 2024, Vol. 44
2024, Vol. 44文章信息
- 许宏斌, 苏艳龙, 张雷, 刘红梅, 刘丽英, 杨溢文, 李琳
- XU Hongbin, SU Yanlong, ZHANG Lei, LIU Hongmei, LIU Liying, YANG Yiwen, LI Lin
- 围封10年对荒漠草原群落物种多样性与优势种空间分布格局的影响
- Effects of ten years enclosure on species diversity and spatial distribution pattern of dominant species in desert steppe communities
- 生态学报. 2024, 44(10): 4334-4341
- Acta Ecologica Sinica. 2024, 44(10): 4334-4341
- http://dx.doi.org/10.20103/j.stxb.202211303463
-
文章历史
- 收稿日期: 2022-11-30
- 网络出版日期: 2024-03-03






2. 内蒙古大学, 生态与环境学院, 呼和浩特 010021
2. School of Ecology and Environment, Inner Mongolia University, Hohhot 010021, China
草地生态系统是我国面积最大的陆地生态系统, 总面积约4×108 hm2, 占我国陆地面积的41.7%[1]。内蒙古荒漠草原处于草原与荒漠两大陆地生态系统的过渡区, 是重要的放牧畜牧业基地, 也是我国北方重要的生态安全屏障[2]。近年来, 由于气候变暖、过牧、乱垦、开矿、道路建设等因素使得草地发生不同程度的退化, 主要表现为群落覆盖度降低、生物量降低、植被空间格局发生变化, 严重阻碍了区域的可持续发展[3—5]。因此, 采取何种草地管理方式既能维持草地生态系统稳定, 又能兼顾草原产业的可持续发展受到许多专家学者的关注[6—7]。
为遏制草地进一步的退化, 我国实施了围栏封育、轮牧、草蓄平衡等一系列生态修复措施, 使退化草原的情况有所好转[8]。围栏封育作为退化草地修复的重要措施之一, 因其成本低、简便易行而广泛应用于退化草原恢复中[9—10]。通常看来, 围封能够改善退化草地的植物群落结构、提升植被生产力、改善土壤条件, 是草地生态系统有效的恢复方式, 退化草地经围封后, 草地群落生产力可在短时间内恢复, 并且植物群落盖度、平均高度、枯落物积累量、生物量等大幅提高[11—13]。高凤等[14]对藏北高寒草地围封3年后发现, 植被平均高度、盖度和地上生物量均极显著增加, 并且土壤的理化性质有所改善;黄国柱等[15]对祁连山中部亚高山草地进行不同年限围封后发现, 围封2年后生物量达到最大值, 之后生物量随着围封年限的增加逐年降低, 并且围封6年后生物多样性显著降低;左万庆等[16]在锡林郭勒盟退化羊草草原进行围封实验时也发现围封可显著提高群落植被生物量、高度和凋落物量。Miao等[17]在长期围封禁牧对科尔沁沙地植物群落影响中发现, 在长期围封区围栏内群落物种多样性增加;而刘刚等[18]在对围封多年荒漠草原研究时发现, 随着围封年限的增加, 群落内物种多样性逐年降低, Ardavan等[19]也发现围封对荒漠草原进行重度放牧和围栏封育试验时得出围栏封育后物种多样性显著提高, 并在围封20年时达到最大。上述研究表明, 围封可在一定程度上对草地恢复过程起到促进作用, 但围封对植被群落的影响是多重因素的综合作用, 因此围封过程对植物群落的群落演替和优势物种分布格局的影响不同, 进而引起草地的恢复效果也不尽一致。
荒漠草原是草原向荒漠过渡的属草原植被中最干旱的一类草原, 因其年际温差大、干旱、多风和降雨量少且波动较大等自然特征严重影响大多数修复措施的实施, 因而围封成为该地区为数不多可以实施的修复措施。如何进一步合理、高效地实施围封政策也是目前荒漠草原利用和管理中面临的重要问题。本研究以短花针茅(Stipa breviflora)荒漠草原为研究对象, 通过开展长期定位围封试验对荒漠草原植物群落的盖度、密度、生物量、生物多样性及优势种空间分布格局进行研究, 探究围封10年后对荒漠草原群落变化特征、优势物种的生态分布格局以及围封后生态恢复效果的变化, 以期为荒漠草原的可持续发展及科学管理提供理论依据。
1 材料与方法 1.1 研究区概况研究区位于内蒙古自治区锡林郭勒盟苏尼特右旗, 地理位置为E112°43′27″, N42°48′04″, 平均海拔为1098 m, 该地区为中温带大陆性季风气候, 春季多风干旱, 夏季炎热少雨, 年平均降水量为270 mm, 多集中在5——8月, 蒸发量为1910 mm, 年均温为3℃。区域内土壤表层被大量的第三纪中生代红色砂岩、泥沙和沙砾层覆盖, 上层覆有较薄的第四组残积物, 土壤类型为淡栗钙土, 是荒漠草原向荒漠过渡的地带性土壤, 沙化性地表, 腐殖质层厚度约20—30 cm, 腐殖质含量约1.0%—1.8%。植物群落以短花针茅(Stipa breviflora)、无芒隐子草(Cleistogenes songorica)和糙隐子草(Cleistogenes squarrosa)为优势种, 主要伴生种有银灰旋花(Convolvulus ammannii)、木地肤(Kochia prostrata)等。
1.2 调查与取样调查取样时间为2021年8月, 围封样地设置在锡林郭勒盟苏尼特右旗林草局围封实验区, 围栏建立于2011年。自由放牧设置在围栏外, 利用方式为全年自由放牧。两样地仅围栏相隔, 土壤状况、水分等条件均为一致, 样地在围封前均为连续放牧草场。在每个样地拉两条延长线相互垂直的50 m样线, 沿每条样线用1 m×1 m的样方框随机取间隔8 m的样方6个, 调查样方内植物的种类、株数、盖度、密度、高度, 将样方内植物齐地面刈割后分种装入信封内带回实验室, 在65℃烘箱中烘至恒重后称重得到生物量。在每个样地内设置3个2 m×2 m的点格局样方, 以样方的同一方向顶点作为坐标原点, 建立二维坐标系, 测量并记录样方内短花针茅和糙隐子草相对于原点的空间位置(X、Y坐标值)用于分析建群种空间分布格局。
1.3 数据处理与分析植物群落原始数据整理和计算采用Excle 2019进行, 群落数量特征、多样性指数特征采用SPSS 22软件进行单因素方差(One-way ANOVA)分析。点格局数据在Programita 2008软件下完成, 数据采用Monte-Carlo随机模拟的方法将坐标数据进行拟合检验, 计算上、下包迹线, 用随机模型重复拟合各组点的坐标值, 计算100次不同空间尺度r的L(r)值, 得到99%的置信区间。当L(r)=0, 则位于上下包迹线之间, 当L(r)>0, 则位于上包迹线之上, 当L(r)<0, 则位于下包迹线之下。用Origin 2021软件绘图, 横坐标为空间尺度r, 纵坐标为L(r)值。
物种多样性指标计算:计算物种重要值以及Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数等4种多样性指数作为描述四子王旗荒漠草原植物群落综合特征的指标。物种重要值计算方法如下[20]:

|
物种多样性指数计算公式如下[21]:

|
式中, H表示Shanno-Wiener多样性指数, S表示样方内物种总数量, Pi表示第i个物种的个体数Ni占总个体数N的比例, Pi = Ni / N, 其中, N表示样方内全部物种个体总数, D表示Simpson优势度指数;式中, J表示Pielou均匀度指数, R表示Margalef物种丰富度指数。
点格局计算:点格局分析基于群落优势种短花针茅和糙隐子草种群分布的二维坐标, 根据Ripley′s的K(r)函数[22]。
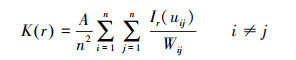
|
式中, K是两点间距离分布的测定指标;r为空间尺度;A为样方面积;n为种群的株数;uij为第i株植物和第j株植物之间的距离, 当uij≤r时, Ir(uij)=1, 当uij>r时, Ir(uij)=0;Wij是以i为圆心、以uij为半径的圆落在面积A中的弧长与整个圆周的比值, 若全部位于样方A中, 则Wij=1;若部分位于样方A中, 0<Wij<1, 即校正边界效应。
在应用中, 常使用Besag修正的L(r)函数, 可使方差更为稳定, 结果更可靠[23]。
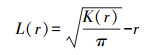
|
当L(r)=0, 则种群在空间尺度r上呈随机分布;当L(r)>0, 则种群在空间尺度r上呈聚集分布;当L(r)<0, 则种群在空间尺度r上呈均匀分布。
2 结果与分析 2.1 荒漠草原群落物种重要值如表 1所示, 苏尼特右旗荒漠草原群落组成较为简单, 研究区内共出现植物23种, 隶属12科19属, 禾本科植物最多为6属7种, 其次为石蒜科为1属3种。围封样地内有10科15属17种, 未围封样地内有12科18属21种, 围封10年后物种所属的科属较未围封样地所属的科属种均有所减少。围封样地各物种重要值在0.0337—0.2170, 其中禾本科植物优势度占群落总优势度的57.4%, 糙隐子草重要值最大为0.2170;未围封样地各物种重要值在0.0270—0.3565, 其中禾本科植物占群落总优势度的84.6%, 短花针茅重要值最大为0.3565。围封样地优势种为糙隐子草、蒺藜(Tribulus terrester)与狭叶锦鸡儿(Caragana stenophylla), 主要伴生种有银灰旋花、短花针茅与鸦葱(Scorzonera austriaca);未围封样地优势种为短花针茅、无芒隐子草, 主要伴生种为狭叶锦鸡儿和草麻黄(Ephedra sinica)。并且除优势种外, 围封下同一物种的优势度较未围封均有提高。未围封样地的植物群落表现为典型的荒漠草原特征, 而围封后群落优势种及主要伴生种发生改变, 可见围封对荒漠草原群落组成有较大影响。
| 生活型 Life Form |
科 Family |
属 Genus |
物种 Species |
重要值Important value | |
| 围封 Enclosure |
未围封 No enclosure |
||||
| 多年生Perennial | 禾本科Poaceae | 隐子草属Cleistogenes | 糙隐子草Cleistogenes squarrosa | 0.2170 | 0.1248 |
| 多年生Perennial | 无芒隐子草Cleistogenes songorica | 0.1815 | |||
| 多年生Perennial | 针茅属Stipa | 短花针茅Stipa breviflora | 0.1341 | 0.3565 | |
| 多年生Perennial | 香茅属Cymbopogon | 芸香草Cymbopogon distans | 0.0812 | 0.0473 | |
| 多年生Perennial | 狗尾草属Setaria | 狗尾草Setaria viridis | 0.0313 | ||
| 一年生Annual | 画眉草属Eragrostis | 画眉草Eragrostis Pilosa | 0.0691 | 0.0416 | |
| 一年生Annual | 稗属Echinochloa | 稗Echinochloa crus-galli | 0.0728 | 0.0633 | |
| 多年生Perennial | 石蒜科Amaryllidaceae | 葱属Allium | 碱韭Allium polyrhizum | 0.0898 | 0.0892 |
| 多年生Perennial | 蒙古韭Allium mongolicum | 0.0568 | 0.0450 | ||
| 多年生Perennial | 细叶韭Allium tenuissimum | 0.0600 | 0.0567 | ||
| 多年生Perennial | 旋花科Convolvulaceae | 旋花属Convolvulus | 刺旋花Convolvulus tragacanthoides | 0.0786 | |
| 多年生Perennial | 银灰旋花Convolvulus ammannii | 0.1832 | 0.1082 | ||
| 小半灌木Small shrubs | 藜科Chenopodiaceae | 地肤属Kochia | 木地肤Kochia prostrata | 0.0760 | 0.0590 |
| 一年生Annual | 苋科Amaranthaceae | 猪毛菜属Kali | 猪毛菜Kali collinum | 0.0337 | 0.0270 |
| 一年生Annual | 菊科Asteraceae | 栉叶蒿属Neopallasia | 栉叶蒿Neopallasia pectinata | 0.0772 | |
| 多年生Perennial | 鸦葱属Scorzonera | 鸦葱Scorzonera austriaca | 0.1390 | ||
| 一年生Annual | 蒺藜科Zygophyllaceae | 蒺藜属Tribulus | 蒺藜Tribulus terrester | 0.1855 | 0.0772 |
| 一年生Annual | 紫草科Boraginaceae | 鹤虱属Lappula | 鹤虱Lappula myosotis | 0.0344 | |
| 草本状灌木Herbaceous shrubs | 麻黄科Ephedraceae | 麻黄属Ephedra | 草麻黄Ephedra sinica | 0.0876 | 0.1245 |
| 多年生Perennial | 莎草科Cyperaceae | 苔草属Carex | 寸草苔Carex duriuscula | 0.1694 | 0.1002 |
| 多年生Perennial | 天门冬科Asparagaceae | 天门冬属Asparagus | 天门冬Asparagus cochinchinensis | 0.0595 | 0.0723 |
| 多年生Perennial | 唇形科Lamiaceae | 兔唇花属Lagochilus | 冬青叶兔唇花Lagochilus ilicifolius | 0.0392 | |
| 矮灌木Bush | 豆科Fabaceae | 锦鸡儿属Caragana | 狭叶锦鸡儿Caragana stenophylla | 0.1853 | 0.1151 |
围封与未围封的群落数量指标如表 2所示, 围封样地的生物量、盖度和高度均极显著高于未围封样地(P<0.01), 围封样地密度显著高于未围封样地密度(P<0.05)。其中围封后盖度变化最大为增加95%, 其余指标生物量、密度和高度在围封后也增加35%—66%, 可见围封可显著提高群落的生物量、高度、盖度与密度。
| 数量特征指标 Quantitative characteristics index |
围封 Enclosure |
未围封 No closure |
显著性 P |
| 生物量Biomass/g | 56.5933±15.2391 | 38.4075±11.8388 | 0.004 |
| 密度Density | 69.3333±23.7959 | 51.2500±18.0201 | 0.048 |
| 盖度Coverage/% | 20.6667±7.0625 | 10.5833±2.4293 | 0.000 |
| 高度Height/cm | 12.1078±2.1902 | 7.2701±1.7235 | 0.000 |
植物群落各多样性指标变化如表 3所示, 围封10年后Margalef丰富度指数、Shannon-wiener多样性指数、Simpson优势度指数和Pielou均匀度指数均低于未围封样地, 但无显著差异。
| 物种多样性指标 Species diversity index |
围封 Enclosure |
未围封 No enclosure |
显著性 P |
| 丰富度指数Margalef | 1.5560±0.3776 | 1.8897±0.5239 | 0.087 |
| 多样性指数Shannon-Wiener | 1.5082±0.2450 | 1.6635±0.2528 | 0.141 |
| 优势度指数Simpson | 0.7033±0.1090 | 0.7461±0.0662 | 0.258 |
| 均匀度指数Pielou | 0.7581±0.1144 | 0.7999±0.075 | 0.298 |
如图 1所示, 围封样地内共有针茅32株、糙隐子草23株;未围封样地内共有针茅41株、糙隐子草17株。围封样地内糙隐子草在0—1 m尺度内呈随机分布, L(r)值在大于0.6 m尺度上呈现明显的上升状态, 并且L(r)值趋于上包迹线。短花针茅在0—0.63 m呈随机分布, 在0.67—0.84 m呈均匀分布, 在0.84—1.0 m呈随机分布, L(r)值在大于0.64 m空间尺度上有趋于下包迹线的趋势。未围封样地内糙隐子草在0—0.39 m、0.61—0.65 m和0.77—0.81 m和0.95—1.0 m呈随机分布, 在大于0.40 m空间尺度上趋近于随机分布;在其余空间尺度呈均匀分布, 未围封糙隐子草L(r)值在大于0.4 m后与下包迹线发生多处重叠, 趋于随机分布与均匀分布的临界点, 表现为在实际样地调查中呈现均匀分布。未围封短花针茅种群在0—0.05 m呈均匀分布, 在其余0.05—1.0 m呈随机分布, 并且在大于0.8 m的空间尺度上L(r)值趋近于上包迹线, 有表现为聚集分布的趋势。

|
| 图 1 荒漠草原优势种空间分布格局 Fig. 1 Spatial distribution pattern of dominant species in desert steppe |
围封是草地恢复的重要手段之一, 围封对退化草地生态系统的改变主要体现在群落结构中。群落的种类组成和优势种是草原植物群落结构的重要特征, 优势种在群落结构和群落环境的控制上发挥重要作用, 优势种的更替也是群落演替不同阶段的标志[16, 24]。在本研究中, 未围封样地中其优势种为中度退化荒漠草原的指示植物无芒隐子草和短花针茅[8, 25], 在围封10年后, 禾本科植物在群落中优势度较未围封下降明显, 该结果与廖晗茹等[26]在蒙古国的荒漠草原中进行围封后得出禾本科的重要值降低的结果一致[26];张攀等[27]在天山北段温性草原围封中同样得出禾本科植物在群落中所占比例减低的结论。围封后群落优势种发生明显替代变为更为低矮的糙隐子草和蒺藜, 主要伴生种为银灰旋花, 表明围封10年样地与未围封样地在群落结构上产生显著差异。由于长期的围封, 缺乏家畜对土壤的扰动, 枯落物大量堆积和生物结皮大量形成, 枯落物在土壤表面形成阻隔层延长降水入渗的时间, 同时枯落物也会吸收部分降水, 使接触到土壤的有效降水量减少;生物结皮与植物对水肥资源进行竞争, 抢夺生活空间, 生物结皮的存在使水分向下渗透困难, 导致水分入渗困难, 一年生浅根系植物大量生长[28]。乔荠瑢等[29]在荒漠草原上研究发现, 放牧会促进短花针茅等禾本科植物的分蘖, 与本研究中未围封样地中禾本科植物在群落中的优势度较围封后有显著的提高的结论相同。在围封10年后, 狭叶锦鸡儿在群落中成为优势种, 围封后草地灌丛化明显, 该结论与周华坤等人[30]与董轲等人[31]的研究结果相同, 并且灌木根系也能将深层水和养分提升至表层, 供给草本利用[32], 进一步促进浅根系草本植物生长。证明围封并不一定会促进退化草地的恢复, 也可能灌丛化草地是该地区的一种稳态。
3.2 围封对荒漠草原群落指标的影响放牧主要通过家畜的采食、践踏和排泄物直接或间接地影响草原生态系统, 围封后消除了家畜对草地群落的影响, 大量研究表明, 在围封后生物量及盖度、密度、高度等会显著提高[33]。在本研究中与未围封样地相比, 围封后生物量、盖度、密度及高度较未围封有显著提高, 与前人的研究结果相同。但是, 在本研究中围封样地中有大量低矮但盖度及地上生物量较大的蒺藜和银灰旋花等草地退化指示生物, 致使围封后盖度、密度、生物量与未围封有显著差异的一大原因;同时围封会限制植物补偿性生长机制的发挥, 枯落物大量积累, 影响资源的利用效率[34]。此外, 未围封会使植物叶片受到家畜采食与践踏, 植物生长较为低矮, 使植物的高度、地上生物量表现出显著的差异。而围封对物种多样性的影响未得出一致性的结论[30, 35—38], 在本研究中围封10年后生物多样性指数略低于未围封草地但无显著差异, 这与左万庆等[16]在锡林郭勒盟退化羊草草原围封后发现围封后草原多样性指标与未围封样地相比无显著差异的结论相同。
3.3 优势种空间分布格局种群的空间格局是种群的基本特征, 也是研究种群间相互作用以及种群与环境间关系的重要手段[2, 22, 39—41]。分布格局的确定除了受种群本身的生物学特性、种间关系和环境条件决定外, 还与空间格局研究范围的大小密切相关[22, 42—43]。在本研究中, 与未围封相比, 围封降低了短花针茅的密度, 该结论与张峰等以短花针茅为建群种的荒漠草原所得出的结论一致, 均为围封后的短花针茅种群数量显著降低[44]。其原因可能为短花针茅为丛生型禾草, 在围封后消除了家畜的踩踏与采食, 短花针茅多以较大的株丛存在, 而在围栏外由于家畜的采食与践踏, 使大株丛比例减少而分散为较多的小株丛脱离母株而使种群数量增大有关。而围栏外的自由放牧对糙隐子草的影响已有较多研究, 如吴思雨等[45]在不同放牧强度对荒漠草原的糙隐子草功能性状研究中发现糙隐子草会对放牧表现出矮小-密植化的适应策略, 并且王鑫厅等[46]对不同恢复阶段糙隐子草空间分布格局研究时发现糙隐子草会在放牧压力作用下进行超补偿性生长从而影响糙隐子草个体的数量、生长、发育、形态等特征。在本文中, 围栏内糙隐子草数量较多其原因可能为围封消除了家畜的采食与践踏, 而围栏外由于家畜的采食与践踏作用再加上降水的稀少, 糙隐子草无法进行超补偿生长, 致使围栏外糙隐子草数量减少。围封后糙隐子草在全尺度上呈现随机分布, 而围栏外的糙隐子草在小尺度呈现出随机分布, 在大尺度呈现出趋近于均匀分布的分布特征, 糙隐子草在围栏内外分布格局差异显著, 其原因可能为围栏外的糙隐子草作为丛生型禾草, 在被家畜采食及践踏后, 由较大株丛分散为较多小植株聚集生长, 然而由于自然资源的匮乏, 小植株之间由于拥挤及对资源竞争发生自疏现象[47], 致使围栏外糙隐子草在围栏外表现在小尺度为随机分布而在大尺度为均匀分布的分布格局, 而围栏内糙隐子草由于免受家畜的采食与践踏, 多为较大株丛, 对资源的竞争能力较强而表现出随机分布的分布格局。围封的短花针茅在小尺度表现为随机分布且趋于聚集分布, 在0.2—0.6 m范围内L(r)值发生降低直至低于下包迹线, 其原因可能为, 围封后短花针茅株丛较大, 且株丛大小处于0.2—0.6 m的株丛占大多数, 就会表现出在小尺度为随机分布趋于聚集分布, 随着研究尺度的增大, 逐渐表现为均匀分布, 在变为随机分布的趋势, 而未围封的短花针茅均表现为随机分布, 其原因为短花针茅为丛生型禾草, 被家畜采食和践踏后大株丛分散成小株丛, 而后又因为种内竞争发生自疏现象, 导致呈现随机分布的分布格局。
4 结论通过对苏尼特右旗荒漠草原围封10年后与未围封的荒漠草原研究后发现:围封10年与未围封样地相比, 围封10年显著提高了荒漠草原的植被盖度、植被密度、植被高度和地上生物量, 但对植物群落多样性均无显著影响。围封10年对优势种短花针茅和糙隐子草的分布格局造成较大影响, 围封后糙隐子草的分布格局出现单一化趋势, 呈现出由主要随机分布变为均匀分布占主要分布格局的现象, 而短花针茅分布格局呈多样化, 主要表现为由单一的随机分布变为随机分布和均匀分布共存的分布格局。围封是目前最为方便有效的退化草原治理措施之一, 在退化草原植被恢复中得到了广泛的应用, 在选择恢复措施时应根据区域内的退化情况和植物的生长环境综合考虑围封的方式和年限, 促进退化草地的快速恢复和合理利用, 实现草地资源的可持续发展。
| [1] |
张璞进, 黄建辉, 木兰, 单玉梅, 晔薷罕, 温超, 常虹, 任婷婷, 陈世苹, 白永飞. 氮水添加对放牧背景下荒漠草原生产力的影响. 生态学报, 2022, 42(13): 5458-5470. |
| [2] |
黄琛, 张宇, 王静, 李元恒, 五十六, 塔娜, 赵萌莉, 韩国栋, 朗巴达拉呼, 赵艳芳. 不同放牧强度下短花针茅荒漠草原植被的空间异质性. 植物生态学报, 2014, 38(11): 1184-1193. |
| [3] |
高英志, 韩兴国, 汪诗平. 放牧对草原土壤的影响. 生态学报, 2004, 24(4): 790-797. DOI:10.3321/j.issn:1000-0933.2004.04.021 |
| [4] |
吴艳玲, 陈立波, 卫智军, 刘红梅, 运向军, 王颖杰, 展春芳. 不同放牧压短花针茅荒漠草原群落植物种的空间异质特征. 干旱区资源与环境, 2012, 26(7): 110-115. |
| [5] |
沈海花, 朱言坤, 赵霞, 耿晓庆, 高树琴, 方精云. 中国草地资源的现状分析. 科学通报, 2016, 61(2): 139-154. |
| [6] |
李强. 不同恢复措施对松嫩平原退化草地的作用[D]. 长春: 东北师范大学, 2010.
|
| [7] |
尚占环, 董世魁, 周华坤, 董全民, 龙瑞军. 退化草地生态恢复研究案例综合分析: 年限、效果和方法. 生态学报, 2017, 37(24): 8148-8160. |
| [8] |
李博. 中国北方草地退化及其防治对策. 中国农业科学, 1997, 30(6): 2-10. |
| [9] |
肖金玉, 蒲小鹏, 徐长林. 禁牧对退化草地恢复的作用. 草业科学, 2015, 32(1): 138-145. |
| [10] |
侯东杰, 陆帅志, 王静, 郭柯. 典型草原围封后羊草地上功能性状对枯落物累积的响应. 生态学报, 2020, 40(18): 6522-6531. |
| [11] |
Su Y Z, Li Y L, Cui J Y, Zhao W Z. Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, Northern China. Catena, 2005, 59(3): 267-278. DOI:10.1016/j.catena.2004.09.001 |
| [12] |
Descheemaeker K, Muys B, Nyssen J, Poesen J, Raes D, Haile M, Deckers J. Litter production and organic matter accumulation in exclosures of the Tigray Highlands, Ethiopia. Forest Ecology and Management, 2006, 233(1): 21-35. DOI:10.1016/j.foreco.2006.05.061 |
| [13] |
Zhu G Y, Deng L, Zhang X B, Shangguan Z P. Effects of grazing exclusion on plant community and soil physicochemical properties in a desert steppe on the Loess Plateau, China. Ecological Engineering, 2016, 90: 372-381. DOI:10.1016/j.ecoleng.2016.02.001 |
| [14] |
高凤, 王斌, 石玉祥, 张更新, 王建, 斯贵才, 韩丛海, 袁艳丽, 胡盎. 藏北古露高寒草地生态系统对短期围封的响应. 生态学报, 2017, 37(13): 4366-4374. |
| [15] |
黄国柱, 席亚丽, 赵传燕, 刘瑞雪, 杨建红, 李娜, 李伟斌. 围封对祁连山亚高山草地群落结构与生物量的影响. 兰州大学学报: 自然科学版, 2020, 56(6): 718-723. |
| [16] |
左万庆, 王玉辉, 王风玉, 师广旭. 围栏封育措施对退化羊草草原植物群落特征影响研究. 草业学报, 2009, 18(3): 12-19. |
| [17] |
Miao R H, Liu Y Z, Wu L Q, Wang D, Liu Y C, Miao Y, Yang Z L, Guo M X, Ma J. Effects of long-term grazing exclusion on plant and soil properties vary with position in dune systems in the Horqin Sandy Land. Catena, 2022, 209: 105860. |
| [18] |
刘刚, 张克斌, 李瑞, 乔锋, 杨俊杰. 人工封育草场管理研究--以宁夏盐池县为例. 水土保持研究, 2007, 14(2): 252-254. |
| [19] |
Ghorbani A, Dadjou F, Moameri M, Fekri A, Andalibi L, Biswas A, Mohammadi Moghadam S, Sharifi J. Effect of grazing exclusion on soil and vegetation characteristics in desert steppe rangelands: a case study from north-western Iran. Arid Land Research and Management, 2021, 35(2): 213-229. |
| [20] |
宝音陶格涛. 不同改良措施下退化羊草(Leymus chinensis)草原群落恢复演替规律研究[D]. 呼和浩特: 内蒙古大学, 2009.
|
| [21] |
郭忠玲, 马元丹, 郑金萍, 刘万德, 金哲峰. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究. 应用生态学报, 2004, 15(11): 2013-2018. |
| [22] |
张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344-349. |
| [23] |
Baseg J. Contribution to the discussion of Dr.Ripley's paper. Journal of the Royal Statistical Society, 1977, 39: 193-195. |
| [24] |
闫玉春, 唐海萍. 围栏禁牧对内蒙古典型草原群落特征的影响. 西北植物学报, 2007, 27(6): 1225-1232. |
| [25] |
刘钟龄, 王炜, 梁存柱, 郝敦元. 内蒙古草原植被在持续牧压下退化演替的模式与诊断. 草地学报, 1998, 6(4): 244-251. |
| [26] |
廖晗茹, Indree Tuvshintogtokh, 郭通, 赵景学. 围封对蒙古荒漠草原和高山草原植物群落组成及稳定性的影响. 北京大学学报: 自然科学版, 2020, 56(3): 471-478. |
| [27] |
张攀, 马婧婧, 程军回, 李锐, 邵明轩, 盛建东, 刘耘华. 围封对天山北坡中段温性草原植被特征和土壤理化性质的影响. 中国草地学报, 2021, 43(5): 41-50. |
| [28] |
卢晓杰, 李瑞, 张克斌. 农牧交错带地表覆盖物对土壤入渗的影响. 水土保持通报, 2008, 28(1): 1-5. |
| [29] |
乔荠瑢, 杨阳, 张峰, 贾丽欣, 陈大岭, 赵萌莉. 不同载畜率下短花针茅分蘖特征和贮藏养分的变化. 中国草地学报, 2019, 41(5): 80-85. |
| [30] |
周华坤, 周立, 赵新全, 周彦艳, 刘伟, 严作良. 围栏封育对轻牧与重牧金露梅灌丛的影响. 草地学报, 2004, 12(2): 140-144. |
| [31] |
董轲, 丁新峰, 郝广, 王金龙, 赵念席, 高玉葆. 围封年限对内蒙古灌丛化草原小叶锦鸡儿灌丛结构及群落种间关联的影响. 生态学报, 2021, 41(14): 5775-5781. |
| [32] |
王凌菲, 徐霞, 江红蕾, 关梦茜, 田晓宇. 内蒙古温带典型草原围封十年草灌景观格局动态. 生态学报, 2020, 40(7): 2234-2241. |
| [33] |
赵京东, 乌云娜, 宋彦涛. 短期围封对辽西北退化草地群落牧草品质的影响. 草业学报, 2021, 30(9): 51-61. |
| [34] |
Huenneke L F, Mooney H A. Grassland structure and function: California annual grassland. Dordrecht: Kluwer Academic, 1989.
|
| [35] |
王顺霞. 放牧方式对围栏草地植被和土壤环境质量影响的研究[D]. 杨凌: 西北农林科技大学, 2012.
|
| [36] |
张玉萍, 宋乃平, 王兴, 随金明. 不同放牧制度对荒漠草原植物多样性及土壤理化性状的影响. 北方园艺, 2018(19): 109-115. |
| [37] |
朱新萍, 贾宏涛, 蒋平安, 赵成义, 胡玉昆, 曹叶飞, 苏建红. 长期围栏封育对中天山草地植物群落特征及多样性的影响. 草业科学, 2012, 29(6): 989-992. |
| [38] |
Meissner R A, Facelli J M. Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrublands of South Australia. Journal of Arid Environments, 1999, 42(2): 117-128. |
| [39] |
陈宝瑞, 杨桂霞, 张宏斌, 段庆伟, 辛晓平. 不同干扰类型下羊草种群的空间格局. 生态学报, 2010, 30(21): 5868-5874. |
| [40] |
张峰, 郑佳华, 赵天启, 乔荠, 赵萌莉, 张心玥, 吴建新. 放牧强度对荒漠草原一年生植物刺穗藜种群特征及空间分布的影响. 草地学报, 2021, 29(7): 1507-1512. |
| [41] |
白小军, 贾琳, 谷会岩. 大兴安岭次生林区优势种落叶松分布格局及竞争作用. 生态学报, 2021, 41(10): 4194-4202. |
| [42] |
赵峰侠, 尹林克. 荒漠内陆河岸胡杨和多枝柽柳幼苗种群空间分布格局及种间关联性. 生态学杂志, 2007, 26(7): 972-977. |
| [43] |
李博. 生态学. 北京: 高等教育出版社, 2000.
|
| [44] |
张峰, 孙嘉伟, 孙宇, 郑佳华, 乔荠瑢, 赵天启, 陈大岭, 赵萌莉. 不同尺度下荒漠草原建群种短花针茅的空间分布对载畜率的响应. 应用生态学报, 2021, 32(5): 1735-1743. |
| [45] |
吴思雨, 宝音陶格涛, 许宏斌, 张璐. 放牧强度对内蒙古典型草原糙隐子草功能性状的影响. 应用生态学报, 2021, 32(2): 392-398. |
| [46] |
王鑫厅, 王炜, 梁存柱, 包俊江. 不同恢复演替阶段糙隐子草种群的点格局分析. 应用生态学报, 2013, 24(7): 1793-1800. |
| [47] |
张景慧, 王铮, 黄永梅, 陈慧颖, 李智勇, 梁存柱. 草地利用方式对温性典型草原优势种植物功能性状的影响. 植物生态学报, 2021, 45(8): 818-833. |