 2022, Vol. 42
2022, Vol. 42文章信息
- 简春霞, 赖帅彬, 周俊杰, 陈志飞, 杨全, 陈阳, 徐炳成
- JIAN Chunxia, LAI Shuaibin, ZHOU Junjie, CHEN Zhifei, YANG Quan, CHEN Yang, XU Bingcheng
- 黄土丘陵区典型小流域草地群落数量分类与结构特征及其影响因素
- Quantitative and structure characteristics and the influencing factors of grassland communities in a typical watershed at loess hilly-gully region
- 生态学报. 2022, 42(4): 1381-1392
- Acta Ecologica Sinica. 2022, 42(4): 1381-1392
- http://dx.doi.org/10.5846/stxb202010272753
-
文章历史
- 收稿日期: 2020-10-27
- 网络出版日期: 2021-11-04

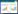




2. 中国科学院水利部水土保持研究所, 杨凌 712100
2. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100, China
黄土高原退耕还林(草)工程实施20年来, 草地逐渐成为生态系统的主体[1], 及时了解草地群落类型、结构特征及其影响因素具有重要的现实意义。黄土高原草地恢复相关研究结果表明, 退耕草地演替在时间序列上大致经历了猪毛蒿、达乌里胡枝子和长芒草、铁杆蒿以及白羊草群落等类型, 群落盖度、物种多样性和生物量等随演替时间延长均会上升[2—5]。上述研究均以撂荒时间为主线, 在实际调查中很难获取演替的准确年代信息, 同时由于该区立地条件复杂多变, 生境异质性导致草地群落构建与演替不可能遵循相同规律, 物种对生境的适应性差异可能促使草地恢复方向与进程产生变化。因此, 在区域内均匀选择样点对准确掌握草地恢复状况、群落结构与生境条件关系更具合理性。
物种组成是群落构建的基本要素, 植被盖度、生产力和物种多样性均为衡量群落结构及功能的重要指标, 物种属性及不同物种组合与群落结构间的关系一直是生态学研究的基本问题。植物分类可在一定程度上揭示植物群落类型与生境关系, 数量分类是植物分类学研究中的重要方法[6]。排序方法能够将物种数据尽可能展示在可视的二维空间, 排序图反映各样方最适生态梯度分布状况, 可了解群落演替方向、速度和干扰状况[7], 聚类方法与排序相结合, 易于更直观了解群落分布、演替规律以及物种与环境间的相关关系[8]。目前国内运用数量生态学方法对草地群落展开研究多集中在高山草甸[9—10]、内蒙古典型草原[11]、湿地草原[12]和西北荒漠草原[13]等, 对黄土高原自然恢复草地相对较少。白文娟等[14]运用TWINSPAN和CA排序方法对安塞不同恢复年限草地研究发现, 40年恢复进程草地可划分为4类群落, 土壤有机质A氮磷钾A土壤水分以及坡度与植物群落变化之间的关系密切。不同学者选择具体方法不尽相同, 但从深度来看, 仅停留在群落分类以及找到显著影响的环境因子层面[15], 缺乏对环境因子与群落类型分异及结构差异的作用方式的思考。
环境因素对草地恢复影响与尺度有关, 大尺度内气候因子占主导作用, 地形与土壤因子则在小尺度内更为关键[16]。沟壑纵横引起小尺度内生境也存在着较大差异, 小尺度的研究可为大尺度容易忽视的环境过滤效应提供更加详细的解释[17]。小流域是水土保持综合治理的基本单元, 基于小流域的研究可为区域尺度的研究提供全面、详细的基础资料[18]。研究表明, 小尺度条件下土壤养分对地下生物量解释度更高, 而植物性状对地上生量解释度更高[19]。张凯等[20]研究结果显示, 地上生物量和盖度主要受土壤有机碳、矿质氮、全氮、土壤含水量和海拔的影响, 物种多样性则更多地受土壤饱和导水率、容重、砂粒和粉粒含量的影响。海拔、土壤碳氮含量、土壤水分等对种群适应性、种间关系以及地上生物量均产生了重要作用[21—22]。生物因素在控制生态系统特性方面与非生物因素同样重要, 功能群组成对初级生产力和土壤养分利用率均产生影响[16, 23]。综合前人的研究, 环境因子对草地群落物种、生产力、多样性等方面的影响非常复杂, 过往研究常通过控制变量方法探讨某一类因子, 或将生物因子和非生物因子分开讨论, 而二者相互作用共同调控生态系统功能与稳定性[24]。因此, 本文对群落类型分异、结构及环境间关系进行系统的研究: 以黄土丘陵区纸坊沟小流域为研究区, 采用数量生态学与结构方程模型相结合方法, 旨在揭示: ①草地群落类型及其物种组成和群落结构差异;②判别引起草地群落分异的生物和环境因素。
1 材料与方法 1.1 研究区概况研究区为陕西省延安市安塞区纸坊沟小流域(109°13′46″—109°16′03″E, 36°42′42″—36°46′28″N), 位于黄土丘陵区沿河一级支流下游, 1973年开始水土保持综合治理, 在水土流失、植被恢复和农田生态系统方面具有良好且广泛的研究基础[13]。流域面积8.27 km2, 海拔1010—1430 m, 属暖温带半湿润气候向半干旱气候过渡区, 年均气温8.8℃。年均降水量为528.8 mm(1951—2016), 但季节分布不均, 5—9月降雨量约占全年82.2%, 无霜期约160 d。土壤类型为风积黄土母质发育而来的黄绵土。植被类型为暖温带落叶阔叶林区向暖温带草原区过渡的森林草原带, 以大量人工种植的刺槐(Robinia pseudoacacia)、小叶杨(Populus simonii)等人工林和退耕地自然恢复草地为主, 典型地带性草本植物有白羊草(Bothriochloa ischaemum)、达乌里胡枝子(Lespedeza davurica)、铁杆蒿(Artemisia sacrorum)、长芒草(Stipa bungeana)等。
1.2 研究方法2019年8月, 根据流域基本地貌特征, 以流域主沟道为纵向样带, 自沟头向下每隔大致1 km设置横向样带, 在两侧随机取样, 依次选择22个受人为干扰较小、地形较均匀且植物生长相似的草地样地(图 1)。记录各样地经纬度、海拔、坡度及坡向信息。对坡向进行数量化转化, 使用南方向为起始0°, 坡向角度余弦值代表坡向;坡度采用同一样地内3个样方坡度均值。各样地内随机设置3个1 m×1 m样方进行物种调查, 样方间距5—10 m。记录样方内出现的所有植物种类、株数、株高、植被盖度、各物种分种盖度。植被密度(PD, N/m 2)为样方内单位面积所有物种之和;群落盖度采用照相法测定[25], 分种盖度采用目估法;物种株高采用皮尺量取样方内每种植物的自然直立高度, 取3次平均。地上生物量(AGB)和地下生物量(BGB)均采用收获法测定。植物地上部分齐地刈割分种装袋, 置于75℃烘箱内烘48 h至恒重(0.01 g/m2), 获得AGB。各样方内对角线上平均取3个点, 用直径9 cm根钻取0—30 cm土层, 去除杂质冲洗干净后置于75℃烘箱内烘48 h至恒重(0.01 g/m2), 获得BGB。根冠比(R/S)为BGB和AGB的比值。

|
| 图 1 研究区地理位置及22个草地样地分布情况 Fig. 1 Geographical location and distribution of 22 grassland plots in the study region |
地上生物量收割后, 在各样方内对角线上平均取3个点, 用直径9 cm根钻取0—30 cm土层土壤样品, 混合均匀后带回实验室自然阴干。剔除土壤中的根系、石块及动植物残体, 将土壤研磨, 过2 mm土壤筛, 进行土壤pH测定。土壤pH采用国标法(NY/T 1377—2007), 使用雷磁E-201型复合电极测定(土水比2.5∶1)。风干土壤过0.15 mm土壤筛, 进行土壤有机碳、土壤全氮、土壤全磷含量测定。土壤有机碳(SOC, g/kg)测定采用重铬酸钾—外加热法(H2SO4-K2Cr2O7);全氮(TN, g/kg)采用连续流动分析仪(英国SEAL-AA3)测定;全磷(TP, g/kg)采用钼锑抗比色法测定[26]。每个土壤样品重复测定3次后取其平均值。
1.4 统计与分析 1.4.1 群落平均高度与功能群划分群落加权平均高度(WH)使用各物种株数的加权平均高度之和求得(公式1)。功能群是指群落中在形态、生活习性、结构和功能方面等方面中某特征相同的物种的集合, 在群落中占据着类似的生态位或发挥着相似的功能[16]。冠层高度在植物对光能及二氧化碳等资源的截取、吸收与利用等方面发挥重要作用, 可作为生态位特征对进行功能群划分[27]。本研究中22个样地群落平均高度为31 cm, 对83种植物高度进行加权平均, 植株平均高度 ≥ 31 cm为高草, < 31 cm为矮草。综上, 本研究将物种划分为禾本科高草(TG)、禾本科矮草(SG)、豆科高草(TL)、豆科矮草(SL)、菊科高草(TA)、菊科矮草(SA)和杂类草7类功能群[28]。
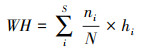
|
(1) |
式中, S为物种总数, ni为物种i的株数, N为总株数, hi为物种i平均高度。
1.4.2 物种多样性指数草地群落物种组成采用各物种重要值(IV)表征, 群落α多样性指数采用丰富度(R)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)和Simpson优势度指数(D)共同表征[29]。

|
(2) |
式中, 相对盖度=某个物种盖度/所有物种盖度之和;相对高度=某个物种高度/所有物种高度之和;相对生物量=某个物种生物量鲜重/所有物种生物量。
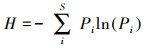
|
(3) |
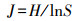
|
(4) |
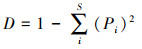
|
(5) |
式中, S为物种总数, Pi为某一物种重要值与所有物种重要值之和的比值。
1.5 数据分析使用Ward聚类法对草地群落进行分类, 结合非度量多维度排序(NMDS)法分析草地群落分布差异。通过NMDS与群落结构特征和环境因素拟合, 分析影响群落分布差异的环境因素[30]。使用指示种表明群落类型间物种组成差异, 单因素方差分析(ANOVA)说明结构特征和物种多样性差异。使用功能群重要值和群落结构特征与环境因素进行结构方程建模, 以探究环境因素影响群落结构特征的内在机理。
偶见种出现频率低, 但在个别样方中显示较高重要值, 属异常值, 对Bray-Curtis距离计算产生影响, 因此剔除在所有样方中出现频率低于5%的物种, 共选取40种常见种进行聚类和排序分析。以行代表样方, 列代表物种, 建立一个63×40的“样方—物种”原始矩阵, 计算样方间Bray-Curtis距离, 基于Bray-Curtis距离进行Ward聚类。Mantel分析代表不同分类水平的二元矩阵间相关性, 本研究选取相关系数最高的分类水平作为最优分组方案。ANOSIM相似性分析(Analysis of Similarities)对聚类结果进行组间差异性非参数检验, R范围[-1, 1], R > 0组间差异大于组内, R < 0组间差异小于组内。使用Permutation置换检验说明组间差异是否显著, 本研究中ANOSIM分析R值为0.524, 大于零模型(quantiles of permutations)99%分位数0.131, 分类结果在P < 0.001水平显著。
参照Wiesmair等[25]的研究方法, 使用vegan包中的metaMDS函数对样地进行NMDS分析。共选择7个环境因子(海拔Altitude、坡度Slope、土壤pH值、土壤有机碳SOC、全氮TN和全磷TP), 6个群落主要特征(植物密度PD、植被盖度Coverag、群落加权平均高度WH、地上生物量AGB、地下生物量BGB和根冠比R/S), 以及4个物种多样性参数(丰富度R、Shannon-Wiener多样性指数H、Pielou均匀度指数J、Simpson优势度指数D)作为响应变量, 样方物种排序得分作为自变量, 使用vegan包中的envfit函数作多元回归分析, 以确定草地群落的类型分布与潜在环境因子间的关系。使用置换的方法检验不同分组间的统计学显著性(P < 0.05), 仅当变量的组间差异显著时回归结果有效(P < 0.05)。使用应力函数stress衡量排序拟合度, Stress < 0.05, 表明拟合极好;Stress < 0.1, 拟合较好;Stress < 0.2, 拟合可接受。Stress > 0.2, 表明拟合较差, 解释力较弱。本研究中, NMDS排序应力函数值为0.12, 拟合结果可接受。指示种分析在labdsv包中indval函数完成, 指示值计算方法为植物的频率×重要值, 并用置换的方法检验不同分组间的统计学显著性(P < 0.05)。
结构方程模型(SEM)适合评估变量间的关系, 常被用于研究因素对观测变量的影响。用最低Akaike information准则(AIC)去除不显著变量, 使用卡方检验(χ, P > 0.05)和近似的均方根误差(RMSEA, P < 0.05)评估模型拟合度。
聚类分析(Ward)、排序(NMDS)以及指示种分析均在R 4.1中完成。单因素方差分析(ANOVA)在SPSS 20.0中完成。SEM分析在Amos21.0中完成。相关绘图在Origin 9.0中完成。
2 结果分析 2.1 草地群落类型22个样点中共发现83种植物, 一年生草本21种, 多年生草本54种, 灌木或乔木8种;83个物种分属26科65属, 其中菊科23种, 禾本科11种, 豆科10种(表 1和3)。对40种常见种在63个样方中的重要值进行Ward聚类, 结合Mantel分析, 在第3级聚类簇上裁剪时分类效果最优(图 2), 最优分类数为5类。使用ANOSIM差异性分析对分类结果进行检验, 组间差异极显著(r2 = 0.73, P = 0.001)。
| 群丛类型 Association type |
指示种 Indicator species |
重要值 Importance value |
指示值 Indicator values |
群丛类型 Association type |
指示种 Indicator species |
重要值 Importance value |
指示值 Indicator values |
|
| Ⅰ.狗尾草群丛 | 狗尾草 | 10.1 | 78*** | Ⅲ.甘草群丛 | 甘草 | 18.4 | 33** | |
| Setaria viridis ass | 鹅冠草 | 7.8 | 29* | Glycyrrhiza uralensis ass | 草木樨状黄耆 | 17.0 | 26* | |
| 猪毛菜 | 7.6 | 31*** | 败酱 | 8.0 | 37* | |||
| 鬼针草 | 4.9 | 62*** | 小红菊 | 5.1 | 33* | |||
| 阿尔泰狗娃花 | 4.1 | 32* | 丛生隐子草 | 3.4 | 40** | |||
| 风毛菊 | 3.4 | 62*** | Ⅳ. 铁杆蒿群丛 Artemisia sacrorum ass |
铁杆蒿 | 16.8 | 47*** | ||
| 老鹳草 | 3.1 | 31* | Ⅴ.白羊草群丛 | 白羊草 | 20.6 | 50*** | ||
| 地锦 | 1.8 | 56*** | Bothriochloa ischaemum ass | 砂珍棘豆 | 2.8 | 30* | ||
| 苦荬菜 | 1.5 | 90*** | 西伯利亚远志 | 2.7 | 33* | |||
| Ⅱ.茵陈蒿群丛 | 茵陈蒿 | 8.6 | 53*** | |||||
| Artemisia capillaris ass | 硬质早熟禾 | 6.0 | 31* | |||||
| 香青兰 | 2.9 | 33* | ||||||
| * 代表物种指示值在不同群丛间差异显著(P < 0.05) | ||||||||
| 指标 Index |
狗尾草群丛 Setaria viridis ass |
茵陈蒿群丛 Artemisia capillaris ass |
甘草群丛 Glycyrrhiza uralensis ass |
铁杆蒿群丛 Artemisia sacrorum ass |
白羊草群丛 Bothriochloa ischaemum ass |
|
| 环境因子 | Altitude/m | 1237 | 1174 | 11155 | 1211 | 1162 |
| Environment factors | Slop/(°) | 22 | 15 | 15 | 13 | 22 |
| SOC/(g/kg) | 4.04 ± 0.79b | 3.22 ± 0.28b | 6.38 ± 2.51a | 6.25 ± 2.53a | 7.86 ± 4.01a | |
| TN/(g/kg) | 0.58 ± 0.04c | 0.64 ± 0.14c | 0.85 ± 0.24ab | 0.73 ± 0.28bc | 1.05 ± 0.36a | |
| TP/(g/kg) | 0.57 ± 0.03a | 0.54 ± 0.04a | 0.55 ± 0.03a | 0.49 ± 0.10a | 0.53 ± 0.05a | |
| pH | 8.50 ± 0.25a | 8.70 ± 0.37a | 8.56 ± 0.15a | 8.40 ± 0.14a | 8.50 ± 0.26a | |
| 群落结构 | PD/(N/m2) | 205 ± 107ab | 145 ± 62bc | 255 ± 74a | 261 ± 127a | 241 ± 112a |
| Community structure | WH/cm | 35.9 ± 15.2a | 37.5 ± 20.1a | 35.8 ± 13.1a | 32.1 ± 12.9a | 25.5 ± 8.3a |
| Coverage/% | 55.1 ± 10.8b | 61.6 ± 17.7ab | 68.8 ± 19.2ab | 74.5 ± 9.9a | 72.7 ± 18.2a | |
| AGB/(g/m2) | 112.6 ± 32.0b | 239.6 ± 62.2a | 197.9 ± 82.6a | 204.3 ± 65.4a | 194.0 ± 59.1a | |
| BGB/(g/m2) | 127.6 ± 52.4b | 392.7 ± 178.98a | 281.7 ± 164.4ab | 395.8 ± 236.5a | 403.8 ± 160.5a | |
| R/S | 1.1 ± 0.3b | 1.7 ± 0.8ab | 1.5 ± 1.1ab | 2.2 ± 1.4ab | 2.4 ± 1.7a | |
| 多样性指数 | R | 15 ± 1a | 12 ± 3b | 13 ± 3a | 13 ± 3a | 14 ± 3a |
| Diversity index | H | 2.36 ± 0.19a | 1.97 ± 0.32b | 2.17 ± 0.23ab | 2.00 ± 0.33b | 2.07 ± 0.31b |
| J | 0.87 ± 0.05a | 0.80 ± 0.05b | 0.84 ± 0.04ab | 0.80 ± 0.07b | 0.81 ± 0.08b | |
| D | 0.76 ± 0.21a | 0.59 ± 0.22b | 0.84 ± 0.04a | 0.80 ± 0.07a | 0.77 ± 0.15a | |
| 平均值±标准差Mean ± SD; 同行数字后不同小写字母表示群丛间差异显著(P < 0.05);Altitude: 海拔; Slop: 坡度; SOC: Soil organic carbon土壤有机碳; TN: Total nitrogen全氮; TP: Total phosphorus全磷; pH: 土壤pH值; PD: plant diversity植物密度; CH: Community Height群落高度; Coverage: 群落盖度; AGB: aboveground biomass地上生物量; BGB: belowground biomass地下生物量; R/S: Root/Shhoot根冠比; R: Richness丰富度; H: Shannon-Wiener index香农多样性指数; J: Pielou's evenness index均匀度指数; D: Simpson's diversity index优势度指数 | ||||||
| 物种 Species |
重要值 Important value |
物种 Species |
重要值 Important value |
|
| 长芒草Stipa bungeana | 12.9 | 地梢瓜Cynanchum thesioides | 2.8 | |
| 达乌里胡枝子Lespedeza davurica | 11.2 | 独行菜Lepidium apetalum | 1.5 | |
| 紫花地丁Viola philippica | 1.6 | 活血丹Glechoma longituba | 1.3 | |
| 蒙古蒿Artemisia mongolica | 5.8 | 角蒿Incarvillea sinensis | 2.7 | |
| 菊叶委陵菜Potentilla tanacetifolia | 5.8 | 节节草Equisetum ramosissimum | 4.4 | |
| 细叶远志Polygala linarifolia | 2.5 | 苦苣Sonchus oleraceus | 3.8 | |
| 阴行草Siphonostegia chinensis | 10.2 | 狼牙刺Sophora davidii | 2.0 | |
| 华北米蒿Artemisia giraldii | 6.4 | 麻花头Klasea centauroides | 4.7 | |
| 黄鹌菜Youngia japonica | 2.5 | 蒲公英Taraxacum mongolicum | 0.9 | |
| 二裂委陵菜Potentilla bifurca | 4.0 | 小蓬草Conyza canadensis | 4.9 | |
| 刺儿菜Cephalanoplos segetum | 2.8 | 猪毛蒿Artemisia scoparia | 7.9 | |
| 中华隐子草Cleistogenes chinensis | 5.9 | 钻叶紫菀Symphyotrichum subulatum | 7.1 | |
| 二色棘豆Oxytropis bicolor | 2.2 | 白花葱Allium yanchiense | 3.9 | |
| 车前Plantago asiatica | 4.4 | 白茅Imperata cylindrica | 1.7 | |
| 翻白委陵菜Potentilla discolor | 1.6 | 白头翁Pulsatilla chinensis | 3.8 | |
| 赖草Leymus secalinus | 5.5 | 百里香Thymus mongolicus | 19.3 | |
| 野草莓Fragaria vesca | 4.4 | 地肤Kochia scoparia | 3.0 | |
| 牡蒿Artemisia japonica | 16.6 | 黄花蒿Artemisia annua | 4.5 | |
| 苜蓿Medicago sativa | 6.8 | 灰叶黄耆Astragalus discolor | 1.5 | |
| 抱茎苦荬菜 Lxeris sonchifolia | 2.9 | 藜Chenopodium album | 2.8 | |
| 地黄Rehmannia glutinosa | 1.0 | 裂叶堇菜Viola dissecta | 3.2 | |
| 杠柳Periploca sepium | 5.2 | 茅莓Rubus parvifolius | 3.9 | |
| 旱芦苇Phragmites australis | 15.1 | 披针叶黄华Thermopsis lanceolata | 8.2 | |
| 火绒草Leontopodium leontopodioides | 4.7 | 牵牛Pharbitis nil | 1.6 | |
| 牻牛儿苗Erodium stephanianum | 4.3 | 茜草Rubia cordifolia | 1.9 | |
| 野豌豆Vicia sepium | 6.1 | 沙棘Hippophae rhamnoides | 2.3 | |
| 艾蒿Artemisia argyi | 7.0 | 田葛缕子Carum buriaticum | 3.3 | |
| 百蕊草Thesium chinense | 3.7 | 乌头叶蛇葡萄Ampelopsis aconitifolia | 1.2 | |
| 柴胡Bupleurum chinense | 4.9 | 野菊Dendranthema indicum | 2.2 | |
| 大针茅Stipa grandis | 12.3 | 益母草Leonurus japonicus | 1.2 | |
| 地耳草Hypericum japonicum | 2.2 | 榆树Ulmus pumila | 2.1 | |
| 表中加粗物种为40种常见物种中的非指示物种 | ||||

|
| 图 2 Ward聚类与Mantel相关性分析 Fig. 2 Ward clustering and Mantel correlation analysis |
按照《中国植物区系和植被地理》的植被分类原则中规定的植物群落命名系统[31], 同一层中拥有相同建群种、优势种或生态指示种的植物群落称为群丛。因此, 以最大指示种对各群丛命名, 22个样地可划分为5个群丛(图 3):

|
| 图 3 22个草地样地Ward聚类分析结果 Fig. 3 Ward cluster analysis results of the investigated 22 grassland communities n表示该聚类簇下的样地数量 |
Ⅰ. 狗尾草群丛(Setaria viridis ass): 以禾本科一年生植物狗尾草为优势种, 伴生有鹅冠草(Roegneria kamoji)、猪毛菜(Salsola collina)、阿尔泰狗娃花(Aster altaicus)等。3个样地中出现, 分布于上坡, 群落平均高度(35.0±15.0)cm, 群落盖度最低(55±10.8)%, AGB最低(112.6±32.0)g/m2, BGB最低为(127.6 ±52.4)g/m2, 根冠比(1.1±0.3)。
Ⅱ. 茵陈蒿群丛(Artemisia capillaris ass): 以茵陈蒿为优势种, 伴生有早熟禾(Poa sphondylodes)、香青兰(Dracocephalum moldavica)等。在2个样地中出现, 均分布于上坡。群落平均高度(40.0±20.0)cm, 盖度(61.6±17.7)%, AGB为(239.6±62.2)g/m2, BGB为(396.1±178.9)g/m2, 根冠比(1.8±0.8)。
Ⅲ. 甘草群丛(Glycyrrhiza uralensis ass): 以甘草(Glycyrrhiza uralensis)为优势种, 伴生有草木樨状黄耆(Astragalus melilotoides)、败酱(Patrinia scabiosfolia)、小红菊(Dendranthema chanetii)等。包含4个样地, 除9号地外均分布于沟坡两侧。群落平均高度(35.0±13.0)cm, 群落盖度最高(74.0±19.2)%, AGB为(197.9±82.6)g/m2, BGB为(281.7±164.4)g/m2, 根冠比(1.5±1.1)。
Ⅳ. 铁杆蒿群丛(Artemisia sacrorum ass): 以铁杆蒿为优势种, 伴生有达乌里胡枝子(Lespedeza davurica)、菊叶委陵菜(Potentilla tanacetifoli)、野草莓(Fragaria vesca)等。在5个样地中出现, 在流域各位置均有分布。群落平均高度(32.5±13.0)cm, 盖度(68.0±9.9)%, AGB为(204.3±65.4)g/m2, BGB为(395.8±236.5)g/m2, 根冠比(2.1±1.4)。
Ⅴ. 白羊草群丛(Bothriochloa ischaemum ass): 以白羊草为优势种, 伴生有长芒草、砂珍棘豆(Oxytropis racemose)、西伯利亚远志(Polygala sibirica)等。在8个样地中出现, 多分布于中下坡, 群落平均高度(25.0±8.0)cm, 盖度(70.0±18.2)%, AGB为(195.2±59.1)g/m2, BGB为(403.8±160.5)g/m2, 根冠比(2.4±1.7)。
2.2 群丛物种组成指示种是指在生态系统中具有重要或独特生物学、生态学特征的一类物种, 它们的存在、种群密度和生理特性等信息, 表征整体群落或生态水平的状况以及变化情况。83种物种中, 有25种植物为5类群丛共有物种, 说明不同群丛物种组成存在相似性和连续性。置换检验结果表明, 21种物种在不同群丛间指示值显著差异(P < 0.05)(表 2), 物种在不同样点间分布和丰度存在显著差异, 可作为区分不同群丛的指示物种, 其余62种非指示物种基本信息见表 3。狗尾草群丛中指示物种最多(9种), 甘草群丛5种, 茵陈蒿群丛和白羊草群丛3种, 铁杆蒿群丛仅1种。白羊草、茵陈蒿、铁杆蒿、猪毛菜(Salsola collina)、鬼针草、风毛菊(Saussurea japonica)、地锦(Parthenocissus tricuspidata)和苦荬菜(Ixeris polycephala)在不同群丛间指示值呈极显著差异, 为强指示物种。砂珍棘豆、早熟禾、鹅冠草和草木樨状黄耆等植物在不同群丛间指示值呈显著差异, 为弱指示种。白羊草、茵陈蒿、铁杆蒿、狗尾草和甘草分别为5类群丛优势物种和指示种, 对草地群落结构分异产生重要影响。
2.3 群丛功能群重要值图 4结果表明, 禾本科、豆科和菊科为最重要功能群, 其在群落中的数量占75%—85%, 植被盖度占48%—82%, 地上生物量占52%—82%, 均远高于其它22科植物的总和。禾本科植物物种重要值占比为21%—36%, 豆科为5%—24%, 菊科为28%—33%, 其他科占比为15%—24%。其它22科中, 唇形科4种, 藜科3种, 萝藦科、牦牛儿苗科、葡萄科、伞形科、玄参科和远志科均各含2种, 百合科、败酱科、车前科、胡颓子科、毛茛科、木贼科、茜草科、十字花科、檀香科、藤黄科、旋花科、榆科和紫葳科均只有1种。5类群丛中不同功能群重要值差异明显(图 4), 狗尾草群丛中禾本科高草和杂类草物种重要值最高(27.8和18.3), 豆科高草和豆科矮草最低(5.3和0.4)。茵陈蒿群丛中除豆科矮草重要值最高(18.9)外, 其它6类功能群的重要值与另外4群丛无明显差异。甘草群丛禾本科高草与豆科矮草均最低(9.2和8.2), 豆科高草最高(16.4), 豆科矮草、菊科和杂类草的重要值与其它群丛无明显差异。铁杆蒿群丛菊科矮草重要值最高(26.2), 其它功能群重要值与白羊草群丛相近。白羊草群丛禾本科矮草重要值最高(16.6), 杂类草重要值最低(15.7)。

|
| 图 4 5类群丛中不同功能群物种重要值的占比 Fig. 4 The proportion of important value of different functional groups in the five associations |
土壤养分含量、群落结构特征及物种多样性参数的单因素方差分析结果表明(表 2), 白羊草、铁杆蒿和甘草群丛(7.86 g/kg、6.25 g/kg和6.38 g/kg)的SOC含量显著高于茵陈蒿和狗尾草群丛(3.22 g/kg, 4.04 g/kg)(P < 0.05)。狗尾草和茵陈蒿群丛(0.58 g/kg, 0.64 g/kg)的TN含量显著低于其它3个群丛(0.85—1.05 g/kg)(P < 0.05), 白羊草群丛最高(1.05 g/kg)。TP含量为0.49—0.57 g/kg, 5类群丛间无显著差异。土壤pH呈弱碱性, 5类群丛间无显著差异, 铁杆蒿群丛最低(8.40)(P < 0.05)。
6个群落结构特征参数中, 群落高度在不同群丛间差异不显著, 而植被密度、群落盖度、AGB、BGB和根冠比差异显著(P < 0.05)。草地群落植物密度为145—261株/m2, 铁杆蒿群丛最高, 茵陈蒿群丛最低;群落平均高度为25—37 cm, 茵陈蒿群丛最高, 白羊草群丛最低。群落平均盖度为55%—74%, 其中铁杆蒿和白羊草显著高于狗尾草群丛(P < 0.05)。茵陈蒿群丛AGB最高(239.6 g/m2), 狗尾草群丛显著低于其它4个群丛(112.6 g/m2)(P < 0.05)。白羊草群丛BGB最高(403.8 g/m2), 显著高于狗尾草群丛(127.6 g/m2)(P < 0.05)。
由表 2可知, 5类群丛物种丰富度在10—17之间, 群丛间无显著差异。Shannon-Wiener指数为1.99—2.36, 其中狗尾草群丛显著高于茵陈蒿、铁杆蒿和白羊草群丛(P < 0.05);均匀度指数为0.80—0.87, 群丛间差异性与Shannon指数相一致;优势度指数为0.70—0.84, 茵陈蒿群丛显著低于其它4个群丛(P < 0.05)。
2.5 草地群落空间分布与环境因子关系排序图中点的坐标反映各物种最适生态梯度分布状况, 样方越接近, 表示物种组成越相似, 并在一定程度上反映群落空间分布的梯度。由图 5可知, 狗尾草群丛与茵陈蒿群丛分布于第一、二象限, 与白羊草群丛、甘草群丛与铁杆蒿群丛距离最远, 说明狗尾草和茵陈蒿群丛物种组成与其它3类群丛差异最大。甘草群丛分布于一、三象限, 白羊草群丛则分布于第三象限, 铁杆蒿群丛分布于甘草群丛与白羊草群丛之间。

|
| 图 5 草地样方与环境因子及群落结构参数NMDS排序图 Fig. 5 Grassland plot NMDS ordination with environmental and community structure factors D: 优势度指数Simpson's diversity index; WH: 群落加权平均高度Community weighted height; SOC: 土壤有机碳Soil organic carbon; TN: 全氮Total nitrogen; BGB: 地下生物量Belowground biomass; R/S: 根冠比Root/Shhoot. |
NMDS排序结果表明, 海拔、坡度、坡向、土壤pH和全磷对样方分布未产生显著影响(P > 0.05), 但土壤有机碳(SOC)和全氮(TN)含量对样方分布产生显著影响(P < 0.05)。SOC和TN含量与第一轴呈显著正相关关系, 第一轴自右向左反映了土壤中C和N含量增加的梯度变化规律;群落平均高度和优势度指数与第二轴呈显著正相关关系, 第二轴自下至上基本反映了群落高度和优势度指数增加的变化规律。沿第一轴自右向左依次分布了狗尾草群丛-茵陈蒿群丛-甘草群丛-铁杆蒿群丛—白羊草群丛, 与SOC与TN增加方向相同。白羊草群丛分布于SOC和TN含量高的区域, 狗尾草、茵陈蒿和甘草群丛相反。从群落结构和物种多样性参数来看, 甘草群丛与铁杆蒿群丛群落高度与优势度指数均显著高于白羊草群丛。
2.6 环境因子对功能群与群落结构影响效应结构方程建模(SEM)结果表明, SOC、TN与植物功能群、植被盖度、AGB、BGB间存在显著相关关系(图 6)。模型拟合度为0.496, CMIN/DF为0.980, RMSEA为0。SEM一般要求卡方值越小, P > 0.05, CMIN/DF > 3, RMSEA < 0.08, 模型拟合度好, 较好解释了土壤养分含量对群落功能群和结构特征的影响。功能群对植被盖度、AGB、BGB的总解释度分别为8%, 45%, 42%。SOC对TN有显著正影响(0.74), 解释度为73%。TN对禾本科矮草和豆科高草均产生极显著正效应(0.31和0.34, P < 0.01)。不同功能群在群落中物种重要值对群落特征的影响存在显著差异, 禾本科高草、豆科植物和菊科高草均对AGB有显著正效应, 禾本科高草和豆科矮草对BGB呈显著正效应(0.20和0.26, P < 0.05), 菊科高草对地下生物量有极显著负效应(-0.40, P < 0.01)。SOC和TN均与植被盖度显著正相关(0.46和0.46, P < 0.01), 禾本科高草与豆科高草存在极显著正相关关系(0.31), 禾本科高草重要值与菊科高草则存在显著负相关关系(-0.54, P < 0.01)。

|
| 图 6 物种功能群、结构特征与土壤C、N间相关关系的结构方程模型 Fig. 6 Structural equation model of the correlation between species functional groups, structural characteristics and soil C and N content TG: 禾本科高草;SG: 禾本科矮草;TL: 豆科高草;SL: 豆科矮草;TA: 菊科高草: SA: 菊科矮草;Others:杂类草 |
聚类分析在不同聚类簇水平裁剪可得到不同结果, 分类数量越多, 相异系数越高[32]。采用Matel相关性系数和Anosim相似性分析综合判定, 将草地分为5种类型(r2=0.73, P < 0.05)时, 符合植物群丛分类客观实际。指示种分析中, 狗尾草群丛和茵陈蒿群丛以一年生物种居多, 甘草群丛和铁杆蒿群丛物种优势度不断增加, 白羊草群丛以多年生丛生禾草为优势物种, 这与众多时空替代法的研究结果相一致[2—5, 15], 表明基于物种丰度和数学方法对草地群落分类具有客观性和合理性。黄土丘陵区草地演替序列一般表现为一年生草本(狗尾草、猪毛蒿、苦卖菜等早期定居物种)-多年生草本(铁杆蒿、茭蒿、委陵菜、硬质早熟禾和草木樨状黄芪等定居-竞争相对平衡种)-多年生丛生禾草(长芒草、二色棘豆、白羊草等竞争优势种)。NMDS排序结果显示, 同一群丛聚集分布, 不同群丛存在明显的边界, 分类结果在排序图上得到很好验证。沿第一轴自右向左依次分布着狗尾草群丛-茵陈蒿群丛-甘草群丛-铁杆蒿群丛-白羊草群丛, 从物种组成以及生境适宜度分布中能够较好的说明5类群丛基本代表了草地恢复不同阶段。
经过近20年自然封育, 纸坊沟流域内草地植被盖度(55.1%—74.5%)、AGB(112.6—239.6 g/m2)、丰富度(9—17)和多样性(1.97—2.36)远高于荒漠草原[33], 与高寒草原[34]和典型草原相近[3, 35], 说明草地恢复到较好水平。研究区草地SOC和TN含量(4.04—7.86, 0.58—1.05 g/kg)低于高寒草原[28]和典型草原[3, 29], TP含量与其他草地类型相比无明显差异, 土壤C、N、P含量相对较低。黄土丘陵区长期土壤侵蚀, 导致草地TOC和TN含量相对较低, 并成为限制草地群落恢复进程的关键因素[36]。NMDS排序与结构方程模型结果均表明, 海拔、坡度、坡向、土壤pH值等因素影响均不显著, 仅SOC和TN含量影响显著, 表明在小流域尺度上, 土壤养分含量对草地群落影响较地形更大[7, 37]。狗尾草群丛-茵陈蒿群丛-甘草群丛-铁杆蒿群丛—白羊草群丛分布方向与SOC和TN增加方向相同, 表明群落演替方向与SOC和TN含量具有同步性SOC对6个主要功能群无显著影响, 但对TN产生显著正效应, 与群落盖度则存在显著正相关关系, SOC对群落的影响与恢复阶段相关性更强。狗尾草群丛SOC与根冠比最低, 群落高度较高, 而白羊草群丛恰恰相反, 生物量逐渐向地下部分转移, 根系生物量增加与SOC积累存在非常强烈的正向促进作用[38]。SOC主要来源于植物枯落物与根系分泌, 自然演替增加植物向土壤输入的C含量[19], 表明SOC与群落生产力关系更加密切, 对物种组成与功能群配比影响不显著。TN与豆科矮草间存在极显著正效应, 与豆科植物具有固氮能力相符, 根瘤菌有助于土壤N含量增加, 有利于禾本科植物生长[29]。TN对禾本科高草影响显著, 禾本科优势物种根系具有较高的丛枝菌根定植率, 相较于其他功能群有更高的N吸收能力[39]。豆科植物重要性随恢复时间延长而不断增加, 表明TN对群落影响体现在对功能群调节方面。
功能群构成可解释5类群丛群落结构显著差异的内在原因[25]。本研究中, 禾本科、豆科和菊科物种重要值比例高达75%—85%, 证实三大科在半干旱区草地群落构建中的重要性[5]。狗尾草群丛盖度、AGB和BGB均显著最低, 而Shannon-Wiener指数和Pielou优势度指数均显著高于白羊草和铁杆蒿群丛(P < 0.05), 这与狗尾草群丛中禾本科高草比例较高, 而豆科物种比例较低有关。狗尾草群丛中如狗尾草、猪毛菜和鬼针草等高草更适应更适应低碳和氮含量环境, 对光资源的竞争也更占优势, 但从植物经济学策略来看, 有限资源制约了群落生产能力, 而投入较多生物量到茎中, 无法长期存在于生态系统中[30]。以豆科高草甘草和草木樨状黄耆为优势种的甘草群丛植物密度、高度和植被盖度均较高, 但其AGB和BGB却较低, 表明豆科高草与禾本科矮草为主要功能群不合理, 限制了草地生产力。除甘草群丛外, 豆科矮草重要值均大于豆科高草, 达乌里胡枝子、砂珍棘豆、二色棘豆和苜蓿等低矮或匍匐生长的物种分布更广泛, 表明豆科植物更适宜于以次优势物种存在[7]。白羊草群丛群落盖度与生物量均较高, 禾本科高草与豆科矮草组合明显提高了植物对土壤养分资源的利用率, 也极大地提高了生产力, 体现出功能群的互补作用[39]。高草与矮草间均呈显著负相关关系(图 6), 菊科和禾本科高草较矮草重要性更高, 而豆科矮草重要性则远高于豆科高草。高草诱导的光照限制抑制了矮草物种的生长, 增加了高草在群落中的竞争排斥能力[24], 植物演替会向更利于自我生存的方向发展, 各功能群均有适宜的生态位。
白羊草、达乌里胡枝子和长芒草等均为半干旱区优良牧草, 但在5类群丛中比例仍相对较低, 提高禾本科高草和豆科矮草物种比重, 降低杂类草数量可改善草地群落物种构成与营养价值。铁杆蒿群丛仅一种强指示物种, 白羊草群丛优势种重要值明显大于狗尾草和茵陈蒿群丛, 演替中后期优势物种竞争力强, 易形成单优势种群落。群落结构提高与物种多样性水平存在负相关关系, 多样性下降可能会削弱草地应对环境干扰的稳定性和恢复力。恢复后期群落可通过施肥改变群落结构和增加群落多样性, 增加草地适口性物种数量, 避免形成单优势种群落, 提高群落稳定性与经济价值[25]。恢复初期群落应补播多年生物种, 迅速增加覆盖度, 加快群落恢复的速度。
4 结论综上所述, 纸坊沟小流域内天然草地群落恢复良好, 草地群落盖度、生物量和多样性均达到较高水平, 但土壤的C、N、P含量偏低, 并成为限制草地恢复的重要因素。草地可划分为狗尾草群丛、茵陈蒿群丛、甘草群丛、铁杆蒿群丛和白羊草群丛, 基本代表了草地恢复的不同阶段, SOC和TN含量高低与草地恢复进程具有同步性。群落中不同功能群间存在互补或拮抗作用, 禾本科高草与豆科矮草共存能显著提高对土壤养分以及光的利用, 而豆科高草作为优势种将不利于草地生产力发展。早期恢复群丛(狗尾草群丛和茵陈蒿群丛)可通过补播多年生物种及提高土壤肥力加快恢复进程;恢复中后期群丛(铁杆蒿群丛和白羊草群丛)易形成单物种群丛, 应通过人为措施改善群落结构, 扩大禾本科与豆科比例, 提高群落生物多样性与稳定性。总体而言, 土壤条件是影响纸坊沟小流域天然草地恢复进程的主要因素, 恢复调控措施选择应考虑草地群丛类型与土壤C、N含量差异。
| [1] |
Gang C C, Zhao W, Zhao T, Zhang Y, Gao X R, Wen Z M. The impacts of land conversion and management measures on the grassland net primary productivity over the Loess Plateau, Northern China. Science of the Total Environment, 2018, 645: 827-836. DOI:10.1016/j.scitotenv.2018.07.161 |
| [2] |
Deng L, Shangguan Z P. Species composition, richness and aboveground biomass of natural grassland in hilly-gully regions of the Loess Plateau, China. Journal of Integrative Agriculture, 2014, 13(11): 2527-2536. DOI:10.1016/S2095-3119(13)60590-0 |
| [3] |
Zhao H F, He H M, Wang J J, Bai C Y, Zhang C J. Vegetation restoration and its environmental effects on the Loess Plateau. Sustainability, 2018, 10(12): 4676. DOI:10.3390/su10124676 |
| [4] |
Liu Y, Wu G L, Ding L M, Tian F P, Shi Z H. Diversity-productivity trade-off during converting cropland to perennial grassland in the semi-arid areas of China. Land Degradation and Development, 2017, 28(2): 699-707. DOI:10.1002/ldr.2561 |
| [5] |
乔文静, 戴银月, 张伟, 付淑月, 杨改河, 韩新辉, 陈正兴, 高德新, 冯永忠. 黄土丘陵区撂荒恢复过程中植物群落组成与土壤养分及酶活性变化的关系. 环境科学, 2018, 39(12): 5687-5698. |
| [6] |
Zheng X X, Liu G H, Fu B J, Jin T T, Liu Z F. Effects of biodiversity and plant community composition on productivity in semiarid grasslands of Hulunbeir, Inner Mongolia, China. Annals of the New York Academy of Sciences, 2010, 1195(S1): 52-64. |
| [7] |
Borcard D, Gillet F, Legendre P. 数量生态学: R语言的应用. 赖江山, 译. 北京: 高等教育出版社, 2014: 104-108.
|
| [8] |
张金屯. 数量生态学(第三版). 北京: 科学出版社, 2018, 336-338. |
| [9] |
Wen L, Dong S K, Li Y Y, Sherman R, Shi J J, Liu D M, Wang Y L, Ma Y S, Zhu L. The effects of biotic and abiotic factors on the spatial heterogeneity of alpine grassland vegetation at a small scale on the Qinghai-Tibet Plateau (QTP), China. Environmental Monitoring and Assessment, 2013, 185(10): 8051-8064. DOI:10.1007/s10661-013-3154-y |
| [10] |
包小婷, 丁陆彬, 姚帅臣, 王景升, 石培礼, 王彤, 李超, 刘文婧. 拉萨河流域植物群落的数量分类与排序. 生态学报, 2019, 39(3): 779-786. |
| [11] |
席小康, 朱仲元, 郝祥云. 锡林河流域草原植物群落分类及其多样性分析. 生态环境学报, 2016, 25(8): 1320-1326. |
| [12] |
赵连春, 秦爱忠, 赵成章, 段凯祥, 王继伟, 文军. 嘉峪关草湖湿地植物功能群组成及其性状对不同生境的响应. 生态学报, 2020, 40(3): 822-833. |
| [13] |
乔鲜果, 郭柯, 赵利清, 刘长成, 赵海卫, 侯东杰, 高趁光. 中国石生针茅草原的分布、群落特征和分类. 植物生态学报, 2017, 41(2): 231-237. |
| [14] |
白文娟, 焦菊英, 马祥华, 温仲明, 焦峰. 黄土丘陵沟壑区退耕地自然恢复植物群落的分类与排序. 西北植物学报, 2005, 25(7): 1317-1322. DOI:10.3321/j.issn:1000-4025.2005.07.006 |
| [15] |
Biondini M. Plant diversity, production, stability, and susceptibility to invasion in restored northern tall grass prairies (United States). Restoration Ecology, 2007, 15(1): 77-87. DOI:10.1111/j.1526-100X.2006.00192.x |
| [16] |
Cleland E E, Collins S L, Dickson T L, Farrer E C, Gross K L, Gherardi L A, Hallett L M, Hobbs R J, Hsu J S, Turnbull L, Suding K N. Sensitivity of grassland plant community composition to spatial vs. temporal variation in precipitation. Ecology, 2013, 94(8): 1687-1696. DOI:10.1890/12-1006.1 |
| [17] |
Sun W Y, Song X Y, Mu X M, Gao P, Wang F, Zhao G J. Spatiotemporal vegetation cover variations associated with climate change and ecological restoration in the Loess Plateau. Agricultural and Forest Meteorology, 2015, 209-210: 87-99. DOI:10.1016/j.agrformet.2015.05.002 |
| [18] |
Wang G X, Liu G S, Li C J. Effects of changes in alpine grassland vegetation cover on hillslope hydrological processes in a permafrost watershed. Journal of Hydrology, 2012, 444-445: 22-33. DOI:10.1016/j.jhydrol.2012.03.033 |
| [19] |
Zhang W, Ren C J, Deng J, Zhao F Z, Yang G H, Han X H, Tong X G, Feng Y Z. Plant functional composition and species diversity affect soil C, N, and P during secondary succession of abandoned farmland on the Loess Plateau. Ecological Engineering, 2018, 122: 91-99. DOI:10.1016/j.ecoleng.2018.07.031 |
| [20] |
张凯, 陈丽茹, 徐慧敏, 李秧秧. 黄土高原水蚀风蚀交错带小流域植物群落特征的空间变异及其影响因素. 应用生态学报, 2019, 30(8): 2521-2530. |
| [21] |
王建国, 樊军, 王全九, 王力. 黄土高原水蚀风蚀交错区植被地上生物量及其影响因素. 应用生态学报, 2011, 22(3): 556-564. |
| [22] |
吴会峰, 宋丽娟, 杜峰, 郝文芳. 黄土丘陵区草地主要种群生态位及其环境解释. 应用生态学报, 2017, 28(11): 3494-3504. |
| [23] |
Gonzalez M M, Crofts A L, Mclaren J R. Plant biomass, rather than species composition, determines ecosystem properties: Results from a long-term graminoid removal experiment in a northern Canadian grassland. Journal of Ecology, 2019, 107(5): 2211-2225. DOI:10.1111/1365-2745.13169 |
| [24] |
Xu Z W, Ren H Y, Li M H, Van Ruijven J, Han X G, Wan S Q, Li H, Yu Q, Jiang Y, Jiang L. Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony. Journal of Ecology, 2015, 103(5): 1308-1316. DOI:10.1111/1365-2745.12441 |
| [25] |
张学霞, 朱清科, 吴根梅, 孙丹珠. 数码照相法估算植被盖度. 北京林业大学学报, 2008, 30(1): 164-169. DOI:10.3321/j.issn:1000-1522.2008.01.030 |
| [26] |
刘海威, 张少康, 焦峰. 黄土丘陵区不同退耕年限草地群落特征及其土壤水分养分效应. 草业学报, 2016, 25(10): 31-39. DOI:10.11686/cyxb2016175 |
| [27] |
MacDonald R L, Burke J M, Chen H Y H, Prepas E E. Relationship between aboveground biomass and percent cover of ground vegetation in Canadian boreal plain riparian forests. Forest Science, 2012, 58(1): 47-53. DOI:10.5849/forsci.10-129 |
| [28] |
Chen Z F, Xiong P F, Zhou J J, Yang Q, Wang Z, Xu B C. Grassland productivity and diversity changes in responses to N and P addition depend primarily on tall clonal and annual species in semiarid Loess Plateau. Ecological Engineering, 2020, 145: 105727. DOI:10.1016/j.ecoleng.2020.105727 |
| [29] |
熊沛枫. 黄土丘陵区草地群落冠层降雨截留特征及优势种对降雨的响应[D]. 杨凌: 西北农林科技大学, 2019.
|
| [30] |
Wiesmair M, Otte A, Waldhardt R. Relationships between plant diversity, vegetation cover, and site conditions: implications for grassland conservation in the Greater Caucasus. Biodiversity and Conservation, 2017, 26(2): 273-291. DOI:10.1007/s10531-016-1240-5 |
| [31] |
陈灵芝. 中国植物区系与植被地理. 北京: 科学出版社, 2014.
|
| [32] |
袁秀, 马克明, 王德. 草地植物群落最优分类数的确定——以黄河三角洲为例. 生态学报, 2013, 33(8): 2514-2521. |
| [33] |
陈林, 辛佳宁, 苏莹, 李月飞, 宋乃平, 王磊, 杨新国, 卞莹莹, 田娜. 异质生境对荒漠草原植物群落组成和种群生态位的影响. 生态学报, 2019, 39(17): 6187-6205. |
| [34] |
熊定鹏, 赵广帅, 武建双, 石培礼, 张宪洲. 羌塘高寒草地物种多样性与生态系统多功能关系格局. 生态学报, 2016, 36(11): 3362-3371. |
| [35] |
Kang M Y, Dai C, Ji W Y, Jiang Y, Yuan Z Y, Chen H Y H. Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia grasslands, China. PLos One, 2013, 8(7): e69561. DOI:10.1371/journal.pone.0069561 |
| [36] |
Zhu H X, Fu B J, Wang S, Zhu L H, Zhang L W, Jiao L, Wang C. Reducing soil erosion by improving community functional diversity in semi-arid grasslands. Journal of Applied Ecology, 2015, 52(4): 1063-1072. DOI:10.1111/1365-2664.12442 |
| [37] |
Liu B, Zhao W Z, Liu Z L, Yang Y T, Luo W C, Zhou H, Zhang Y Y. Changes in species diversity, aboveground biomass, and vegetation cover along an afforestation successional gradient in a semiarid desert steppe of China. Ecological Engineering, 2015, 81: 301-311. DOI:10.1016/j.ecoleng.2015.04.014 |
| [38] |
Yang Y, Dou Y X, An S S. Environmental driving factors affecting plant biomass in natural grassland in the Loess Plateau, China. Ecological Indicators, 2017, 82: 250-259. DOI:10.1016/j.ecolind.2017.07.010 |
| [39] |
Luo R Y, Fan J L, Wang W J, Luo J F, Kuzyakov Y, He J S, Chu H Y, Ding W X. Nitrogen and phosphorus enrichment accelerates soil organic carbon loss in alpine grassland on the Qinghai-Tibetan Plateau. Science of the Total Environment, 2019, 650: 303-312. DOI:10.1016/j.scitotenv.2018.09.038 |