 2023, Vol. 43
2023, Vol. 43文章信息
- 王成, 周诗薇, 刘红玉, 李玉凤
- WANG Cheng, ZHOU Shiwei, LIU Hongyu, LI Yufeng
- 多类群水鸟共存栖息地网络结构稳定性及其优化方案——以盐城滨海湿地为例
- Robustness and its optimization of the habitat network structure for the multi guilds coexistence: a case study of Yancheng coastal wetlands
- 生态学报. 2023, 43(22): 9193-9205
- Acta Ecologica Sinica. 2023, 43(22): 9193-9205
- http://dx.doi.org/10.20103/j.stxb.202211153293
-
文章历史
- 收稿日期: 2022-11-15
- 采用日期: 2023-03-15





2. 安徽农业大学 经济管理学院, 合肥 230036
2. School of Economics and Management, Anhui Agricultural University, Hefei 230036, China
滨海湿地作为全球迁徙水鸟最重要的栖息地, 也是水鸟物种多样性最为丰富的区域之一, 其在维护全球水鸟种群安全方面具有重要的生态价值[1—2]。然而, 自改革开放以来, 我国经济快速发展, 外向型经济模式加快了沿海地带的开发进程, 致使滨海湿地被大范围侵占。研究资料显示, 中国在过去50年损失了53%的温带滨海湿地[3], 大范围围垦开发正加速滨海湿地水鸟栖息地的空间隔离化和破碎化, 水鸟栖息地保护形势十分严峻。
近年来, 随着滨海湿地在水鸟多样性保护中作用逐渐显现, 很多研究将目光聚焦到了滨海湿地[4—8]。一些研究聚焦于围垦开发活动对滨海湿地内部生态系统的稳定产生的负面作用, 一定程度上揭示了水鸟栖息地丧失的主要外在因素[6, 8]。还有一些研究是从某一群落出发, 运用生态位模型, 揭示水鸟在栖息地利用的时空变化与环境变量的关联性等[4, 7]。随着研究的深入, 一些学者开展了滨海湿地斑块连通性、物种适宜生境空间连接性变化等研究, 反映水鸟在区域生境利用的景观过程[9—10]。然而, 水鸟在重要生境“源地”之间的活动在景观上是非连贯的, 且通过踏脚石斑块作为跳板, 由一块“源地”向其他“源地”扩散[1, 9]。而这些栖息地结构性要素反映了水鸟在滨海湿地内部栖息、觅食和繁殖的动态过程, 因此, 栖息地网络结构性分析有助于揭示水鸟对网络结构要素利用的差异, 并反映现状栖息地结构存在的薄弱环节, 为后续栖息地结构的优化和恢复提供了数据支撑。
盐城滨海湿地是雁鸭类在我国的重要越冬地, 也是鸻鹬类在北半球迁徙的重要中转站, 每年吸引了数十万只各类水鸟在此越冬、栖息和中转[11—12]。其中, 雁鸭类、鸻鹬类和鹭类是数量占比最高的三大类群, 近年来数量占比超过70%[13]。总结现有研究发现, 很多研究聚焦于丹顶鹤等单一物种在盐城滨海湿地栖息地间连通性[5], 却较少关注滨海湿地在支撑多类群水鸟内部迁移方面的重要价值。因此, 在三类水鸟的适宜生境分布研究基础上[7], 开展三类水鸟在盐城滨海湿地的栖息地网络结构现状研究, 能够有效揭示盐城滨海湿地对水鸟多样性支撑方面的不足和空缺。近年来, 基于交叉学科知识的复杂网络理论被广泛运用到网络结构仿真模拟中[14—15], 因其具有高度的适应性、良好的兼容性和模拟结果的直观性近年来逐渐被应用到生态网络构建与评估、生态空间优化等景观生态学领域[15—16]。然而, 复杂网络在水鸟栖息地网络中的应用尚不多见。因此, 本研究基于滨海湿地水鸟多样性维持视角, 利用复杂网络模型模拟栖息地网络受到模拟攻击后网络结构稳定性和连通性的变化, 将进一步拓展该方法的应用领域, 也为滨海湿地水鸟栖息地保护与水鸟多样性水平的提升提供技术和方案参考。
基于此, 本研究将结合盐城滨海湿地主要类群水鸟适宜生境评价数据, 运用复杂网络理论, 开展多类群水鸟栖息地网络结构现状分析, 试图解决一下主要科学问题:(1)多类群水鸟栖息地网络结构现状如何;(2)不同情境下栖息地网络鲁棒性有何差异;(3)如何优化现有水鸟栖息地网络?上述科学问题的解决将有助于揭示盐城滨海湿地多类群水鸟栖息地利用现状, 破解制约盐城滨海湿地水鸟多样性水平提升和水鸟栖息地功能维持的主要网络缺陷, 为后续盐城滨海湿地水鸟栖息地的生态恢复实践提供重要的数据支撑。
1 材料与方法 1.1 研究区选择本研究北起射阳河南至盐城市与南通市行政边界, 总面积为2993.28 km2, 涵盖了北缓、核心、南缓、大丰和条子泥栖息地等五个主要水鸟栖息地(图 1)。该区域是鸻鹬类、雁鸭类和鹭类等主要水鸟类群分布区。研究区是典型的淤泥质海岸带, 围垦历史悠久[6, 8], 并对该区域的水鸟种群和栖息生境产生了深远影响, 使得原本带状连片的栖息地被分割为若干个空间隔离的栖息地, 水鸟在研究区内活动空间限制, 越冬和繁殖种群数量持续下降[7, 11, 17]。此外, 持续的人类干扰也对研究区内部水鸟栖息地网络的稳定性和连通性产生了一定程度破坏, 亟需对现状水鸟栖息地网络开展研究和保护工作。

|
| 图 1 研究区位置图 Fig. 1 Location map of the study area |
本研究涉及的数据源主要有生境斑块、踏脚石斑块数据和水鸟分布数据。其中, 生境和踏脚石斑块数据主要来源于2020年4月高分二号高分辨率遥感影像(空间分辨率2m), 利用面向对象方法对研究区水鸟主要栖息地的生境和踏脚石斑块进行解译。水鸟分布数据主要涉及雁鸭类、鸻鹬类和鹭类等三类水鸟, 其来源于研究团队2018年4月、7月、12月、2019年4月、7月、12月和2020年4月等七次野外调查获得, 调查采用样点法和样线法, 选择天气晴朗、风力较小(3级以下)条件进行, 按照2—3人一组于清晨7点至傍晚5点沿固定样线调查记录水鸟集群数量和坐标信息。三年七次野外调查共记录雁鸭类水鸟26种105934只, 鸻鹬类水鸟45种668057只, 鹭类水鸟12种116026只, 三类水鸟在各栖息地内数量如表 1所示。
| 水鸟类群 Waterbird guilds |
年份 Year |
北缓栖息地 Beihuan habitat site |
核心栖息地 Hexin habitat site |
南缓栖息地 Nanhuan habitat site |
大丰栖息地 Dafeng habitat site |
条子泥栖息地 Tiaozini habitat site |
| 雁鸭类Anatidae | 2018 | 5760 | 27786 | 832 | 7799 | 12848 |
| 2019 | 6264 | 14400 | 8195 | 482 | 20936 | |
| 2020 | 18 | 176 | 29 | 0 | 409 | |
| 鸻鹬类Shorebirds | 2018 | 4554 | 27454 | 10411 | 12875 | 152978 |
| 2019 | 4676 | 29080 | 29366 | 3865 | 359281 | |
| 2020 | 458 | 75 | 2253 | 622 | 30109 | |
| 鹭类Herons | 2018 | 3330 | 50147 | 7565 | 1941 | 2228 |
| 2019 | 2893 | 31714 | 7902 | 743 | 2029 | |
| 2020 | 80 | 5009 | 176 | 12 | 257 |
重要生境节点是水鸟日常觅食、栖息和繁殖等行为的主要场所, 因此识别水鸟的重要生境节点, 是栖息地网络结构分析的重要基础。本研究将主要水鸟类群生境适宜性评价结果中次适宜和最适宜生境区定义为水鸟重要生境区[7], 即适宜性指数值大于0.6的区域。同时, 将适宜性指数在0.5—0.6的次适宜区域定义为扩张生境, 将低适宜区域定义为生境缓冲地带。
除了重要生境节点, 栖息地网络中生态阻力值关系到物种在生境节点之间迁移和水平阻力情况, 本研究依据随机森林模拟水鸟适宜生境分布概率值, 将阻力值设置为概率值的倒数, 得到研究区0.01—1.00之间的生态阻力值。
1.3.2 基于电路理论的潜在生态廊道识别生态廊道是区域内具有一定宽度的景观带, 是维护区域生态过程的主要通道, 也是水鸟在区域内迁移、觅食和栖息的主要通道, 具有重要的生态价值[18]。然而, 水鸟在区域内活动是随机自由的, 无法准确测算物种在生境间如何移动。而在电路理论中, 电荷具有随机游走的特性[19], 与水鸟随机或半随机运动具有高度一致性。因此, 本研究采用电路理论和图论来确定潜在生态廊道。
具体来说, 将景观看作一个导电表面, 借助于图论和电路理论, 将异质性景观抽象为一个由一系列节点和电阻组成的电路, 节点代表重要生境区或源地[20]。根据物理学中的欧姆定律, 在同一电路中, 导体中的电流与导体两端的电压呈正比, 与导体的电阻呈反比。因此两个节点间的电流可用以下公式表示:

|
(1) |
式中, I为节点间的电流, V为电压, 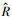
| 名称Term | 单位Unit | 生态学意义Ecological significance |
| 电阻Resistance, R | Ω | 电阻越大, 对物种运动行为阻碍能力越强 |
| 有效电阻Effective resistance, |
Ω | 两个节点间隔离程度的测定指标, 随着节点间连接路径的增加, 有效电阻随之减小 |
| 电流Electric current, I | A | 反映水鸟到达目前生境之前通过相应节点或路径的净次数, 用来预测物种通过相应节点或者路径的净迁移概率, 预测出具有较高通过水平的区域 |
| 电压Voltage, V | V | 物种离开任一节点成功到达给定目标节点的概率, 即成功扩散的概率 |
具体步骤为:首先, 根据图论理论计算景观阻力阈值;其次, 运用电路模型来模拟特定景观阻力阈值下物种的迁移, 预测物种迁移通道范围阈值;最后, 根据景观阻力阈值、物种迁移范围阈值、最小阻力值(0.01)和适宜生境分布确定潜在的生态廊道。在此需要解释的是, 最小阻力值反映了景观要素间的连通水平, 而电流及电阻值反映了物种在区域内的随机扩散属性, 因此景观廊道与物种扩散通道的模拟能够最大程度反映研究区水鸟栖息地潜在生态廊道的真实状态。生态廊道的识别在Linkage Mapper软件和Circuitscape软件(http://www.circuitscape.org/linkagemapper)中进行。
1.3.3 踏脚石斑块的确定与分析踏脚石是位于重要生境区或源地之间一系列小的生境斑块或生境岛屿。在栖息地生境丧失和破碎化条件下, 踏脚石成为不同生境区之间的重要联系, 是物种在区域内迁移的重要中转站和停歇点[21]。目前踏脚石识别存在四大原理[22], 即踏脚石连接度原理、踏脚石间距原理、踏脚石消失原理和踏脚石群原理, 需要根据目标物种的生态习性、行为和当地现状来确定踏脚石生境。本研究根据廊道内高的电流值、潜在廊道区和低阻力值区的叠加来确定三类水鸟踏脚石斑块[20]。本研究将潜在生态廊道中每一个踏脚石视为一个一般节点, 而每个重要生境区或源地视为重要节点。利用Build Network and Map Linkages工具构建不同水鸟类群在生境节点间迁移的踏脚石网络, 该网络模拟了重要生境节点与踏脚石之间的物种交流。
1.4 网络的稳定性分析生境廊道具有空间连续性, 保障了水鸟在区域内活动。然而, 踏脚石分布在破碎的生境景观中, 其具有数量多, 形状和大小不一等特点。而物种沿着踏脚石移动的路径和方向是随机的。当区域内某一踏脚石丧失, 即代表网络中某一节点的丧失, 其对网络的鲁棒性具有重要影响[16]。为了模拟踏脚石丧失对网络的影响, 本研究设置了三种情景方案, 即随机破坏场景、人类干扰场景和最佳保护场景下节点移除规则对区域踏脚石网络稳定性的影响, 三种情景方案和描述见表 3。节点中心度量化了廊道内踏脚石网络的整体连通性, 其值越大表明该节点重要性越高且连接的网络数量越多。本研究节点中心度的计算是在Linkage Mapper软件中Centrality Mapper工具中进行[23]。
| 情景方案 Scenario |
模拟情景 Simulation scenario |
描述 Description |
原因 Reason |
| 1 | 随机破坏情景 | 随机移除踏脚石节点 | 潜在生境斑块被随机破坏 |
| 2 | 人类干扰情景 | 根据踏脚石面积由大到小的顺序移除节点 | 自然状态下, 较小的生境斑块更易被破坏 |
| 3 | 最佳保护情景 | 根据节点中心度大小移除节点 | 具有高中心性的垫脚石节点对区域生境网络的稳定性具有较高价值 |
为了模拟踏脚石网络稳定性, 本研究假设重要节点不会受到破坏, 仅对一般节点(即踏脚石)可能受到的破坏场景进行模拟, 以测算踏脚石不断丧失对区域生境网络稳定性产生的负面影响。为了衡量三个情景方案引发的踏脚石网络稳定性的变化, 本研究设置了四个网络指标, 即网络连接的鲁棒性、全局效率、割点和同配系数。其中, 网络连接的鲁棒性和全局效率反映了踏脚石网络的稳定性[24], 割点和同配系数反映了网络形态的变化[14, 25]。
网络连接的鲁棒性指标反映了网络破坏后系统保持稳定连接的能力, 该指标测量了网络损坏时的连接状态, 其计算公式如下[25]所示:
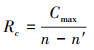
|
(2) |
式中, Cmax为踏脚石节点移除后网络的最大连通子图中节点的数量, n为网络中原始节点数量, n′为网络中被移除的节点数量。
全局效率(Global efficiency, Eglob)是描述网络内部结构非常重要的参数。该指标刻画了网络中最短路径的传输效率。最短路径长度越短, 网络全局效率越高, 网络传递信息速率越快。因此, 该指标反映了网络损坏后网络节点间的传输效率。其数学表达式为:
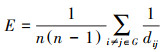
|
(3) |
式中, G为网络中的节点集, i和j是网络节点集G中任意两个节点, dij是节点i和j之间最短路径的长度。
割点的涵义是当某一节点被移除时, 网络将分为两个或多个子网络, 那么该节点被称之割点[25]。该指标是表征节点重要性的一个指标, 其反映了连接网络的脆弱程度。
同配系数用来刻画网络的同配性, 其测量了具有相近度值的节点是否倾向于相互连接, 该指标反映了移除节点后网络的形状, 其数学表达式为[14]:
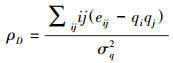
|
(4) |
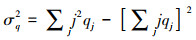
|
(5) |
式中, eij为在一个随机两个节点余度的联合概率分布, q为余度的标准化分布, σq2为余度qj的方差。同配系数取值在[-1, 1], 若同配系数为正, 则为同配网络, 反之为异配网络, 其绝对值大小表示网络属于同配或异配网络的强弱程度。本研究复杂网络分析利用R语言igraph包进行相关计算。
2 结果与分析 2.1 多类群水鸟生境及潜在廊道分布 2.1.1 栖息地阻力面和重要生境分析研究区鸻鹬类、雁鸭类和鹭类种群和生境主要分布于北缓至南缓栖息地、大丰至条子泥栖息地一带。生态阻力面分析结果显示(图 2), 研究区生态阻力值较小的区域主要集中在五个栖息地内、南缓栖息地与大丰港之间以及大丰栖息地与条子泥栖息地中间地区, 较小的阻力值表明这些区域生境质量较高, 三类水鸟分布广泛。

|
| 图 2 水鸟栖息地阻力面和电流分布 Fig. 2 Resistance surface and flow distribution of waterbird habitats |
重要生境区分布显示(图 2), 三类水鸟重要生境区主要分布在各栖息地内部以及大丰至条子泥栖息地一带。从数量和面积统计看出(表 4), 北部生境区数量为18个, 而南部生境区数量仅6个。然而, 北部生境区总面积和平均面积均低于南部生境区。北部生境区总面积为5509.19 hm2, 相当于南部生境区的1/2, 而平均面积仅相当于1/6。上述结果表明南部区域单一生境面积更大, 且生境利用更加集中。高电流值区域主要分布在各栖息地内重要生境区周边, 这些区域普遍生态阻力较小, 便于水鸟在各生境区内迁移。
| 重要生境区 Important habitat |
数量 Quantity |
总面积 Total area/hm2 |
平均面积 Average area/hm2 |
面积占比 Proportion/% |
| 北部Northern area | 18 | 5509.19 | 306.07 | 1.84 |
| 南部Southern area | 6 | 10934.69 | 1822.45 | 3.65 |
根据公式(1)得到水鸟栖息地潜在廊道电流值(图 3), 可以发现其电流值较高的廊道主要分布在南缓栖息地西部以及大丰栖息地北部和南部区域。而从踏脚石分布来看(图 3), 其主要分布在重要生境区附近, 不同区域分布特征存在差异。其中, 北部踏脚石主要分布在各栖息地内部, 且分布均匀。南部踏脚石主要集中在大丰栖息地及其以南的区域, 条子泥栖息地内踏脚石较少, 空间上呈现出明显的北多南少的格局。

|
| 图 3 水鸟栖息地潜在廊道和踏脚石 Fig. 3 Potential corridors and stepping stones for waterbird habitat |
从踏脚石数量和面积来看(表 5), 北部生境区踏脚石数量为96个, 总面积达到8080.70 hm2, 而南部生境区为80个, 总面积为6802.78 hm2, 踏脚石数量和面积均低于北部生境区。而从踏脚石平均面积来看, 南北部踏脚石平均面积相近, 分别为84.17 hm2和85.03 hm2, 这表明南部生境区虽然踏脚石数量较少, 但是单一踏脚石斑块面积更大。
| 栖息地踏脚石群 Habitat stepping stone groups |
数量Quantity | 总面积 Total area/hm2 |
平均面积 Average area/hm2 |
面积占比 Proportion/% |
| 北部Northern area | 96 | 8080.70 | 84.17 | 2.70 |
| 南部Southern area | 80 | 6802.78 | 85.03 | 2.27 |
从踏脚石类型来看(图 4), 北部踏脚石主要以养殖塘、芦苇沼泽和农田为主, 面积分别为2759.57 hm2、1550.97 hm2和1309.95 hm2, 南部踏脚石以光滩和养殖塘为主, 其面积分别为2482.34 hm2和1965.06 hm2。南北踏脚石仍以自然和人工生境类型为主, 非生境类型踏脚石面积较小。

|
| 图 4 水鸟栖息地踏脚石类型面积统计 Fig. 4 Statistics of the types and areas of stepping stones of waterbird habitats |
研究区鸻鹬类、雁鸭类和鹭类水鸟重要生境之间通过踏脚石网络进行连接(图 5), 重要生境区面积较大, 具有较强的景观稳定性, 支撑了鸻鹬类和雁鸭类水鸟的活动。而踏脚石位于重要生境区之间, 其斑块面积普遍较小, 具有一定的脆弱性。本研究将重要生境视为重要节点, 将踏脚石视为一般节点, 通过节点间的空间连接情况来反映区域水鸟栖息地保护网络现状。

|
| 图 5 水鸟栖息地潜在廊道的踏脚石网络 Fig. 5 Stepping stone networks of potential corridors for waterbird habitats |
踏脚石网络的空间分布显示(图 5), 踏脚石网络由35个重要节点和153个一般节点组成, 而这些节点由404个活跃网络和31个非活跃网络构成。其中, 北部网络(图 5)由25个重要节点和93个一般节点组成, 包含259个活跃网络和21个非活跃网络。南部网络(图 5)由10个重要节点和60个一般节点组成, 包含了145个活跃网络和10个非活跃网络。上述结果表明北部网络连接性更好, 其生境间的连通性水平更高, 水鸟在该区生境踏脚石的选择性更加多样。
2.3 不同情境下生境踏脚石网络稳定性分析 2.3.1 踏脚石网络现状分析根据自然断点法将踏脚石节点中心度分为五个级别(图 6), 踏脚石节点中心度的分布与踏脚石空间分布特征一致, 即南北生境区密度大, 中间区域密度小。其中, 北部生境区(图 6)节点中心度分布较为均为, 主要分布在生境廊道内, 廊道外节点度值较低。廊道内高的中心度值主要分布在踏脚石密度较大的区域, 重要生境与踏脚石之间的连通性水平较高。南部生境区(图 6)节点中心度空间分布呈现北多南少的格局。其中, 北部节点中心度的高值点较多, 而南部条子泥栖息地内部节点中心度以低的度值点为主。条子泥栖息地重要生境面积较大且以光滩为主, 对鸻鹬类等小型涉禽觅食和栖息的支撑能力较强, 因此小斑块的踏脚石对鸻鹬类吸引力较弱。

|
| 图 6 节点中心度和生境保护廊道分布图 Fig. 6 Distribution map of habitat node centrality and habitat conservation corridor |
本研究利用四个网络参数来分析不同干扰情景下网络结构的稳定性差异。北部网络情景分析显示(图 7), 当按照踏脚石节点面积由大到小移除47.92%的节点后, 情景二网络连接的鲁棒性和全局效率呈现急剧下降, 其值仅0.58和0.09, 这反映出面积较大的踏脚石被移除后将对整个水鸟栖息地保护网络的连通性产生强烈的负面影响。随机破坏情景(情景一)下, 踏脚石网络连接的鲁棒性表现良好, 当随机节点被移除70.83%后网络的稳定性才发生剧烈变化。当移除70.83%的节点后全局效率累计下降值达到0.1, 表明网络的连通性水平随着节点的移除不断下降。最佳保护情景下(情景三)下, 网络连接的鲁棒性表现较好, 均高于0.91, 且全局效率呈逐渐上升的趋势, 这表明中心度值大的节点对整个栖息地网络连通性的维持起到了良好的支撑作用。从割点数量来看, 随着节点的移除, 情景一的割点数量率先达到峰值, 其次是情景二, 最后是情景三。上述结果表明随机移除节点会导致网络结构出现破坏, 亟需加强重要节点的保护力度, 防止栖息地网络结构的进一步破坏。从同配系数来看, 最佳保护情景下(情景三)网络形状异配转为同配后一直保持稳定, 且随着节点的移除同配强度逐渐增加, 这表明随着低的中心度值节点被移除, 网络形状和结构更加稳定, 区域内栖息地保护网络具有较强的抗破坏能力。

|
| 图 7 北部生境区在不同情境下生境网络参数 Fig. 7 Habitat network parameters of Northern habitat area in different scenarios |
与北部网络结果不同的是, 南部网络随机破坏情景(情景一)和最佳保护情景(情景三)下网络连接的鲁棒性出现剧烈变化对应的节点移除数量更少, 且全局效率变化更为剧烈(图 8)。当踏脚石节点过少或分布不均匀, 任意的节点移除都会对网络整体的稳定性和连通性产生重要影响。情景一和情景二割点变化曲线相似, 网络结构较北部生境区更为脆弱。同配系数表现较北部生境区更加剧烈, 且三种情景下网络的同配和异配性快速更迭, 区域栖息地网络稳定性较差。

|
| 图 8 南部生境区在不同情境下生境网络参数 Fig. 8 Habitat network parameters of Southern habitat area in different scenarios |
踏脚石能有效支持水鸟种群的外溢。由于鸻鹬类、雁鸭类和鹭类是研究区主要的水鸟类群, 数量占比约70%—80%, 因此该三类水鸟栖息地网络研究对区域水鸟多样性水平提升意义重大。图 5显示出大丰港将潜在生态廊道分割成南北两大块, 且北部踏脚石节点密度较高面积却较小(表 4), 这与南北生境区主导水鸟类群的差异有关。其中, 北生境区是以雁鸭类和鹭类为主, 主要栖息在芦苇沼泽、农田和养殖塘。而北缓栖息地的农作物以及核心栖息地芦苇成为其觅食和栖息主要场所。然而, 雁鸭类对环境因素的阈值响应表明[7], 其利用斑块普遍较小且分布分散, 因此重要生境总面积仅5509.19 hm2(表 4)。相比而言, 南部生境区水鸟重要生境面积更大, 达到10934.69 hm2, 这与鸻鹬类生境利用类型有关, 潮间带滩涂和养殖塘内鱼、虾、螺等资源非常丰富, 吸引了鸻鹬类在此聚集停留[6, 11]。
踏脚石节点分布和结构稳定性是本研究的主要发现。具体来说, 在最优保护情景下北部网络整体的连通效率较高(图 7)。然而, 在人类干扰情景下面积较大的踏脚石斑块被移除后网络连接的鲁棒性快速下降, 连通的效率呈现断崖式下降, 这突出了斑块面积较大的踏脚石节点的重要价值。因此, 今后要防范大面积踏脚石斑块的丧失对整个栖息地网络稳定性产生影响[5]。从生态学意义来说, 面积较大的踏脚石节点形成了对现有重要生境的补充[26]。研究区雁鸭类和鹭类越冬期的数量可达数万只, 均集中在北缓至南缓栖息地这一狭长的滨海湿地内[11]。而雁鸭类普遍选择靠近水源的中低盖度植被生境。鹭类的环境适应力更强, 芦苇沼泽和养殖塘均是其栖息生境。而且, 鹭类广泛分布在北生境区的各个角落, 且分布更加分散, 空间选择性更强。同时, 在各重要生境区之间密集的踏脚石也构成了局部区域稳定的生境保护网络[27]。虽然北部扩展生境水鸟利用率较低, 但是也为水鸟提供了重要的缓冲带。尤其, 这些扩展生境人类开发强度低, 通过后期的生境改造, 可以营造为水鸟踏脚石节点或重要生境区, 增强北生境区栖息地保护网络的稳定性。
南部生境区是鸻鹬类北上重要的觅食通道。条子泥栖息地受潮汐作用, 鸻鹬类来回穿梭于海堤内外光滩与扬水滩[1]。从模拟情景来看, 南部网络结构稳定性和连通性比北生境区差, 鸻鹬类高度聚集在条子泥光滩和高潮栖息地中, 养殖塘仅作为觅食补充地或北迁的跳板[12, 28]。同时, 大丰与条子泥之间潜在廊道狭窄脆弱, 受外力影响极易造成南北生境区空间隔离。然而, 这种踏脚石节点的破坏对鸻鹬类的影响目前难以进行量化和预测。因此优化和维持现有条子泥栖息地内以光滩和半自然生境为主的重要生境区和踏脚石斑块是南部生境区水鸟栖息地网络保护的主要方向和目标。
3.2 维持和优化多类群水鸟栖息地保护网络结构的稳定性和连通性根据情景三模拟的结果显示当踏脚石中心度值较小的节点被移除后网络的连通性增强且结构稳定。为此, 本研究将踏脚石网络中心度值较小的节点(第一级和第二级)进行移除。同时, 考虑到面积较大的踏脚石节点在网络稳定性方面的重要意义[21, 29], 根据情景二中较大面积踏脚石节点移除后网络连接的鲁棒性分析结果, 保留面积大于31.98 hm2且中心度值较小的踏脚石节点, 最后得到优化后的多类群水鸟栖息地保护网络(图 9)。

|
| 图 9 优化后的多类群水鸟栖息地保护网络 Fig. 9 Waterbird Habitat conservation after optimization |
优化后的踏脚石网络节点中心度平均值明显提高, 其中, 北部生境区优化后的踏脚石节点中心度平均值为1107.42, 比优化前高123.22;而南部生境区中心度比优化前高57.47。除了中心度均值提高以外, 优化后北部生境区踏脚石节点数量由93个下降为78个, 南部生境区踏脚石节点数量由60个下降47个, 节点数量分别下降了16.13%和21.67%。随着冗余踏脚石节点的移除, 北部生境区的空间连通性提高。其中, 大丰栖息地南部两条廊道中西部廊道宽度明显变窄, 该区域主要以养殖塘生境为主, 节点中心度值普遍较小且面积较小。条子泥栖息地西部在优化后保护廊道面积变小, 该处人类活动强度逐年增强[8]。作为鸻鹬类高潮时短暂觅食停歇地, 具有高度的可替代性[30—31]。随着该处冗余踏脚石节点的移除, 条子泥栖息地生境保护廊道南边界北移, 整个廊道内踏脚石节点和网络得到优化, 区域水鸟栖息地保护网络结构更加稳定, 重要生境区之间的连通性增强。
4 结论本研究运用踏脚石识别相关原理、生态阻力面、电路理论和复杂网络方法, 构建了区域水鸟栖息地网络, 并对现状网络节点和结构进行了稳定性分析, 得到以下结论:(1)多类群水鸟共有24个重要生境节点和176个踏脚石节点, 北部生境节点和踏脚石平均面积为306.07 hm2和84.17 hm2, 而南部为1822.45 hm2和85.03 hm2;(2)踏脚石节点数量呈现北多南少的格局, 条子泥栖息地内踏脚石节点较少, 且以低的度值为主;(3)最佳保护情境下北部和南部生境区网络结构稳定性最好, 情景一下南部栖息地网络中踏脚石节点变化对网络整体的稳定性和连通性产生重要影响;(4)根据情景二模拟结果, 本研究优化了多类群水鸟栖息地网络, 优化后北部生境区踏脚石节点中心度均值提高了123.22, 踏脚石数量下降为78个, 南部生境区中心度均值提高了57.47, 踏脚石数量下降到47个, 冗余踏脚石节点移除后栖息地网络空间连通性水平增强。
| [1] |
Duan H L, Xia S X, Jackson M V, Zhao N, Liu Y, Teng J K, Meng Z, Yu X B, Shi J B. Identifying new sites of significance to waterbirds conservation and their habitat modification in the Yellow and Bohai Seas in China. Global Ecology and Conservation, 2020, 22: e01031. DOI:10.1016/j.gecco.2020.e01031 |
| [2] |
Ma T T, Li X W, Bai J H, Cui B S. Habitat modification in relation to coastal reclamation and its impacts on waterbirds along China's coast. Global Ecology and Conservation, 2019, 17: e00585. DOI:10.1016/j.gecco.2019.e00585 |
| [3] |
Hou X Y, Wu T, Hou W, Chen Q, Wang Y D, Yu L J. Characteristics of coastline changes in mainland China since the early 1940s. Science China Earth Sciences, 2016, 59(9): 1791-1802. DOI:10.1007/s11430-016-5317-5 |
| [4] |
Wang G, Wang C, Guo Z R, Dai L J, Wu Y Q, Liu H Y, Li Y F, Chen H, Zhang Y N, Zhao Y X, Cheng H, Ma T W, Xue F. A multiscale approach to identifying spatiotemporal pattern of habitat selection for red-crowned cranes. Science of the Total Environment, 2020, 739: 139980. DOI:10.1016/j.scitotenv.2020.139980 |
| [5] |
欧维新, 袁薇锦. 基于景观连接度的盐城滨海湿地丹顶鹤生境斑块重要性评价. 资源科学, 2015, 37(4): 823-831. |
| [6] |
颜凤, 刘本法, 余仁栋, 乔亚军, 安树青, 苏安劼. 围填海对盐城珍禽自然保护区越冬水鸟群落及空间分布的影响. 生态科学, 2018, 37(6): 20-29. |
| [7] |
王成, 刘红玉, 李玉凤, 王刚, 董斌, 陈浩, 张亚楠, 赵永强. 盐城滨海湿地水鸟类群生境适宜性及生态阈值研究: 对栖息地景观结构恢复的启示. 生态与农村环境学报, 2022, 38(7): 897-908. |
| [8] |
张晓祥, 严长清, 徐盼, 戴煜暄, 晏王波, 丁贤荣, 朱晨曦, 梅丹丹. 近代以来江苏沿海滩涂围垦历史演变研究. 地理学报, 2013, 68(11): 1549-1558. |
| [9] |
Herrera L P, Sabatino M C, Jaimes F R, Saura S. Landscape connectivity and the role of small habitat patches as stepping stones: an assessment of the grassland biome in South America. Biodiversity and Conservation, 2017, 26(14): 3465-3479. DOI:10.1007/s10531-017-1416-7 |
| [10] |
Merken R, Deboelpaep E, Teunen J, Saura S, Koedam N. Wetland suitability and connectivity for trans-Saharan migratory waterbirds. PLoS One, 2015, 10(8): e0135445. DOI:10.1371/journal.pone.0135445 |
| [11] |
任武阳. 滨海湿地生境多样性及其对越冬水鸟栖息地利用影响研究——以盐城保护区为例[D]. 南京: 南京师范大学, 2019.
|
| [12] |
Yang Z Y, Lagassé B J, Xiao H, Jackson M V, Chiang C Y, Melville D S, Leung K S K, Li J, Zhang L, Peng H B, Gan X J, Liu W L, Ma Z J, Choi C Y. The southern Jiangsu coast is a critical moulting site for Spoon-billed Sandpiper Calidris pygmaea and Nordmann's Greenshank Tringa guttifer. Bird Conservation International, 2020, 30(4): 649-660. DOI:10.1017/S0959270920000210 |
| [13] |
阮得孟, 孙勇, 程嘉伟, 刘大伟, 鲁长虎. 盐城自然保护区新洋港河口不同生境冬季鸟类群落组成及其梯度变化. 生态学报, 2015, 35(16): 5437-5448. |
| [14] |
汪小帆, 李翔, 陈关荣. 网络科学导论. 北京: 高等教育出版社, 2012.
|
| [15] |
裴燕如, 孙炎浩, 于强, 马骏, 王慧媛, 岳德鹏. 黄河流域典型矿区生态空间网络优化——以鄂榆地区为例. 煤炭学报, 2021, 46(5): 1541-1554. |
| [16] |
傅丽华, 莫振淳, 彭耀辉, 谢美, 高兴燕. 湖南茶陵县域生态空间网络稳定性识别与重构策略. 地理学报, 2019, 74(7): 1409-1419. |
| [17] |
刘红玉, 周奕, 郭紫茹, 戴凌骏, 王成, 王刚, 李玉凤. 盐沼湿地大规模恢复的概念生态模型——以盐城为例. 生态学杂志, 2021, 40(1): 278-291. |
| [18] |
Peng J, Yang Y, Liu Y X, Hu Y N, Du Y Y, Meersmans J, Qiu S J. Linking ecosystem services and circuit theory to identify ecological security patterns. Science of the Total Environment, 2018, 644: 781-790. |
| [19] |
Urban D, Keitt T. Landscape connectivity: a graph-theoretic perspective. Ecology, 2001, 82(5): 1205-1218. |
| [20] |
宋利利, 秦明周. 整合电路理论的生态廊道及其重要性识别. 应用生态学报, 2016, 27(10): 3344-3352. |
| [21] |
华彦, 李国忠, 高桂华. 踏脚石原理在自然保护区设计上的应用. 野生动物, 2005, 26(4): 30-31. |
| [22] |
邬建国. 景观生态学: 格局、过程、尺度与等级 (2版). 北京: 高等教育出版社, 2007.
|
| [23] |
McRae B H. Centrality Mapper Connectivity Analysis Software. (2012-08-09) http://www.Circuitscape.org/linkagemapper.
|
| [24] |
Ventresca M, Aleman D. Network robustness versus multi-strategy sequential attack. Journal of Complex Networks, 2015, 3(1): 126-146. |
| [25] |
Tian L, Bashan A, Shi D N, Liu Y Y. Articulation points in complex networks. Nature Communications, 2017, 8(1): 1-9. |
| [26] |
吴未, 廉文慧. 生态系统服务功能视角下建设用地扩张对生境网络的影响——以苏锡常地区白鹭栖息地为例. 长江流域资源与环境, 2018, 27(5): 1043-1050. |
| [27] |
Shi F N, Liu S L, Sun Y X, An Y, Zhao S, Liu Y X, Li M Q. Ecological network construction of the heterogeneous agro-pastoral areas in the upper Yellow River Basin. Agriculture, Ecosystems & Environment, 2020, 302: 107069. |
| [28] |
Hu W, Chen T Y, Xu Z P, Wu D W, Lu C H. Occurrence dataset of waterbirds in the Tiaozini Wetland, a World Nature Heritage, China. Biodiversity Data Journal, 2022, 10: e90724. |
| [29] |
张启舜, 李飞雪, 王帝文, 李满春, 陈东. 基于生态网络的江苏省生态空间连通性变化研究. 生态学报, 2021, 41(08): 3007-3020. |
| [30] |
牛一醒, 陈巧尔, 高帅, 高征. 江苏东台条子泥湿地水鸟群落的季节变化和生境利用状况. 湿地科学, 2022, 20(03): 357-365. |
| [31] |
高帅, 刘威, 张帅, 伊剑锋, 王征. 江苏东台条子泥和如东湿地越冬水鸟多样性研究. 生态与农村环境学报, 2021, 37(09): 1176-1182. |