 2023, Vol. 43
2023, Vol. 43文章信息
- 肖昱承, 王端阳, 陈波, 胡光镇, 马云鹤, 杨雯, 赵景学
- XIAO Yucheng, WANG Duanyang, CHEN Bo, HU Guangzhen, MA Yunhe, YANG Wen, ZHAO Jingxue
- 短期增温对西藏念青唐古拉山典型植物群落结构和稳定性的影响
- Effects of short-term warming on structure and stability of typical plant communities in Nyenchenthanglha Mountain, Tibet
- 生态学报. 2023, 43(20): 8608-8619
- Acta Ecologica Sinica. 2023, 43(20): 8608-8619
- http://dx.doi.org/10.20103/j.stxb.202210313100
-
文章历史
- 收稿日期: 2022-10-31
- 采用日期: 2023-07-21

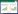




2. 兰州大学生命科学学院, 兰州 730000;
3. 草种创新与草地农业生态系统全国重点实验室, 兰州 730000
2. College of Life Sciences, Lanzhou University, Lanzhou 730000, China;
3. Statel Key Laboratory of Herbage Improvement and Grassland Agro-ecosystems, Lanzhou 730000, China
全球气候变暖显著地改变着陆地生态系统的结构和功能。青藏高原作为世界屋脊和地球第三极, 由于其生态系统脆弱, 且对气候变暖极其敏感, 近年来受到众多学者的广泛关注[1]。在过去几十年间, 青藏高原气候变暖幅度显著高于全球其他区域, 这将对高寒草地植物群落的结构和稳定性产生很大影响。反之, 高寒草地生态系统的微小变化也会反馈到全球气候变化[2—3]。
气候变暖对植物群落的影响机制包括直接和间接两方面[4—5]。直接影响包括与温度有关的光合[6]、呼吸等生理生化过程及其各种酶和代谢产物等的变化[7]。间接影响包括因土壤理化性质等变化引起的植物对水分、养分吸收利用, 植物生长发育繁殖[8—9]乃至群落演替等方面的影响[10]。增温会改变高寒草甸群落的高度、密度、盖度、频度和重要值, 从而影响其群落结构和生态系统功能[11]; 同时, 增温会使植物生物量分配明显趋向土壤深层。此外, 增温也会改变物种生态位, 进而影响群落演替过程中种群的环境适应性和资源利用能力[12], 以及物种多样性和物种共存特征[13]。
物种多样性是影响群落稳定性的一个重要因素。Alexandra等[14]的研究发现, 植物群落生物量的空间稳定随着物种多样性的增加而增加。Chalcarft[15]实验发现, 在样地尺度, 生态系统空间稳定性与物种多样性呈正线性相关关系, 但种群空间稳定性则随物种多样性的增加而降低; 而在区域尺度, 生态系统和种群的空间稳定性均随物种多样性的增加而增加。
青藏高原拥有典型的高寒草地生态系统, 沿着山体垂直带, 由于植物群落在低海拔普遍受干旱胁迫, 而在高海拔主要受低温限制, 植被生产力和群落物种丰富度随海拔升高普遍呈先增加后降低的单峰分布格局[16—18]。气候变暖会改变这种水热因子对植被生长的限制, 影响植物群落结构和生产力。近年来, 在青藏高原高寒草地生态系统, 多名学者利用开顶式增温箱(OTC)开展了气候变暖对植物群落结构[11, 19]、植物生理生化过程[20, 21]、土壤理化性质[19]和微生物群落结构[22—23]等影响的研究工作。
沿着同一山体垂直样带, 温度、水分等环境因子随着海拔梯度协同变化且呈现出不同水热组合形式, 结合增温控制实验, 可为典型高山植物群落结构及其稳定性对气候变暖响应机制研究提供天然平台[24—25]。西藏念青唐古拉山脉位于青藏高原腹地, 其中段是高寒草甸向高寒草原的过渡区域, 对气候变化极为敏感和脆弱。然而, 对西藏念青唐古拉山这同一高大山体海拔梯度上、不同典型植物群落结构及其空间稳定性的研究还未见报道。本研究基于沿山体垂直带的OTC增温实验平台, 选择西藏念青唐古拉山高寒草原、高寒草甸和流石滩三种典型植物群落, 开展植物群落结构、物种多样性、生物量、物种生态位宽度和群落稳定性等方面的定量对比研究, 探讨增温对同一山体海拔梯度上、不同植物群落结构及其空间稳定性的影响, 可为探讨青藏高原地区应对和适应未来气候变化提供科学依据。
1 材料和方法 1.1 研究地点概况研究区位于西藏自治区拉萨市当雄县境内的念青唐古拉山南坡(30°30′N, 91°03′E)(图 1)。受大气环流和地形影响, 研究区降水主要集中在夏季, 冬季寒冷干燥, 雨热同期, 干湿季分明[26]; 年平均气温为1.8℃, 最冷月(1月)平均气温-9℃, 最热月(7月)平均气温为11.1℃, 平均年降水量为442mm, 90%的降水集中在5—9月植被生长季期间(1963—2013)[27]。

|
| 图 1 研究区域及样地概况 Fig. 1 Overview of the study area and sample plot 图中从左到右, 从上到下依次为:研究区域、样地景观、实验设计示意图、开顶式(OTC)增温装置 |
实验选择念青唐古拉山南坡的高寒草原(海拔4500m)、高寒草甸(海拔4900m)以及流石滩(海拔5300m)三个典型植物群落类型作为研究对象。其中高寒草原的优势种为高山嵩草(Kobresia pygmaea)、丝颖针茅(Stipa capillacea)和圆萼刺参(Morina chinensis), 植被盖度在15%—30%以上; 高寒草甸的优势种为高山嵩草、粗壮嵩草(Carex brunnescens)和灰毛蓝钟花(Cyananthus incanus), 植被盖度最大可达90%;流石滩的优势种为高山嵩草、紫花针茅(Stipa purpurea)、钉柱委陵菜(Potentilla saundersiana)和山居雪灵芝(Arenaria edgeworthiana), 植被盖度在60%左右[28]。
1.2 实验设计分别在三种植物群落类型区设置一个20m×20m的围栏样地, 设置2种处理(对照和增温), 每个处理设计有6个重复, 共计12个小区, 对照和增温处理交替且均匀分布, 每个小区大小为2m×2m。开顶式增温装置(OTC)于2020年5月安装, 常年持续使用, 采用聚碳酸酯板(透光率>95%)材质, 高48cm, 顶部口径100cm, 底部口径150cm, 增温幅度为2.2—3.2℃(图 1)。
1.3 植物群落调查与取样2021年, 在每一个小区中设置50cm×50cm的样方(图 1), 记录样方内出现的物种名称、每种植物的绝对高度和每种植物的分盖度及样方的总盖度。对样方中出现的植物物种按功能群进行分类, 分为莎草类(Sedges)、禾本类(Grasses)、豆科(Legumes)、垫状植物(Cushions)、杂草(Forbs)五大类。为保证实验延续性和避免破坏样地, 地上生物量参考王忠[28]方法进行计算, 即利用样方调查得到的物种高度、分盖度建立生物体积与地上生物量的估算方程。地下生物量通过采集土柱(深度20cm, 直径3.5cm), 并用2mm网筛进行冲洗, 得到的根系经65 ℃烘箱干燥48小时后称重进行估算。
1.4 参数计算方法 1.4.1 重要值(IV)利用单个物种的相对盖度(RC)、相对高度(RH)、相对生物量(RB)计算各物种的重要值。

|
(1) |
式中, RC为某一物种盖度/所有物种的盖度和×100%;RH为某一物种个体高度/所有物种个体高度之和×100%;RB为某一物种生物量/所有物种生物量之和×100%
1.4.2 物种多样性指数(1) Shannon-Wiener多样性指数(H):

|
(2) |
(2) Pielou均匀度指数(E):

|
(3) |
式中, Pi为第 i个物种数量占群落总个体数量的比例; S为样地内物种丰富度。
1.4.3 物种生态位宽度本文采用两种不同物种生态位的计算方法, 能够更全面地反映该种群对环境适应的状况或对资源利用的程度, 计算方法如下。
(1) Levins生态位宽度(BL):

|
(4) |
(2) Shannon生态位宽度(BS):

|
(5) |
式中, Pij为第i个物种在第j个资源水平下的重要值占该物种在所有资源水平上的重要值总和的比例; r为样方总数。
1.4.4 群落空间稳定性(ICV)

|
(6) |
式中, μ为样方群落总盖度的平均值; σ为标准差。
1.5 数据分析方法使用SPSS 26.0进行了最小显著性差异(LSD)多重比较分析, 探讨在同一处理下不同植物群落间的差异和同一植物群落的不同处理下的差异; 双因素方差分析探讨增温和植物群落对群落特征的影响。图片采用R 4.1.2中的ggplot2包进行制作。
2 结果分析 2.1 增温对三种典型植物群落物种组成和结构的影响在高寒草原中, 对照样地有20个物种, 增温样地有16个物种。与对照样地相比, 增温样地增加了青藏薹草(Carex moorcroftii)、西藏三毛草(Trisetum tibeticum)、蕨麻(Potentilla anserina)这3个物种, 减少了山居雪灵芝、钉柱委陵菜、冰川棘豆(Oxytropis glacialis)、蓝玉簪龙胆(Gentiana veitchiorum)、马先蒿(Pedicularis resupinata)、垫状点地梅(Androsace tapete)、灰毛蓝钟花这7个物种。在高寒草甸中, 对照样地有18个物种, 增温样地有22个物种。与对照样地相比, 增温样地增加了青藏薹草、矮生嵩草(Kobresia humilis)、紫花针茅、高山豆(Tibetia himalaica)、垫状点地梅、西藏风毛菊(Saussurea tibetica)、肉果草(Lancea tibetica)、狼毒(Stellera chamaejasme)、大戟(Euphorbia pekinensis)这9个物种, 减少了高山棘豆(Oxytropis alpina)、蓝玉簪龙胆、葶苈(Draba nemorosa)、珠芽蓼(Polygonum viviparum)、二裂委陵菜(Potentilla bifurca)这5个物种。在流石滩中, 对照样地有13个物种, 增温样地有15个物种。与对照样地相比, 增温样地增加了高山唐松草(Thalictrum alpinum)、湿生萹蕾(Gentianopsis paludosa)、冰岛蓼(Koenigia islandica)这3个物种, 减少了大戟这1个物种(附表 1)。
在三种典型植物群落中, 增温样地较对照样地增减的物种多为偶见种和伴生种, 这些物种在群落中的数量较少, 所占比例不大, 对群落的组成和结构影响较小。但是增温改变了各物种的重要值, 进而群落的优势物种发生改变。如表 1所示, 在高寒草原中, 优势物种由高山嵩草、丝颖针茅和圆萼刺参演变为青藏薹草、粗壮嵩草、丝颖针茅和火绒草(Leontopodium leontopodioides); 在高寒草甸中, 优势物种由粗壮嵩草、高山嵩草和灰毛蓝钟花演变为高山嵩草; 在流石滩中, 优势物种由高山嵩草、紫花针茅、山居雪灵芝和钉柱委陵菜演变为高山嵩草、钉柱委陵菜和小叶金露梅(Potentilla parvifolia)。
| 物种 Species |
高寒草原 Alpine steppe | 高寒草甸 Alpine meadow | 流石滩 Screes | |||||
| 对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
|||
| 青藏薹草 Carex moorcroftii | — | 0.10 | — | 0.09 | — | — | ||
| 粗壮嵩草 Kobresia robusta | 0.06 | 0.17 | 0.11 | 0.08 | — | — | ||
| 高山嵩草 Kobresia pygmaea | 0.27 | 0.03 | 0.23 | 0.26 | 0.10 | 0.21 | ||
| 丝颖针茅 Stipa capillacea | 0.11 | 0.14 | 0.03 | 0.07 | 0.04 | 0.04 | ||
| 紫花针茅 Stipa purpurea | 0.05 | 0.05 | — | 0.01 | 0.17 | 0.06 | ||
| 山居雪灵芝 Arenaria edgeworthiana | — | — | — | — | 0.21 | 0.08 | ||
| 钉柱委陵菜 Potentilla saundersiana | 0.01 | — | 0.07 | 0.04 | 0.11 | 0.13 | ||
| 小叶金露梅 Potentilla parvifolia | — | — | — | — | 0.08 | 0.19 | ||
| 火绒草 Leontopodium leontopodioides | 0.01 | 0.18 | — | — | — | — | ||
| 灰毛蓝钟花 Cyananthus incanus | 0.02 | — | 0.12 | 0.08 | — | — | ||
| 圆萼刺参 Morina chinensis | 0.10 | 0.01 | — | — | — | — | ||
| 表中, “—”表示该物种在对照或增温处理下的该植物群落中无分布 | ||||||||
在增温处理下, 高寒草甸的盖度显著降低(P < 0.05), 但对其他两个群落的株高和盖度影响不显著。在对照条件下, 高寒草原的高度显著高于高寒草甸和流石滩(P < 0.05);高寒草原和高寒草甸的高度显著高于流石滩(P < 0.05)。在增温条件下, 高寒草甸和高寒草原的盖度显著高于流石滩(P < 0.05);高寒草甸和流石滩的盖度显著高于高寒草原(P < 0.05)(图 2)。

|
| 图 2 增温对三种典型植物群落特征的影响 Fig. 2 Effects of warming on the characteristics of three typical plant communities ns表示同一类型植物群落的增温和对照之间无显著性差异; * * *(0.01>P)、* *(0.01>P>0.01)、*(0.05>P>0.01)表示同一类型植物群落的增温和对照之间有显著性差异; 不同小写字母表示在对照处理下的两种植物群落类型之间存在显著性差异, 反之无显著性差异; 不同大写字母表示在增温处理下的两种植物群落类型之间存在显著性差异, 反之无显著性差异; F′图为F图高寒草原和高寒草甸在对照和增温处理下的地下生物量 |
对三种典型植物群落的高度和盖度进行植物群落和增温的双因素方差分析(表 2), 增温对群落的盖度影响显著(P < 0.05), 植物群落及其与增温的交互作用对群落的高度、盖度存在显著性影响(P>0.05)。
| 影响因子 Impact factor |
植物群落 Plant community |
增温 Warming |
植物群落×增温 Plant community× Warming |
| 高度 Height | 35.126*** | 1.346 | 4.274* |
| 盖度 Coverage | 17.221*** | 11.114** | 3.868* |
| Shannon-Weiner指数 Shannon-Weiner index | 14.983*** | 3.122 | 0.297 |
| Pielou均匀度 Pielou homogeneous degree | 0.09 | 1.480 | 1.572 |
| 地上生物量 Aboveground biomass | 31.085*** | 15.969*** | 4.059* |
| 地下生物量 Belowground biomass | 108.497*** | 18.303*** | 20.673*** |
| Levins生态位宽度指数 Levins niche breadth index | 6.955** | 2.154 | 0.363 |
| Shannon生态位宽度指数 Shannon niche breadth index | 5.046** | 0.581 | 0.738 |
| 表中, 数字为双因素方差分析的F值; * * *(0.001>P)、* *(0.01>P>0.01)、*(0.05>P>0.01)表示该影响因子对群落特征存在显著影响, 未标注表示无显著影响 | |||
在同一类型植物群落中, 增温后群落的Shannon-Wiener指数呈降低趋势, 但这种影响并不显著(P>0.05);增温对群落的Pielou均匀度无显著影响(P < 0.05)。在同一处理下, 高寒草甸的Shannon-Wiener指数最高, 与高寒草原存在显著性差异(P < 0.05), 但与流石滩之间无显著性差异(P>0.05);各群落的Pielou均匀度均无显著性差异(P>0.05)(图 2)。在双因素方差分析中, Shannon-Wiener指数在不同类型植物群落间存在显著影响(P < 0.05), 增温及其与植物群落的交互作用均对两个物种多样性指数无显著影响(P>0.05)(表 2), 这说明增温不会影响群落的物种多样性。
在同一植物群落类型中, 增温降低了群落的地上生物量, 但在流石滩中这种变化不显著(P>0.05);增温极显著增大了高寒草甸的地下生物量(P < 0.01)。在同一处理下, 高寒草甸的地上和地下生物量均最大, 除增温样地中高寒草甸与流石滩的地上生物量间无显著差异(P>0.05)外, 都与其他两个群落之间存在显著性差异(P < 0.05)。根据双因素方差分析(表 2), 群落的地上和地下生物量在不同植物群落间存在差, 增温对地上和地下生物量的影响极显著(P < 0.01), 二者的交互作用对地上生物量的影响显著(P < 0.05), 对地下生物量的影响极显著(P < 0.01)。这说明增温和植物群落类型会影响植物群落的生物量, 并影响地上地下生物量分配策略。
2.3 增温对物种生态位宽度和群落稳定性的影响由图 3和表 3知, 两种方法计算的生态位宽度在增温后均呈现下降趋势, 但并不显著(P>0.05)。同时, 在双因素方差分析中, 不同类型植物群落对两种生态位宽度指数影响显著(P < 0.05), 但增温及其与植物群落的交互作用对两种生态位宽度指数无显著影响(P> 0.05)(表 2), 这表明增温改变了群落中物种利用资源的能力。

|
| 图 3 增温对三种典型植物群落物种平均生态位宽度的影响 Fig. 3 Effects of warming on the averaged species niche width of three typical plant communities |
| 功能群 Function groups |
物种 Species |
生态位宽度Niche width | ||||||||||||||||
| Levins指数 BL | Shannon指数 BS | |||||||||||||||||
| 高寒草原 Alpine steppe |
高寒草甸 Alpine meadow |
流石滩 Screes |
高寒草原 Alpine steppe |
高寒草甸 Alpine meadow |
流石滩 Screes |
|||||||||||||
| 对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
|||||||
| 莎草类 Sedges |
青藏薹草 Carex moorcroftii | — | 2.88 | — | 3.99 | — | — | — | 1.08 | — | 1.39 | — | — | |||||
| 粗壮嵩草 Kobresia robusta | 2.81 | 2.81 | 5.39 | 2.78 | — | — | 1.07 | 1.06 | 1.73 | 1.06 | — | — | ||||||
| 黑褐薹草 Carex atrofusca | 4.20 | 1.00 | 3.27 | 1.00 | — | — | 1.51 | — | 1.28 | — | — | — | ||||||
| 高山嵩草 Kobresia pygmaea | 5.32 | 1.00 | 5.79 | 5.64 | 4.17 | 4.20 | 1.73 | — | 1.77 | 1.76 | 1.51 | 1.50 | ||||||
| 矮生嵩草 Kobresia humilis | — | — | — | 2.87 | — | — | — | — | — | 1.07 | — | — | ||||||
| 禾本类 Grasses |
西藏三毛草 Trisetum tibeticum | — | 2.94 | — | — | — | — | — | 1.09 | — | — | — | — | |||||
| 丝颖针茅 Stipa capillacea | 2.28 | 2.46 | 2.87 | 3.74 | 1.00 | 1.68 | 0.95 | 1.00 | 1.08 | 1.35 | — | 0.59 | ||||||
| 紫花针茅 Stipa purpurea | 1.77 | 1.00 | — | 1.00 | 4.74 | 2.00 | 0.63 | — | — | — | 1.58 | 0.69 | ||||||
| 豆科植物 Legumes |
冰川棘豆 Oxytropis glacialis | 1.00 | — | 1.00 | 1.00 | — | — | — | — | — | — | — | — | |||||
| 高山棘豆 Oxytropis alpina | — | — | 1.00 | — | — | — | — | — | — | — | — | — | ||||||
| 高山豆 Tibetia himalaica | — | — | — | 1.00 | — | — | — | — | — | — | — | — | ||||||
| 垫状植物 Cushions |
点地梅 Androsace umbellata | — | — | 3.97 | 1.00 | 3.10 | 3.07 | — | — | 1.50 | — | 1.23 | 1.22 | |||||
| 山居雪灵芝 Arenaria edgeworthiana | 1.00 | — | — | — | 5.21 | 4.56 | — | — | — | — | 1.69 | 1.56 | ||||||
| 杂草类 Herbs |
二裂委陵菜 Potentilla bifurca | 1.00 | 1.66 | 1.00 | — | — | — | — | 0.59 | — | — | — | — | |||||
| 蕨麻 Potentilla anserina | — | 1.67 | 1.69 | 1.99 | — | — | — | 0.59 | 0.60 | 0.69 | — | — | ||||||
| 乳白香青 Anaphalis lactea | 2.08 | 1.00 | — | — | 1.99 | 1.00 | 0.96 | — | — | — | 0.69 | — | ||||||
| 钉柱委陵菜 Potentilla saundersiana | 1.00 | — | 5.13 | 3.00 | 5.36 | 5.44 | — | — | 1.70 | 1.10 | 1.73 | 1.74 | ||||||
| 高山唐松草 Thalictrum alpinum | — | — | 3.90 | 5.22 | — | 1.00 | — | — | 1.46 | 1.53 | — | — | ||||||
| 棱子芹 Pleurospermum camtschaticum | 1.00 | 1.00 | 1.00 | 1.00 | 2.89 | 2.19 | — | — | — | — | 1.08 | 0.92 | ||||||
| 马先蒿 Pedicularis reaupinanta | 1.00 | — | — | — | 3.80 | 4.22 | — | — | — | — | 1.36 | 1.52 | ||||||
| 大戟 Euphorbia pekinensis | — | 1.00 | 4.69 | 2.17 | 3.34 | — | — | — | 1.58 | 0.91 | 1.28 | — | ||||||
| 灰毛蓝钟花 Cyananthus incanus | 2.68 | — | 5.07 | 4.07 | — | — | 1.04 | — | 1.70 | 1.47 | — | — | ||||||
| “—”表示该物种在对照或增温处理下的该植物群落中无分布 | ||||||||||||||||||
使用两种生态位指数计算发现, 在高寒草原中, 对照处理下生态位宽度较大的物种(生态位宽度值前三位的物种)为高山嵩草、黑褐薹草、粗壮嵩草; 增温处理下生态位宽度较大的物种为西藏三毛草、西藏薹草、粗壮嵩草。在高寒草甸中, 对照处理下生态位宽度较大的物种为高山嵩草、粗壮嵩草、钉柱委陵菜; 增温处理下生态位宽度较大的物种为高山嵩草、高山唐松草、灰毛蓝钟花。在流石滩中, 对照处理下生态位宽度较大的物种为钉柱委陵菜、山居雪灵芝、紫花针茅; 增温处理下生态位宽度较大的物种为钉柱委陵菜、山居雪灵芝、马先蒿(表 3)。以上排序中, 高寒草原和高寒草甸的莎草类和禾本类植物居多, 流石滩中的垫状植物和杂草类植物居多, 增温后这些功能群的生态位宽度仍较大, 能够很好地利用资源, 增温对其造成的影响不大。
未增温情况下高寒草甸的群落稳定性最大, 增温后流石滩的群落稳定性最大(图 4)。增温后高寒草甸和流石滩的群落稳定性分别降低了66.8%和10.1%, 而高寒草原的群落稳定性增加了22.2%。研究结果表明, 高寒草甸较高寒草原和流石滩植物群落更易受到增温的影响。

|
| 图 4 增温对群落稳定性指数的影响 Fig. 4 Effects of warming on community stability index |
前人研究表明, 增温会改变典型高山植物群落的优势物种、物种丰富度和组成, 进而改变其群落结构[29]。本研究结果表明, 短期增温造成三种典型植物群落中的物种组成发生变化, 其中高寒草原的物种数量由20种减少为16种, 高寒草甸的物种数量由18种增加到22种, 流石滩的物种数量由13种增加到15种。这主要是增温造成了土壤水分的降低, 进而影响高山植物的生存状况[30]; 且这些变化的物种主要为伴生种或偶见种, 他们在群落中的数量较少, 所占比例不大, 对群落物种组成的影响较小。
低温是高山植物生长的主要限制因子, 模拟增温缓解了低温胁迫对高山植物生长发育的影响[11, 31], 进而影响其群落结构。本研究中, 莎草类植物高山嵩草为三种典型高山植物群落的优势物种(表 1)。由于高山嵩草根系分布较浅, 在温度升高导致的水分减少将可能抑制其生长[32—33]。因此, 在水分受限的高寒草原群落中, 增温后高山嵩草的优势度降低。但在高寒草甸群落中, 由于其土壤水分较为充足, 所以增温后高山嵩草的重要值减少不明显。在更高海拔的流石滩群落中, 低温是限制植物生长的最主要因素, 因此增温后高山嵩草的优势度上升。一些学者认为, 在植物群落中存在一些对温度变化极其敏感的物种, 增温对其的影响会改变植物的种间关系, 进而影响群落优势物种和组成[32], 如本研究中, 增温后高寒草甸群落中杂类草火绒草的重要值从0.01增加至0.18(表 1), 由偶见种演变为优势种, 导致其群落结构发生改变。
3.2 增温对不同植物群落物种多样性和生物量的影响目前学术界主要采用OTC增温模拟气候变暖对不同植物群落物种多样性的影响, 且发现增温对植物群落物种多样性有正效应、负效应和不显著影响等三类研究结果[34—37]。本研究中, 短期增温对三种典型高山植物群落的多样性没有显著影响, 这与马丹丹等[38]、马丽等[37]和吴红宝等[36]的研究结果吻合。这主要是由于植物群落中的物种组成变化是一个长时间的群落演替过程[39]。一方面, 由于本增温实验持续年限较短, 导致高山植物群落处于短期波动状态, 还未呈现出明显的多样性变化; 另一方面, 随着增温时间的延长, 由于样地围封及OTC增温装置的阻隔, 会限制风力和动物对种子的传播, 并减少群落演替过程中的新物种出现, 从而导致物种多样性可能出现变化。
低温胁迫是高海拔地区高寒草甸植物生长的主要限制因子[11—28]。增温可以缓解低温对于植物生长的胁迫, 为其提供更适宜的生长环境, 从而增加了地下生物量。同时, 也有研究表明受干旱胁迫影响时, 部分植物的根系生物量会增长[40]。本研究中, 利用OTC装置的增温处理下高寒草甸中植物地下生物量大幅增加。魏春雪[41]等的研究也表明, 增温增加了植物的根系生物量, 这与本研究中高寒草原和高寒草甸群落的根系生物量对增温的响应一致。流石滩和高寒草原群落中的植物受干旱胁迫的影响, 增温处理后其地下生物量无显著变化。王琪等[42]的研究表明, 增温会加重植物受到的干旱胁迫, 从而减少植物的地上生物量。本研究中, 增温后三种典型高山植物群落的地上生物量均呈降低趋势。此外, 增温也会通过影响土壤养分含量, 从而影响植物对养分的吸收, 进而间接影响植物的生长发育[43—44]。
3.3 增温对种群生态位和群落稳定性的影响研究发现, 在高山植物群落中优势物种的生态位宽度通常较大, 这也与李一凡等[45]的结果一致。这是因为物种生态位宽度越大, 其对环境资源的支配力越大, 适应能力越强, 往往能竞争成为植物群落中的优势种。另外, 增温后群落中各物种整体的生态位呈现下降趋势。由于短期增温后引起群落的波动, 并处于波动的“谷底”, 因而对资源的利用率未达到最高, 导致生态位有不同程度的下降。在高寒草原和高寒草甸植物群落中, 莎草类和禾本类在增温后仍保持较大的生态位宽度, 草地并未出现退化现象[46]; 在流石滩植物群落中, 垫状植物和杂草类植物在增温后仍保持较大的生态位宽度。这从另一方面也佐证了前述关于三种植物群落物种多样性变化不显著的结论。
本研究中, 增温改变了群落的稳定性, 造成高寒草甸和流石滩群落的稳定性降低, 而高寒草原群落的稳定性升高。这种现象被很多学者认为是物种多样性的丧失会导致群落稳定性的降低[47—48]。但在本研究中, 短期增温后高寒草甸的物种多样性并未表现出显著的变化趋势, 与前人的一些研究结果不同。另外, 高寒草甸的优势物种在未增温前仅有粗壮嵩草、高山嵩草、灰毛蓝钟花等3个物种, 在增温后演变为高山嵩草1个物种(表 1), 且上述3个优势物种生态位均降低(表 3), 群落中的优势物种发生明显变化, 从而导致高寒草甸植物群落的稳定性大幅下降[49]。
4 结论短期增温对西藏念青唐古拉山典型植物群落结构和稳定性影响的研究结果表明, 增温对群落物种组成影响较小, 对群落结构的影响主要表现在优势物种的改变, 对群落的物种多样性无影响。增温会降低三种典型高山植物群落的地上生物量, 显著降低高寒草原和高寒草甸的地上生物量, 显著增加高寒草甸的地下生物量, 进而导致高寒草甸地上地下生物量分配策略的改变。增温会造成群落中部分物种生态位宽度的下降, 同时增温后高寒草甸的稳定性大幅下降。总体上, 在全球气候变暖的趋势下, 青藏高原区高寒草甸带的植物群落更易受到影响。
| 物种 Species |
高寒草原 Alpine steppe |
高寒草甸 Alpine meadow |
流石滩 Screes |
|||||
| 对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
对照 Control |
增温 Warming |
|||
| 青藏薹草 Carex moorcroftii | — | 10.06 | — | 9.34 | — | — | ||
| 粗壮嵩草 Carex sargentiana | 5.77 | 16.73 | 11.16 | 7.56 | — | — | ||
| 黑褐薹草 Carex atrofusca | 9.15 | 2.03 | 6.08 | 3.88 | — | — | ||
| 高山嵩草 Carex parvula | 26.67 | 3.31 | 22.91 | 26.26 | 10.14 | 20.95 | ||
| 矮生嵩草 Carex alatauensis | — | — | — | 3.56 | — | — | ||
| 西藏三毛草 Trisetum tibeticum | — | 6.63 | — | — | — | — | ||
| 丝颖针茅 Stipa capillacea | 11.45 | 13.75 | 3.36 | 6.90 | 3.60 | 3.89 | ||
| 紫花针茅 Stipa purpurea | 4.67 | 5.02 | — | 1.22 | 16.52 | 5.97 | ||
| 早熟禾 Poa annua | — | — | — | — | — | — | ||
| 冰川棘豆 Oxytropis glacialis | 1.98 | — | 0.90 | 0.65 | — | — | ||
| 高山棘豆 Oxytropis alpina | — | — | 0.89 | — | — | — | ||
| 高山豆 Tibetia himalaica | — | — | — | 0.48 | — | — | ||
| 垫状点地梅 Androsace tapete | — | — | 5.79 | 0.22 | 4.52 | 4.94 | ||
| 山居雪灵芝 Arenaria edgeworthiana | 0.27 | — | — | — | 21.40 | 8.23 | ||
| 二裂委陵菜 Potentilla bifurca | 0.44 | 2.37 | 0.57 | — | — | — | ||
| 蕨麻 Argentina anserina | — | 3.70 | 2.05 | 3.05 | — | — | ||
| 乳白香青 Anaphalis lactea | 8.10 | 3.23 | — | — | 1.69 | 1.37 | ||
| 钉柱委陵菜 Potentilla saundersiana | 0.70 | — | 6.93 | 4.38 | 11.46 | 13.17 | ||
| 西藏风毛菊 Saussurea tibetica | — | — | — | 2.93 | — | — | ||
| 冷蒿 Artemisia frigida | — | — | — | — | — | — | ||
| 藏沙蒿 Artemisia wellbyi | — | — | — | — | — | — | ||
| 西藏报春 Primula tibetica | — | — | 1.64 | 0.11 | — | — | ||
| 高山唐松草 Thalictrum alpinum | — | — | 7.53 | 8.72 | — | 1.41 | ||
| 云雾龙胆 Gentiana nubigena | — | — | — | — | — | — | ||
| 线叶龙胆 Gentiana farreri | — | — | — | — | — | — | ||
| 棱子芹 Pleurospermum camtschaticum | 1.67 | 1.26 | 0.45 | 0.30 | 3.56 | 4.51 | ||
| 龙胆 Gentiana scabra Bunge | 0.54 | — | 0.59 | — | — | — | ||
| 肉果草 Lancea tibetica | — | — | — | 0.47 | — | — | ||
| 小叶金露梅 Dasiphora parvifolia | — | — | — | — | 8.03 | 19.22 | ||
| 蚤缀 Arenaria serpyllifolia | — | — | — | — | 4.85 | 6.05 | ||
| 马先蒿 Pedicularis resupinata | 0.61 | — | — | — | 3.32 | 4.38 | ||
| 未知杂草 | — | — | 7.86 | 5.48 | — | — | ||
| 狼毒 Stellera chamaejasme | 4.46 | 5.20 | — | 1.11 | — | — | ||
| 微孔草 Microula sikkimensis | 5.61 | 4.06 | — | — | — | — | ||
| 火绒草 Leontopodium leontopodioides | 0.84 | 17.78 | — | — | — | — | ||
| 大戟 Euphorbia pekinensis | — | 3.44 | 1.25 | 2.10 | 4.98 | — | ||
| 鸦跖花 Oxygraphis glacialis | — | — | 0.49 | 0.66 | — | — | ||
| 密花毛果草 Lasiocaryum densifiorum | — | — | — | — | — | — | ||
| 藏玄参 Oreosolen wattii | — | — | — | — | — | — | ||
| 灰毛蓝钟花 Cyananthus incanus | 2.26 | — | 12.48 | 8.37 | — | — | ||
| 独一味 Phlomoides rotata | — | — | — | — | — | — | ||
| 三裂碱毛茛 Halerpestes tricuspis | — | — | 1.90 | 2.23 | — | — | ||
| 西藏蒲公英 Taraxacum tibetanum | — | — | — | — | — | — | ||
| 平卧轴藜 Axyris prostrata | — | — | — | — | — | — | ||
| 圆穗蓼 Bistorta macrophylla | — | — | — | — | — | — | ||
| 葶苈 Draba nemorosa | — | — | 4.61 | — | — | — | ||
| 红景天 Rhodiola rosea | — | — | — | — | 5.92 | 2.73 | ||
| 圆萼刺参 Morina chinensis | 10.48 | 1.43 | — | — | — | — | ||
| 蓝翠雀花 Delphinium caeruleum | 0.54 | — | — | — | — | — | ||
| 珠芽蓼 Bistorta vivipara | — | — | 0.57 | — | — | — | ||
| 喉毛花 Comastoma pulmonarium | 3.79 | — | — | — | — | — | ||
| 锦鸡儿 Caragana sinica | — | — | — | — | — | — | ||
| 冰岛蓼 Koenigia islandica | — | — | — | — | — | 3.16 | ||
| “—”表示该物种在对照或增温处理下的该植物群落中无分布 | ||||||||
| [1] |
周华坤, 周立, 赵新全, 刘伟, 李英年, 古松, 周兴民. 青藏高原高寒草甸生态系统稳定性研究. 科学通报, 2006, 51(1): 63-69. |
| [2] |
Zhao J X, Luo T X, Li R C, Li X, Tian L H. Grazing effect on growing season ecosystem respiration and its temperature sensitivity in alpine grasslands along a large altitudinal gradient on the central Tibetan Plateau. Agricultural and Forest Meteorology, 2016, 218/219: 114-121. DOI:10.1016/j.agrformet.2015.12.005 |
| [3] |
Zhao J X, Sun F D, Tian L H. Altitudinal pattern of grazing exclusion effects on vegetation characteristics and soil properties in alpine grasslands on the central Tibetan Plateau. Journal of Soils and Sediments, 2019, 19(2): 750-761. DOI:10.1007/s11368-018-2056-6 |
| [4] |
Rustad L, Campbell J, Marion G, Norby R, Mitchell M, Hartley A, Cornelissen J, Gurevitch J, GCTE-NEWS. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia, 2001, 126(4): 543-562. DOI:10.1007/s004420000544 |
| [5] |
Jonasson S, Michelsen A, Schmidt I K, Nielsen E V. Responses in microbes and plants to changed temperature, nutrient, and light regimes in the Arctic. Ecology, 1999, 80(6): 1828-1843. DOI:10.1890/0012-9658(1999)080[1828:RIMAPT]2.0.CO;2 |
| [6] |
Klanderud K, Totland Ø. Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology, 2005, 86(8): 2047-2054. DOI:10.1890/04-1563 |
| [7] |
侯颖, 王开运, 张超. 大气二氧化碳浓度与温度升高对红桦幼苗养分积累和分配的影响. 应用生态学报, 2008, 19(1): 13-19. DOI:10.13287/j.1001-9332.2008.0001 |
| [8] |
Arft A M, Walker M D, Gurevitch J, Alatalo J M, Bret-Harte M S, Dale M, Diemer M, Gugerli F, Henry G H R, Jones M H, Hollister R D, Jónsdóttir I S, Laine K, Lévesque E, Marion G M, Molau U, Mølgaard P, Nordenhäll U, Raszhivin V, Robinson C H, Starr G, Stenström A, Stenström M, Totland Ø, Turner P L, Walker L J, Webber P J, Welker J M, Wookey P A. Responses of tundra plants to experimental warming: meta-analysis of the international tundra experiment. Ecological Monographs, 1999, 69(4): 491-511. |
| [9] |
Henry G H R, Molau U. Tundra plants and climate change: the International Tundra Experiment (ITEX). Global Change Biology, 1997, 3(S1): 1-9. DOI:10.1111/j.1365-2486.1997.gcb132.x |
| [10] |
Couteaux M M, Bottner P, Berg B. Litter decomposition, climate and liter quality. Trends in Ecology & Evolution, 1995, 10(2): 63-66. |
| [11] |
李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895-905. |
| [12] |
张继义, 赵哈林, 张铜会, 赵学勇. 科尔沁沙地植物群落恢复演替系列种群生态位动态特征. 生态学报, 2003, 23(12): 2741-2746. |
| [13] |
刘加珍, 陈亚宁, 张元明. 塔里木河中游主要植物种群的生态特征分析. 地理研究, 2003, 22(5): 663-670. |
| [14] |
Weigelt A, Schumacher J, Roscher C, Schmid B. Does biodiversity increase spatial stability in plant community biomass?. Ecology Letters, 2008, 11(4): 338-347. DOI:10.1111/j.1461-0248.2007.01145.x |
| [15] |
Chalcraft D R. Changes in ecological stability across realistic biodiversity gradients depend on spatial scale. Global Ecology and Biogeography, 2013, 22(1): 19-28. DOI:10.1111/j.1466-8238.2012.00779.x |
| [16] |
Li R, Luo T, Tang Y, Du M, Zhang X. The altitudinal distribution center of a widespread cushion species is related to an optimum combination of temperature and precipitation in the central Tibetan Plateau. Journal of Arid Environments, 2013, 88: 70-77. DOI:10.1016/j.jaridenv.2012.07.018 |
| [17] |
Wang Z, Luo T X, Li R C, Tang Y H, Du M Y. Causes for the unimodal pattern of biomass and productivity in alpine grasslands along a large altitudinal gradient in semi-arid regions. Journal of Vegetation Science, 2013, 24(1): 189-201. DOI:10.1111/j.1654-1103.2012.01442.x |
| [18] |
Guo G X, Kong W D, Liu J B, Zhao J X, Du H D, Zhang X Z, Xia P H. Diversity and distribution of autotrophic microbial community along environmental gradients in grassland soils on the Tibetan Plateau. Applied Microbiology and Biotechnology, 2015, 99(20): 8765-8776. DOI:10.1007/s00253-015-6723-x |
| [19] |
赵艳艳, 周华坤, 姚步青, 王文颖, 董世魁, 赵新全. 长期增温对高寒草甸植物群落和土壤养分的影响. 草地学报, 2015, 23(4): 665-671. |
| [20] |
马志良. 青藏高原东缘高寒灌丛根系分泌物碳氮输入对增温的响应. 生态环境学报, 2020, 29(4): 643-649. |
| [21] |
蔡瑛莹. 土壤增温和氮沉降对杉木幼树细根分泌物通量和化学组成的影响[D]. 福州: 福建师范大学, 2018: 35-38.
|
| [22] |
王丽霞, 郭宏宇, 霍玉珠, 庞金玲, 王欢欢, 马成仓, 王银华. 增温和增氮对天津滨海湿地芦苇凋落物分解微生物群落组成和多样性的影响. 天津师范大学学报: 自然科学版, 2022, 42(1): 37-44. |
| [23] |
姚世庭, 芦光新, 邓晔, 范月君, 周华坤, 党宁, 王英成, 张海娟, 颜珲璘. 模拟增温对高寒草地土壤原核生物群落组成及多样性影响. 草地学报, 2021, 29(S1): 27-34. |
| [24] |
Körner C. The use of 'altitude' in ecological research. Trends in Ecology & Evolution, 2007, 22(11): 569-574. |
| [25] |
Fu G, Shen Z X, Zhang X Z, Zhou Y T. Response of soil microbial biomass to short-term experimental warming in alpine meadow on the Tibetan Plateau. Applied Soil Ecology, 2012, 61: 158-160. DOI:10.1016/j.apsoil.2012.05.002 |
| [26] |
张立, 次仁曲珍, 洛桑催成, 扎西, 德庆卓嘎, 普布次仁, 平措班旦, 格桑加措, 巴桑吉巴, 央金, 李宝海. 西藏当雄县高寒草甸及河滩草甸牧草营养价值分析. 中国畜牧杂志, 2018, 54(2): 73-75. |
| [27] |
李翔, 王忠, 赵景学, 罗天祥. 念青唐古拉山南坡高寒草甸生产力对温度和降水变化的敏感性及其海拔分异. 生态学报, 2017, 37(17): 5591-5601. |
| [28] |
王忠. 念青唐古拉山南坡高寒草甸生产力沿海拔梯度的变化机理研究[D]. 北京: 中国科学院大学, 2011.
|
| [29] |
赵成章, 樊胜岳, 殷翠琴, 贺学斌. 毒杂草型退化草地植被群落特征的研究. 中国沙漠, 2004, 24(4): 507-512. |
| [30] |
王志鹏, 张宪洲, 何永涛, 石培礼, 俎佳星, 牛犇, 李猛. 降水变化对藏北高寒草原化草甸降水利用效率及地上生产力的影响. 应用生态学报, 2018, 29(6): 1822-1828. |
| [31] |
王晨晨, 王珍, 张新杰, 李寅龙, 刘军利, 韩国栋. 增温对荒漠草原植物群落组成及物种多样性的影响. 生态环境学报, 2014, 23(1): 43-49. |
| [32] |
Dorji T, Totland Ø, Moe S R, Hopping K A, Pan J B, Klein J A. Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet. Global Change Biology, 2013, 19(2): 459-472. |
| [33] |
Dorji T, Hopping K A, Wang S P, Piao S L, Tarchen T, Klein J A. Grazing and spring snow counteract the effects of warming on an alpine plant community in Tibet through effects on the dominant species. Agricultural and Forest Meteorology, 2018, 263: 188-197. |
| [34] |
Zhang Y Q, Welker J M. Tibetan alpine tundra responses to simulated changes in climate: aboveground biomass and community responses. Arctic and Alpine Research, 1996, 28(2): 203. |
| [35] |
Ganjurjav H, Gao Q Z, Gornish E S, Schwartz M W, Liang Y, Cao X J, Zhang W N, Zhang Y, Li W H, Wan Y F, Li Y, Danjiu L B, Guo H B, Lin E D. Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai-Tibetan Plateau. Agricultural and Forest Meteorology, 2016, 223: 233-240. |
| [36] |
吴红宝, 高清竹, 干珠扎布, 李钰, 闫玉龙, 胡国铮, 王学霞, 严俊, 何世丞. 放牧和模拟增温对藏北高寒草地植物群落特征及生产力的影响. 植物生态学报, 2019, 43(10): 853-862. |
| [37] |
马丽, 张骞, 张中华, 郭婧, 杨晓渊, 周秉荣, 邓艳芳, 王芳, 佘延娣, 周华坤. 梯度增温对高寒草甸物种多样性和生物量的影响. 草地学报, 2020, 28(5): 1395-1402. |
| [38] |
马丹丹, 王瑾瑜. 长期增温对荒漠草原短花针茅群落组成和土壤养分及酶活性特征的影响. 甘肃农业大学学报, 2020, 55(3): 144-153. |
| [39] |
Zhang C H, Willis C G, Klein J A, Ma Z, Li J Y, Zhou H K, Zhao X Q. Recovery of plant species diversity during long-term experimental warming of a species-rich alpine meadow community on the Qinghai-Tibet Plateau. Biological Conservation, 2017, 213: 218-224. |
| [40] |
董义凤, 霍光伟, 李平芳, 立青次木, 高晓艺. 干旱胁迫对克氏针茅和多根葱生理生态指标的影响分析. 绿色科技, 2022, 24(1): 26-28. |
| [41] |
魏春雪, 杨璐, 汪金松, 杨家明, 史嘉炜, 田大栓, 周青平, 牛书丽. 实验增温对陆地生态系统根系生物量的影响. 植物生态学报, 2021, 45(11): 1203-1212. |
| [42] |
王琪, 郑佳华, 赵萌莉, 张军. 增温对荒漠草原不同退化程度草地恢复初期影响的研究. 草地学报, 2022, 30(5): 1077-1085. |
| [43] |
Alward R D, Detling J K, Milchunas D G. Grassland vegetation changes and nocturnal global warming. Science, 1999, 283(5399): 229-231. |
| [44] |
Eatherall A. Modelling climate change impacts on ecosystems using linked models and a GIS. Climatic Change, 1997, 35(1): 17-34. |
| [45] |
李一凡, 刘梦芸, 甘先华, 黄芳芳, 张卫强. 深圳市坝光湿地园银叶树群落优势种生态位特征. 生态环境学报, 2020, 29(11): 2171-2178. |
| [46] |
刘学敏, 罗久富, 陈德朝, 朱欣伟, 周金星. 若尔盖高原不同退化程度草地植物种群生态位特征. 浙江农林大学学报, 2019, 36(2): 289-297. |
| [47] |
Zhang Y H, Feng J C, Loreau M, He N P, Han X G, Jiang L. Nitrogen addition does not reduce the role of spatial asynchrony in stabilising grassland communities. Ecology Letters, 2019, 22(4): 563-571. |
| [48] |
薛亚芳, 宗宁, 何念鹏, 田静, 张永清. 长期围封和自由放牧对高寒草甸土壤微生物群落结构及碳源代谢多样性的影响. 应用生态学报, 2018, 29(8): 2705-2712. |
| [49] |
Wu Q, Ren H Y, Wang Z W, Li Z G, Liu Y H, Wang Z, Li Y H, Zhang R Y, Zhao M L, Chang S X, Han G D. Additive negative effects of decadal warming and nitrogen addition on grassland community stability. Journal of Ecology, 2020, 108(4): 1442-1452. |