |
| [31] |
Lu X Y, Kelsey K C, Yan Y, Sun J A, Wang X D, Cheng G W, Neff J C. Effects of grazing on ecosystem structure and function of alpine grasslands in Qinghai-Tibetan Plateau: a synthesis. Ecosphere, 2017, 8(1): e01656. DOI:10.1002/ecs2.1656 |
| [32] |
张镱锂, 刘林山, 摆万奇, 沈振西, 阎建忠, 丁明军, 李双成, 郑度. 黄河源地区草地退化空间特征. 地理学报, 2006, 61(1): 3-14. |
| [33] |
邵建翔, 刘育红, 马辉, 魏卫东. 退化高寒草地浅层土壤理化性质Meta分析. 草地学报, 2022, 30(6): 1370-1378. |
| [34] |
闫玉春, 唐海萍, 张新时. 草地退化程度诊断系列问题探讨及研究展望. 中国草地学报, 2007, 29(3): 90-97. |
| [35] |
Tang G. Digital elevation model of China (1KM). National Tibetan Plateau Data Center, 2019.
|
| [36] |
任继周. 草业科学研究方法. 北京: 中国农业出版社, 1998: 95.
|
| [37] |
赵新全. 三江源区退化草地生态系统恢复与可持续管理. 北京: 科学出版社, 2011.
|
| [38] |
马玉寿, 郎百宁, 李青云, 施建军, 董全民. 江河源区高寒草甸退化草地恢复与重建技术研究. 草业科学, 2002, 19(9): 1-5. DOI:10.3969/j.issn.1001-0629.2002.09.001 |
| [39] |
魏卫东, 刘育红, 马辉. 高寒草甸退化草地土壤有机碳含量Meta分析研究. 环境科学与管理, 2018, 43(10): 53-58. |
| [40] |
Luo Y Q, Hui D F, Zhang D Q. Elevated co2stimulates net accumulations of carbon and nitrogen in land ecosystems: a meta-analysis. Ecology, 2006, 87(1): 53-63. DOI:10.1890/04-1724 |
| [41] |
张彦军, 党水纳, 任媛媛, 梁婷, 郁科科, 邹俊亮, 刘方. 基于Meta分析的土壤呼吸对凋落物输入的响应. 生态环境学报, 2020, 29(3): 447-456. |
| [42] |
Hedges L V, Gurevitch J, Curtis P S. The meta-analysis of response ratios in experimental ecology. Ecology, 1999, 80(4): 1150-1156. DOI:10.1890/0012-9658(1999)080[1150:TMAORR]2.0.CO;2 |
| [43] |
李雪萍, 许世洋, 李敏权, 王建喜, 郭致杰, 漆永红, 荆卓琼, 李建军, 谢志军. 甘南州不同退化程度高寒草甸植被及土壤特性的演化规律. 生态学报, 2022, 42(18): 7541-7552. |
| [44] |
李军豪, 杨国靖, 王少平. 青藏高原区退化高寒草甸植被和土壤特征. 应用生态学报, 2020, 31(6): 2109-2118. |
| [45] |
王婷, 花蕊, 楚彬, 周睿, 叶国辉, 牛钰杰, 唐庄生, 花立民. 高寒草原退化对植物群落及土壤理化性质的影响. 草原与草坪, 2019, 39(4): 65-71. |
| [46] |
郝爱华, 薛娴, 彭飞, 尤全刚, 廖杰, 段翰晨, 黄翠华, 董斯扬. 青藏高原典型草地植被退化与土壤退化研究. 生态学报, 2020, 40(3): 964-975. |
| [47] |
杨冲, 王春燕, 王文颖, 毛旭峰, 周华坤, 陈哲, 索南吉, 靳磊, 马华清. 青藏高原黄河源区高寒草地土壤营养特征变化及质量评价. 生态环境学报, 2022, 31(5): 896-908. |
| [48] |
佘延娣, 杨晓渊, 马丽, 张中华, 王党军, 黄小涛, 马真, 姚步青, 周华坤. 退化高寒草甸植物群落和土壤特征及其相互关系研究. 草地学报, 2021, 29(S1): 62-71. |
| [49] |
杨军, 刘秋蓉, 王向涛. 青藏高原高山嵩草高寒草甸不同退化阶段植物群落与土壤养分. 应用生态学报, 2020, 31(12): 4067-4072. |
| [50] |
王玉琴, 尹亚丽, 李世雄. 不同退化程度高寒草甸土壤理化性质及酶活性分析. 生态环境学报, 2019, 28(6): 1108-1116. |
| [51] |
周丽, 张德罡, 贠旭江, 董永平, 王加亭, 赵雅丽. 退化高寒草甸植被与土壤特征. 草业科学, 2016, 33(11): 2196-2201. |
| [52] |
何婷, 邹德强, 陈智华, 文勇立. 石渠县不同沙化草地植被及土壤剖面特征研究. 草学, 2021, No.258(1): 39-44. |
| [53] |
卢虎, 姚拓, 李建宏, 马文彬, 柴晓虹. 高寒地区不同退化草地植被和土壤微生物特性及其相关性研究. 草业学报, 2015, 24(5): 34-43. |
| [54] |
陈乐乐, 施建军, 王彦龙, 马玉寿, 董全民, 侯宪宽. 高寒地区不同退化程度草地群落结构特征研究. 草地学报, 2016, 24(1): 210-213. |
| [55] |
刘玉, 马玉寿, 施建军, 王彦龙, 李世雄, 景美玲, 闵星星. 大通河上游高寒草甸植物群落的退化特征. 草业科学, 2013, 30(7): 1082-1088. |
| [56] |
仁青吉, 武高林, 任国华. 放牧强度对青藏高原东部高寒草甸植物群落特征的影响. 草业学报, 2009, 18(5): 256-261. |
| [57] |
张帆, 李元淳, 王新, 朱剑霄. 青藏高原高寒草甸退化对草地群落生物量及其分配的影响. 草业科学, 2021, 38(8): 1451-1458. |
| [58] |
周华坤, 赵新全, 温军, 陈哲, 姚步青, 杨元武, 徐维新, 段吉闯. 黄河源区高寒草原的植被退化与土壤退化特征. 草业学报, 2012, 21(5): 1-11. |
| [59] |
张倩, 杨晶, 姚宝辉, 蔡志远, 孙小妹, 王缠, 郭怀亮, 谭宇尘, 苏军虎. 放牧管理模式对高寒草甸鼢鼠鼠丘群落演替的影响. 生态学报, 2020, 40(8): 2802-2811. |
| [60] |
温璐, 董世魁, 朱磊, 施建军, 刘德梅, 王彦龙, 马玉寿. 环境因子和干扰强度对高寒草甸植物多样性空间分异的影响. 生态学报, 2011, 31(7): 1844-1854. |
| [61] |
段敏杰, 高清竹, 万运帆, 李玉娥, 郭亚奇, 旦久罗布, 洛桑加措. 放牧对藏北紫花针茅高寒草原植物群落特征的影响. 生态学报, 2010, 30(14): 3892-3900. |
| [62] |
张扬建, 朱军涛, 沈若楠, 王荔. 放牧对草地生态系统影响的研究进展. 植物生态学报, 2020, 44(5): 553-564. |
| [63] |
Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199(4335): 1302-1310. |
| [64] |
Xiao H, Peng Z, Xu C L, Zhang D G, Chai J L, Pan T T, Yu X J. Yak and Tibetan sheep trampling inhibit reproductive and photosynthetic traits of Medicago ruthenica var. inschanica. Environmental Monitoring and Assessment, 2018, 190(9): 1-16. |
| [65] |
Li L, Zhang J, He X Z, Hou F J. Sheep trampling modifies soil and plant C: N: P stoichiometry in a typical steppe of the loess plateau. Rangeland Ecology & Management, 2021, 76: 100-108. |
| [66] |
苗彦军, 付娟娟, 孙永芳, 褚希彤, 益西措姆, 呼天明, 许岳飞. 牦牛放牧模式对西藏高山嵩草草甸群落特征的影响. 草地学报, 2014, 22(5): 935-941. |
| [67] |
许岳飞, 益西措姆, 付娟娟, 陈浩, 苗彦军, 陈俊, 呼天明, 镡建国. 青藏高原高山嵩草草甸植物多样性和土壤养分对放牧的响应机制. 草地学报, 2012, 20(6): 1026-1032. |
| [68] |
杨元武, 李希来, 周旭辉, 祁银姐, 师月英, 李成一, 周华坤. 高寒草甸植物群落退化与土壤环境特征的关系研究. 草地学报, 2016, 24(6): 1211-1217. |
| [69] |
李通, 崔丽珍, 杨雅茜, 宋秀芳, 唐立, 崔骁勇, 郝彦宾, 王艳芬. 黄河流域生态水文学研究态势分析. 中国科学院大学学报, 2023, 40(1): 29-38. |
| [70] |
李成阳, 张文娟, 赖炽敏, 彭飞, 陈小杰, 薛娴, 王涛, 尤全刚, 杜鹤强. 黄河源区不同退化程度高寒草原群落生产力、物种多样性和土壤特性及其关系研究. 生态学报, 2021, 41(11): 4541-4551. |
| [71] |
姜林, 胡骥, 杨振安, 詹伟, 赵川, 朱单, 何奕忻, 陈槐, 彭长辉. 植物功能群去除对高寒草甸群落结构、多样性及生产力的影响. 生态学报, 2021, 41(4): 1402-1411. |
| [72] |
Dai L C, Fu R Y, Guo X W, Du Y G, Lin L, Zhang F W, Li Y K, Cao G M. Long-term grazing exclusion greatly improve carbon and nitrogen store in an alpine meadow on the northern Qinghai-Tibet Plateau. CATENA, 2021, 197: 104955. |
| [73] |
肖玉, 陈米贵, 周杰, 郭正刚. 青藏高原腹地青藏苔草草原不同退化程度的植物群落特征. 应用与环境生物学报, 2014, 20(4): 639-645. |
| [74] |
赵景学, 陈晓鹏, 曲广鹏, 多吉顿珠, 尚占环. 藏北高寒植被地上生物量与土壤环境因子的关系. 中国草地学报, 2011, 33(1): 59-64. |
| [75] |
厉方桢, 钟华平, 欧阳克蕙, 赵小敏, 李愈哲. 基于机器学习的阿勒泰地区草地地下生物量估测与数字制图. 草业学报, 2022, 31(8): 13-23. |
| [76] |
魏晶晶, 秦瑞敏, 张中华, 李永良, 邓艳芳, 马丽, 苏洪烨, 佘延娣, 常涛, 周宸宇, 杨旭芹, 邵新庆, 孙建, 周华坤. 不同退化程度高寒草地植物群落与土壤性质变化及相关性分析. 草地学报, 2022, 30(11): 3035-3045. |