 2021, Vol. 41
2021, Vol. 41文章信息
- 李明娟, 赵娟娟, 江南, 潘妮, 张曼琳, 束承继
- LI Mingjuan, ZHAO Juanjuan, JIANG Nan, PAN Ni, ZHANG Manlin, SHU Chengji
- 深圳近海公园植物群落结构特征及其与景观格局的相关性
- Characteristics of plant community structure and its relationship with landscape pattern in Shenzhen offshore parks
- 生态学报. 2021, 41(22): 8732-8745
- Acta Ecologica Sinica. 2021, 41(22): 8732-8745
- http://dx.doi.org/10.5846/stxb202105291415
-
文章历史
- 收稿日期: 2021-05-29

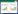




近年来, 随着城市化进程的加快, 生物多样性面临着严重挑战[1]。深圳是中国乃至全球快速城市化的代表之一[2-3]。尽管城市地域狭小、人口剧增、土地资源有限, 但由于城市绿地建设、养护与管理受到重视, 深圳城市绿化目前已处于国内领先水平[4]。城市公园植物是城市绿地中提供生态服务的主体, 具有固碳释氧、降温增湿、防风减噪等生态功能, 对城市气候与环境具有较大的影响[5]。公园植物群落也为城市昆虫、鸟类等多种动物提供重要栖息地[6], 其多样性水平对城市生态系统的健康与稳定具有重要作用。随着生态文明建设的推进, 人们对城市生态环境的保护意识与需求不断提高。在未来, 良好的城市环境也将成为城市可持续发展的关键优势与竞争力[4]。
城市公园作为城市绿岛, 其植物种类繁多, 拥有大量生长繁茂的乔木、灌木、草本花卉和野生草本[7-8]。城市土地利用类型复杂, 景观斑块类型众多, 而城市公园中的植物群落往往是城市植物多样性最高的景观斑块[9]。景观的结构和连通性对生态功能和生态过程有重要影响[10-11]。已有研究表明, 公园内部绿地景观斑块的大小、形状和数目及其边缘效应等对斑块内部的植物丰富度和群落结构模式具有显著影响[12-13]。例如, 林地景观斑块的面积越大、连通性越强, 更有利于维持公园内部的植物多样性[14]。城市公园的景观格局较大程度上由公园设计者决定, 植物群落也受到人为干扰的重要影响, 但亦具有自然属性。因此, 研究和合理设计公园植物群落, 使之与公园景观格局相适应, 对提高公园植物群落多样性十分重要。
深圳作为一个沿海城市, 其近海公园数量较多。与内陆城市公园相比, 近海城市公园往往具有更特殊的地理位置与环境条件。以往对近海公园的研究多针对植物景观评价[15]、物种多样性[16]、单纯的景观格局[17]等某一主题进行研究, 很少对近海公园植物群落结构进行全面而多层次的分析报道。关于群落结构特征与公园景观格局相关性的研究更是缺乏[18-19]。基于此, 本研究在实地调查的基础上, 选取深圳海岸附近生态环境良好, 植物群落具有代表性的公园进行研究。将实地调查数据与遥感数据相结合, 从局部到整体分析公园植物群落特征, 并探讨其对公园景观格局的影响。有助于更好地理解园林景观中植物群落与宏观景观格局的相互关系的规律[20], 为合理设计植物景观提供依据, 对提高城市绿化质量以及健全生物多样性保护机制具有重要意义。
1 研究地区与研究方法 1.1 研究区概况深圳市位于广东省南部, 东经113°43′—114°38′, 北纬22°24′—22°52′之间, 地势东高西低, 以低丘陵地为主, 西部为滨海平原。其建成区面积为923.2 km2, 属亚热带海洋性气候, 地带性植被属于南亚热带季风常绿阔叶林, 常年平均气温23.0℃, 年均降雨量1935.8 mm[21]。本研究选择了深圳市具有代表性的6个近海公园, 其地理分布(图 1)及特征概况(表 1)如下。

|
| 图 1 深圳近海公园的研究区范围及样地分布 Fig. 1 Research area and sample distribution of offshore parks in Shenzhen |
| 公园名称 Park name |
地理位置 Location |
建成年限 Construction-completed year |
公园面积 Park area/hm2 |
公园类型 Park type |
| 洪湖公园Honghu park | 罗湖区 | 1984 | 62.14 | 人工型 |
| 荔枝公园Lizhi park | 福田区 | 1982 | 29.37 | 人工型 |
| 香蜜公园Xiangmi park | 福田区 | 2017 | 49.16 | 人工型 |
| 东湖公园Donghu park | 罗湖区 | 1961 | 91.48 | 半自然型 |
| 海上田园公园Haishangtianyuan park | 宝安区 | 1999 | 102.74 | 半自然型 |
| 华侨城湿地公园Huaqiaocheng park | 南山区 | 2016 | 67.97 | 半自然型 |
采用基于百米网格的分层随机抽样法。首先, 采用100 m×100 m的等距网格覆盖深圳市近海公园的遥感图像, 然后, 在公园中随机抽取网格作为实地调查的大样地, 各公园抽样调查面积不小于公园总面积的15%。将百米网格大样地中被道路、水体分开的每个绿地斑块, 记为不同的样方。以大样地中的绿地斑块样方为单位进行实地调查。
本次植物调查只针对乔木、灌木和草本三类生活型进行, 藤本植物等其他类别的植物不进行调查记录。以绿地斑块为单位, 分别调查每个样方斑块中的所有乔、灌植物种, 同时在每个样方斑块中设置1 m×1 m的草本样方, 对草本植物典型群落进行抽样调查。草本样方量采用物种数累积法确定。
调查指标包括:(1)乔木、灌木植物的种名、株数、株高、冠幅、生长状况。(2)草本植物的种名、频数、盖度、株高、生长状况。植物分类参照《中国植物志》中采用的恩格勒系统, 本研究所指的乔木是指有明显主干, 株高3米以上的植物。详细的调查方法与指标测定方法参考美国UFORE模型的调查手册和i-Tree-Eco4.16.2017[22]。
1.2.2 景观格局数据以SPOT6/7高分辨率卫星影像为主要数据源, 分辨率1.5 m, 时相为2017-12-19。首先用ENVI将样本公园按照边界线剪裁, 再利用高分影像分类软件eCognition进行分类, 分为林地、草地、水体和人工表面四类[23]。同时通过GPS定位、实地踏勘确定斑块边界, 校正相关数据, 进一步确定分类结果, 并进行精度计算(>90%)。最后, 用Fragstas进行景观分析并导出景观指数值。选择的指标包括斑块大小、形状、类型等对景观结构具有重要意义的指标[24](表 2), 其计算公式参考布仁仓等人的研究[25]。
| 层次 Category |
景观指数 Landscape index |
描述 Description |
生态学含义 Ecological implications |
| 类别Class | AREA_MN | 平均斑块面积 | 表征景观破碎度 |
| NP | 斑块数量 | 表征景观破碎度 | |
| PARA | 周长面积比 | 表征景观形状的复杂程度 | |
| PAFRAC | 周长面积分维数 | 表征景观形状的复杂程度 | |
| FRAC_MN | 平均分维数 | 表征景观形状的复杂程度 | |
| GYRATE_MN | 平均回转半径 | 表征景观斑块的大小 | |
| 类别/景观 | PD | 斑块密度 | 表征景观破碎度 |
| Class/Landscape | LPI | 最大斑块指数 | 表征最大斑块占据整个景观面积的比例 |
| WLPI | 林地最大斑块指数 | 表征林地最大斑块占整个景观面积的比例 | |
| 景观Landscape | SHAPE | 斑块形状指数 | 表征整体景观的形状复杂程度 |
| URB% | 人工表面比例 | 景观中人工表面占整个景观面积的比例 | |
| SHDI | 景观多样性 | 斑块类型与数量在景观中分布的均衡状况 | |
| SHEI | 景观均匀度 | 可与优势度指数转换, 两者之间呈反比关系 | |
| AREA_MN:平均斑块面积Mean patch area; NP:斑块数量Number of patches; PARA:周长面积比Perimeter area ratio; PAFRAC:周长面积分维数Perimeter area fractal dimension; FRAC_MN:平均分维数Mean fractal dimension index; GYRATE_MN:平均回转半径Mean radius of gyration; PD:斑块密度Patch density; LPI:最大斑块指数The largest patch index; WLPI:林地最大斑块指数The largest patch index for woodland; SHAPE:斑块形状指数Shape index; URB%:人工表面比例Proportion of urban constructed land; SHEI:景观多样性Shannon-Wiener diversity index;SHEI:景观均匀度Shannon-Wiener evenness index | |||
城市公园植物群落是高度人工化的群落, 并且受到人为设计、养护管理等强烈干扰。草本植物位于群落结构的最底层, 拥有良好的土壤条件, 其中通常含有许多植物的种子。相比乔灌层, 草本层经常会发生一些自生植物、野生植物的演替。一些野生杂草与乔灌植物的幼苗, 若未影响到主体景观的观赏效果, 通常不会进行专门的清理。在有些维护较差的绿地中, 草本层几乎已演替为自然群落[26]。因此本文将乔灌层和草本层分开进行分析。
使用PC-ORD 5.0软件, 以乔灌草植物重要值为基础进行TWINSPAN分类。将植物分为乔灌植物和草本植物两大类, 剔除乔灌和草本数据中总频度小于等于2%的偶见种。参数设置:“假种(Pseudo species)”切割水平采用5级, 各水平值分别是0.00、0.02、0.05、0.10、0.20;每次分类的指示种的最大数量为5个, 分类的最大水平数为6次, 分类最小种组采用5个。
1.3.2 物种多样性计算物种多样性评价指标选取了Patrick物种丰富度指数(S)、Simpson指数(D)、Shannon-Wiener指数(H)以及Pielou均匀度指数(E), 其计算公式参考张峰和张金屯等人的研究[27]。
1.3.3 相似度分析利用Jaccard β相似性系数来评价植物群落相似度, 计算公式为:
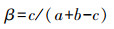
|
式中, a、b分别为两公园的物种数, c为两公园的共有物种数。其数值越大, 代表群落相似性程度越大[28]。
1.3.4 群落稳定性计算在生态学的研究中, 关于植物群落稳定性的定义较多, 其具体概念往往是和特定研究相联系的[29-30]。本研究采用Godron贡献定律法[31]进行稳定性测定, 同时参考了刘瑞雪等在人为干扰影响下的城市植物中的研究方法[32-33]。基于数量生态学的原理, 该方法可提供植物群落稳定与否的信息, 适于用来探讨人工植物群落配置的合理性与低维护性[34]。
将植物群落中所有植物的频度从大到小排列, 计算出相对频度, 并以由大到小的顺序逐步累加起来, 再计算物种总数倒数的累积百分数, 并和相对频度的累积百分数一一对应。通过Matlab对两者作散点图, 用一条曲线连接各散点, 同时在坐标轴数值的两端点处连一条直线, 与曲线交点即为所求交点。所求交点坐标越趋近于(20, 80)的稳定点, 群落越稳定, 反之则越不稳定。
1.3.5 相关性分析及描述性统计分析相关性分析及描述性统计分析通过IBM SPSS Statistics(Version 16)软件完成。
2 结果与分析 2.1 近海公园植物群落结构特征 2.1.1 公园植物群落结构根据重要值排序确定植物群落优势种, TWINSPAN将深圳近海公园共161个乔灌样地划分为20组, 其中样地数大于总量3%的有6组, 共141个样地, 占总样地量的87%;将146个草本样地划分为21组, 其中样地量大于总量10%的有3组, 共76个样地, 占总样地量的60%。
由表 3可知, 从样地量来看, TS-4是近海公园中最常见的乔灌群落类型, H-1是最常见的草本群落类型。其中TS-4在东湖公园、海上田园、华侨城湿地公园、香蜜公园中分别占比为56%、35%、94%、33%。从指示种来看, 乔灌指示种中观赏植物出现较多。标志性乔木中, 小叶榄仁(Terminalia neotaliala)、羊蹄甲(Bauhinia purpurea)、黄花风铃木(Handroanthus chrysanthus)等多个树种具有热带性特征, 也是华南地区的常见乡土树种。标志性灌木中, 以翠芦莉(Ruellia simplex)、龙船花(Ixora chinensis)、朱槿(Hibiscus rosa-sinensis)等为主, 形成了色彩丰富的花灌木景观;草本指示种中出现了两耳草(Paspalum conjugatum)、短叶水蜈蚣(Kyllinga brevifolia)、香附子(Cyperus rotundus)等多种自生草本植物, 也发现喜旱莲子草(Alternanthera philoxeroides)、地毯草(Axonopus compressus)等具有入侵风险的植物。
| 植物类别 Plant category |
编号 Code |
样地量 Plot size |
指示种 Indicator species |
占各公园样地量比例 The proportion of sample plots in each park/% |
主要伴生种 Main associated species |
|||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | |||||
| 乔灌植物 Trees and shrubs |
TS-1 | 9 | 小叶榄仁+羊蹄甲 | 0 | 10 | 4 | 6 | 1 | 11 | 毛果杜英(Elaeocarpus rugosus)+蒲桃(Syzygium jambos)+南洋楹(Falcataria moluccana)+黄槿(Hibiscus tiliaceus) |
| TS-2 | 5 | 黄花风铃木+海南菜豆树(Radermachera hainanensis)+垂叶榕(Ficus benjamina)+人面子(Dracontomelon duperreanum) | 12 | 0 | 0 | 0 | 0 | 4 | 玫红栎铃木(Tabebuia rosea)+榕树(Ficus microcarpa)+木棉(Bombax ceiba)+澳洲鸭脚木(Schefflera actinophylla) | |
| TS-3 | 6 | 叶子花(Bougainvillea spectabilis)+四季桂(Osmanthus fragrans (Fragrans Group)+白兰(Michelia×alba) | 3 | 0 | 4 | 0 | 3 | 0 | ||
| TS-4 | 63 | 龙船花+朱槿+灰莉(Fagraea ceilanica) | 56 | 35 | 13 | 94 | 5 | 33 | 榕树+马缨丹(Lantana camara)+金叶假连翘(Duranta erecta ′Golden Leaves′)+凤凰木+黄金榕(Ficus microcarpa ′Golden Leaves′)+垂叶榕 | |
| TS-5 | 9 | 翠芦莉+蒲葵(Livistona chinensis)+假连翘(Duranta erecta)+变叶木(Codiaeum variegatum)+小叶榕 | 0 | 5 | 18 | 0 | 0 | 0 | ||
| TS-6 | 49 | 红背桂(Excoecaria cochinchinensis)+鹅掌藤 (Schefflera arboricola) |
29 | 20 | 60 | 0 | 8 | 0 | 朱槿+朱蕉(Cordyline fruticosa)+灰莉+假槟榔(Archontophoenix alexandrae)+黄葛树+花叶艳山姜(Alpinia zerumbet ′Variegata′)+金脉爵床(Sanchezia oblonga) | |
| 草本植物Herbs | H-1 | 39 | 两耳草+短叶水蜈蚣+喜旱莲子草+酢浆草 | 18 | 0 | 31 | 0 | 17 | 7 | |
| H-2 | 21 | 巴西鸢尾(Neomarica gracilis)+水鬼蕉(Hymenocallis littoralis)+海芋(Alocasia odora)+弓果黍(Cyrtococcum patens) | 26 | 5 | 13 | 0 | 3 | 7 | 香附子+华南毛蕨(Cyclosorus parasiticus) | |
| H-3 | 16 | 香附子+华南毛蕨+地毯草 | 1 | 0 | 7 | 0 | 5 | 4 | 纤梗叶下珠(Phyllanthus tenellus)+ 犁头尖(Typhonium blumei) |
|
| TS:乔灌植物Trees and shrubs;H:草本植物Herbs;Ⅰ为东湖公园, Ⅱ为海上田园, Ⅲ为洪湖公园, Ⅳ为华侨城湿地公园, Ⅴ为荔枝公园, Ⅵ为香蜜公园 | ||||||||||
从表 4可以看出, 近海公园植物群落总体丰富度极高, 均大于150种。从植物层次来看, 灌木层植物种数低于草本层和乔木层;从植物类别来看, 除香蜜公园外, 近海公园群落层乡土植物占比均高于50%, 即以乡土植物为主。在各植物层次中, 乡土植物比例大都以草本植物最高。
| 公园 Park name |
植物层 Plant layer |
植物类别 Plant category |
植物种数 Number of species |
乡土植物种 的比例 The proportion of native plant species |
Shannon- Wiener指数 Shannon- Wiener index |
Pielou指数 Pielou index |
Simpson指数 Simpson index |
| 东湖公园 | 草本层 | 总体 | 77 | 66.23 | 3.24 | 0.75 | 0.92 |
| 乡土 | 51 | 3.18 | 0.81 | 0.94 | |||
| 灌木层 | 总体 | 74 | 40.54 | 2.77 | 0.64 | 0.88 | |
| 乡土 | 30 | 1.77 | 0.52 | 0.72 | |||
| 乔木层 | 总体 | 99 | 46.46 | 3.79 | 0.83 | 0.96 | |
| 乡土 | 46 | 3.39 | 0.89 | 0.96 | |||
| 群落 | 总体 | 250 | 50.8 | 3.69 | 0.67 | 0.93 | |
| 乡土 | 122 | 2.77 | 0.58 | 0.83 | |||
| 海上田园 | 草本层 | 总体 | 57 | 63.16 | 3.17 | 0.79 | 0.93 |
| 乡土 | 36 | 2.81 | 0.78 | 0.9 | |||
| 灌木层 | 总体 | 40 | 47.5 | 2.69 | 0.73 | 0.9 | |
| 乡土 | 19 | 2 | 0.68 | 0.75 | |||
| 乔木层 | 总体 | 68 | 45.59 | 3.25 | 0.77 | 0.93 | |
| 乡土 | 31 | 2.16 | 0.63 | 0.77 | |||
| 群落 | 总体 | 165 | 52.12 | 3.95 | 0.77 | 0.97 | |
| 乡土 | 80 | 3.32 | 0.76 | 0.93 | |||
| 华侨城湿地公园 | 草本层 | 总体 | 75 | 56 | 3.07 | 0.71 | 0.9 |
| 乡土 | 42 | 2.79 | 0.75 | 0.89 | |||
| 灌木层 | 总体 | 38 | 52.63 | 2.11 | 0.58 | 0.76 | |
| 乡土 | 20 | 1.63 | 0.54 | 0.68 | |||
| 乔木层 | 总体 | 70 | 52.86 | 3.08 | 0.72 | 0.89 | |
| 乡土 | 37 | 2.4 | 0.67 | 0.81 | |||
| 群落 | 总体 | 183 | 54.1 | 3.49 | 0.67 | 0.91 | |
| 乡土 | 96 | 2.82 | 0.62 | 0.85 | |||
| 洪湖公园 | 草本层 | 总体 | 66 | 54.55 | 2.48 | 0.59 | 0.81 |
| 乡土 | 36 | 2.46 | 0.69 | 0.83 | |||
| 灌木层 | 总体 | 56 | 57.14 | 2.32 | 0.58 | 0.82 | |
| 乡土 | 32 | 1.73 | 0.5 | 0.67 | |||
| 乔木层 | 总体 | 75 | 48 | 3.62 | 0.84 | 0.96 | |
| 乡土 | 36 | 3.06 | 0.85 | 0.93 | |||
| 群落 | 总体 | 197 | 52.79 | 2.85 | 0.54 | 0.86 | |
| 乡土 | 100 | 2.24 | 0.49 | 0.74 | |||
| 荔枝公园 | 草本层 | 总体 | 83 | 50.6 | 2.9 | 0.66 | 0.82 |
| 乡土 | 42 | 3.12 | 0.83 | 0.94 | |||
| 灌木层 | 总体 | 40 | 62.5 | 2.57 | 0.7 | 0.88 | |
| 乡土 | 25 | 2.06 | 0.64 | 0.8 | |||
| 乔木层 | 总体 | 53 | 47.17 | 3.15 | 0.79 | 0.93 | |
| 乡土 | 25 | 2.34 | 0.73 | 0.85 | |||
| 群落 | 总体 | 176 | 52.27 | 3.53 | 0.68 | 0.94 | |
| 乡土 | 89 | 2.88 | 0.64 | 0.88 | |||
| 香蜜公园 | 草本层 | 总体 | 65 | 58.46 | 3.06 | 0.73 | 0.9 |
| 乡土 | 38 | 2.32 | 0.64 | 0.79 | |||
| 灌木层 | 总体 | 60 | 31.67 | 2.9 | 0.71 | 0.88 | |
| 乡土 | 19 | 1.85 | 0.63 | 0.75 | |||
| 乔木层 | 总体 | 54 | 50 | 2.38 | 0.6 | 0.74 | |
| 乡土 | 27 | 1.36 | 0.41 | 0.47 | |||
| 群落 | 总体 | 179 | 46.93 | 3.73 | 0.72 | 0.94 | |
| 乡土 | 83 | 2.75 | 0.62 | 0.87 |
深圳近海公园中(表 5), 人工型和半自然型公园的Jaccard β多样性指数在不同的植物层间表现为:乔木层(0.46)>草本层(0.45)>灌木层(0.35), 其中, 乔木层、草本层相似性指数较大, 且与总体相似性指数相近, 灌木层相似性指数最低, 明显低于总体层。乡土层植物相似性指数为0.47, 略高于总体层。
| 植物层次 Plant level |
Jaccard β多样性指数 Jaccard β diversity |
植物层次 Plant level |
Jaccard β多样性指数 Jaccard β diversity |
|
| 草本Herbs | 0.45 | 乡土Native plants | 0.47 | |
| 灌木Shrubs | 0.35 | 总体Total plant communities | 0.46 | |
| 乔木Trees | 0.46 |
人工型公园与半自然公园两两比较的Jaccard β多样性指数如表 6, 可以看出, 香蜜公园与海上田园的相似性指数最低, 为0.23;东湖公园与洪湖公园的相似性指数最高, 为0.36。总体上, 各公园之间的相似性指数处于0.23—0.36之间, 远低于总体层次上的相似性指数(0.46)。
| 公园名称Park name | 洪湖公园 | 荔枝公园 | 香蜜公园 |
| 东湖公园 | 0.36 | 0.33 | 0.31 |
| 海上田园 | 0.35 | 0.28 | 0.23 |
| 华侨城湿地公园 | 0.28 | 0.28 | 0.28 |
从下表 7可看出, 近海公园植物群落的稳定性, 基本呈现灌木层>总体层>草本层>乔木层的规律。总体植物群落层次上, 半自然公园的稳定性得分分别为96.89, 96.79, 96.14, 高于人工型公园(94.4, 93.17, 88.97)。草本层中, 海上田园、东湖公园植物群落偏离稳定点较远(距离9.37、10.15);灌木层中, 各公园植物群落离稳定点普遍较近(平均距离小于5);乔木层中东湖公园植物群落偏离稳定点较远(距离10.92)。
| 公园类型 Park type |
公园名称 Park name |
植物层 Plant layer |
拟合曲线 Fitting curve |
P | 交点坐标 Intersection coordinates |
距离 Distance |
得分 Score |
| 半自然型 | 东湖公园 | 草本层 | y=-0.0103x2+1.633x+36.0739 | < 0.001 | (27.17,72.82) | 10.15 | 89.85 |
| Half natural type | 灌木层 | y=-0.0089x2+1.2340x+60.0421 | < 0.001 | (19.38,80.61) | 0.87 | 99.13 | |
| 乔木层 | y=-0.0116x2+1.7916x+31.5557 | < 0.001 | (27.72,72.27) | 10.92 | 89.08 | ||
| 总体层 | y=-0.0103x2+1.5187x+45.5703 | < 0.001 | (23.96,76.04) | 3.11 | 96.89 | ||
| 海上田园 | 草本层 | y=-0.0098x2+1.5561x+38.92 | < 0.001 | (26.62,73.37) | 9.37 | 90.63 | |
| 灌木层 | y=-0.0116x2+1.6778x+42.4356 | < 0.001 | (24.00,75.99) | 5.66 | 94.34 | ||
| 乔木层 | y=-0.0107x2+1.5896x+41.7839 | < 0.001 | (25.07,74.92) | 7.18 | 92.82 | ||
| 总体层 | y=-0.0105x2+1.6594x+34.1625 | < 0.001 | (27.80,72.20) | 3.21 | 96.79 | ||
| 华侨城湿地公园 | 草本层 | y=-0.0085x2+1.2934x+51.5359 | < 0.001 | (23.11,76.88) | 4.41 | 95.59 | |
| 灌木层 | y=-0.0063x2+0.9273x+66.4206 | < 0.001 | (19.23,80.76) | 1.08 | 98.92 | ||
| 乔木层 | y=-0.0089x2+1.3722x+47.6040 | < 0.001 | (24.30,75.69) | 6.09 | 93.91 | ||
| 总体层 | y=-0.0096x2+1.3932x+50.53 | < 0.001 | (22.73,77.27) | 3.86 | 96.14 | ||
| 人工型 | 洪湖公园 | 草本层 | y=-0.0063x2+0.9273x+66.4199 | < 0.001 | (18.54,81.45) | 2.06 | 97.94 |
| Artificial type | 灌木层 | y=-0.0075x2+1.0324x+66.9500 | < 0.001 | (17.37,82.62) | 3.71 | 96.29 | |
| 乔木层 | y=-0.0116x2+1.8270x+28.8152 | < 0.001 | (28.54,71.45) | 12.08 | 87.92 | ||
| 总体层 | y=-0.0072x2+1.0057x+66.5717 | < 0.001 | (17.80,82.20) | 5.6 | 94.4 | ||
| 荔枝公园 | 草本层 | y=-0.0081x2+1.2436x+53.3051 | < 0.001 | (22.68,77.31) | 3.8 | 96.2 | |
| 灌木层 | y=-0.0102x2+1.4715x+49.3554 | < 0.001 | (22.59,77.40) | 3.67 | 96.33 | ||
| 乔木层 | y=-0.0110x2+1.6876x+36.2488 | < 0.001 | (26.63,73.36) | 9.38 | 90.62 | ||
| 总体层 | y=-0.0095x2+1.3758x+51.7971 | < 0.001 | (22.27,77.73) | 6.83 | 93.17 | ||
| 香蜜公园 | 草本层 | y=-0.0096x2+1.4663x+45.2557 | < 0.001 | (24.55,75.44) | 6.44 | 93.56 | |
| 灌木层 | y=-0.0105x2+1.5434x+45.9254 | < 0.001 | (23.56,76.43) | 5.04 | 94.96 | ||
| 乔木层 | y=-0.0061x2+0.9841x+60.1536 | < 0.001 | (21.50,78.49) | 2.13 | 97.87 | ||
| 总体层 | y=-0.0105x2+1.5643x+42.7914 | < 0.001 | (24.83,75.17) | 11.03 | 88.97 |
可以看出, 各近海公园中都包含水体景观元素(表 8), 其中海上田园、洪湖公园、华侨城湿地公园水体景观比例较大, 分别达39.90%、38.30%和50.90%。近海公园中林地景观面积都大于草地景观, 人工表面景观占比最低。
| 公园名称 Park name |
林地 Forest |
草地 Grassland |
人工表面 Artificial surface |
水体 Waterbody |
|||||||
| 面积 Area/hm2 |
% | 面积 Area/hm2 |
% | 面积 Area/hm2 |
% | 面积 Area/hm2 |
% | ||||
| 东湖公园 | 59.50 | 65.10 | 11.74 | 12.8 | 13.37 | 14.60 | 6.86 | 7.50 | |||
| 海上田园 | 43.36 | 42.20 | 5.68 | 5.50 | 12.61 | 12.20 | 41.09 | 39.90 | |||
| 洪湖公园 | 30.55 | 49.10 | 0.87 | 1.30 | 6.91 | 11.10 | 23.82 | 38.30 | |||
| 华侨城湿地公园 | 14.41 | 21.20 | 18.29 | 26.90 | 0.68 | 1.00 | 34.60 | 50.90 | |||
| 荔枝公园 | 17.06 | 58.10 | 4.08 | 13.80 | 2.22 | 7.50 | 6.01 | 20.40 | |||
| 香蜜公园 | 22.29 | 45.30 | 13.65 | 27.70 | 11.57 | 23.50 | 1.65 | 3.30 | |||
公园整体景观格局指数如下表 9。从斑块密度来看, 人工型公园如洪湖公园、荔枝公园斑块密度较大, 分别为2.46、2.40。半自然型公园(东湖公园)斑块密度最小, 平均斑块面积最大。东湖公园、荔枝公园的林地最大斑块指数最高。但各个公园间的景观多样性、景观均匀度相差不大。
| 公园名称 Park name |
斑块密度 PD/(n/hm2) |
平均斑块面积 AREA_MN/hm2 |
斑块形状指数 SHAPE |
林地最大斑块指数 WLPI |
景观多样性 SHDI |
景观均匀度 SHEI |
| 东湖公园 | 1.17 | 0.86 | 1.87 | 54.06 | 1.02 | 0.73 |
| 海上田园 | 1.89 | 0.53 | 1.68 | 40.8 | 1.15 | 0.83 |
| 洪湖公园 | 2.46 | 0.41 | 1.75 | 41.97 | 1.02 | 0.74 |
| 华侨城湿地公园 | 1.87 | 0.42 | 1.68 | 5.94 | 1.07 | 0.77 |
| 荔枝公园 | 2.40 | 0.53 | 1.73 | 56.73 | 1.11 | 0.80 |
| 香蜜公园 | 1.72 | 0.58 | 2.00 | 43.49 | 1.17 | 0.73 |
经检验, 公园中各景观格局指数、植物群落结构特征指数均呈正态分布。从表 10中可看出, 公园斑块密度和林地面积比、林地最大斑块面积比呈显著负相关(P < 0.05)。斑块密度变大时, 景观破碎度增大, 不利于大型植物斑块的保存。斑块形状指数、人工表面比例与乡土植物比例呈显著负相关(P < 0.01), 可能是因为乡土植物更容易受到人为干扰的不利影响。
| 植物群落结构特征指数 Plant community characteristic indexes |
斑块密度 PD/(n/hm2) |
斑块形状指数 SHAPE |
人工表面比例 URB% |
景观多样性 SHDI |
景观均匀度 SHEI |
| 林地面积比Proportion of woodland | -0.831* | 0.387 | 0.456 | -0.226 | 0.217 |
| 林地最大斑块指数 The largest patch index for woodland |
-0.822* | 0.387 | 0.553 | 0.038 | 0.054 |
| 物种丰富度Species richness | -0.491 | 0.345 | 0.140 | -0.739 | 0.646 |
| Shannon-Wiener指数Shannon-Wiener index | -0.508 | 0.171 | 0.277 | 0.650 | -0.405 |
| Pielou指数Pielou index | -0.367 | 0.088 | 0.238 | 0.785 | -0.524 |
| Simpson指数Simpson index | -0.518 | 0.083 | 0.253 | 0.727 | -0.558 |
| 乡土植物比例The proportion of native plants | 0.614 | -0.937** | -0.941** | -0.510 | -0.454 |
| *表示P < 0.05, **表示P < 0.01, 表中数值代表相关系数平方值 | |||||
将城市公园景观类型中的林地、草地、人工表面、水体的景观格局指数与植物群落结构特征指数作相关性分析, 筛选出显著相关的指标。
(1) 草地景观格局指数与植物群落结构特征指数的相关性分析
从表 11中可以看出, 公园中草地景观的平均分维数与草本植物的Pielou指数呈显著负相关(P < 0.05);草地景观平均斑块面积、平均回转半径与灌木物种丰富度呈显著正相关、与灌木乡土植物种数呈显著负相关(P < 0.05)。草地景观最大斑块指数与乔木植物Shannon-Wiener指数、Pielou指数、Simpson指数呈显著负相关关系(P < 0.05)。
| 植物群落结构特征指数 Plant community characteristic index |
平均分维数 FRAC_MN |
平均斑块面积 AREA_MN/hm2 |
平均回转半径 GYRATE_MN/m |
最大斑块指数 LPI |
| 物种丰富度Species richness | 0.815* S | 0.887* S | ||
| Shannon-Wiener指数 Shannon-Wiener index |
-0.880* T | |||
| Pielou指数Pielou index | -0.842* H | -0.957** T | ||
| Simpson指数Simpson index | -0.816* T | -0.978** T | ||
| 乡土植物种数Native plant species | -0.889* S | -0.941** S | ||
| H:草本Herb;S:灌木Shrub;T:乔木Tree | ||||
(2) 林地景观格局指数与植物群落结构特征指数的相关性分析
从表 12中可以看出, 公园中林地景观平均斑块面积与草本乡土植物占比呈显著正相关(P < 0.05);林地周长面积比与草本Shannon-Wiener指数、Pielou指数、Simpson指数、乡土植物占比均呈显著负相关(P < 0.05)。林地周长面积分维数与草本物种丰富度呈显著负相关关系(P < 0.05)。林地景观斑块数量、斑块密度与灌木植物Shannon-Wiener指数、Pielou指数、Simpson指数均呈显著负相关关系(P < 0.05)。林地最大斑块指数与灌木Simpson指数呈显著正相关(P < 0.05)。
| 植物群落结构特征指数 Plant community characteristic index |
平均斑块面积 AREA_MN/hm2 |
周长面积比 PARA |
周长面积分维数 PAFRAC |
斑块数量 NP |
斑块密度 PD/(n/hm2) |
最大斑块指数 LPI |
| 物种丰富度Species richness | -0.839* H | |||||
| Shannon-Wiener指数 Shannon-Wiener index |
-0.922** H | -0.839* S | -0.880* S | |||
| Pielou指数Pielou index | -0.985** H | -0.838* S | -0.869* S | |||
| Simpson指数Simpson index | -0.968** H | -0.834* S | -0.882* S | 0.841* S | ||
| 乡土植物占比 The proportion of native plants |
0.865* H | -0.849* H |
(3) 水体景观格局指数与灌木植物群落结构特征指数的相关性分析
表 13中, 水体景观格局指数仅与灌木物种多样性呈显著相关关系。水体景观的斑块数量、斑块密度、最大斑块指数与灌木植物Shannon-Wiener指数呈显著负相关(P < 0.05)。水体周长面积比与灌木Shannon-Wiener指数、Simpson指数呈显著负相关(P < 0.05)。水体周长面积分维数与乡土植物占比呈显著负相关关系(P < 0.01)。
| 植物群落结构特征指数 Plant community characteristic indexes |
斑块数量 NP |
斑块密度 PD/(n/hm2) |
最大斑块指数 LPI |
周长面积比 PARA |
周长面积分维数 PAFRAC |
| 物种丰富度Species richness | |||||
| Shannon-Wiener指数Shannon-Wiener index | -0.813* S | -0.933** S | -0.927** S | -0.887* S | |
| Simpson指数Simpson index | -0.911* S | -0.827* S | |||
| 乡土植物占比Proportion of native plants | -0.958** S |
TS-4和H-1分别是公园中最常见的乔灌和草本群落类型, 在人工型公园中占比较大, 是人工养护下形成的植物群落。乔灌群落中主要物种为榕树、龙船花等乡土植物, 具有较好的适应性, 其中多数植物具有较好的遮荫、空气净化等生态功能和观赏效果[35]。草本中的结缕草、地毯草, 作为常见的草坪草具有较多优点, 例如耐践踏、寿命长, 具有广适性[36]。因此, 良好的景观效果、生态功能、人工管理的低维护性, 和乡土植物的适应性可能是该类植物群落分布较为普遍的原因。同时, 乡土植物较强的适应能力有益于构建稳定的植物群落, 发挥其生态功能。
公园灌木植物种类丰富度较低, 是其结构与功能特点决定的。灌木通常被用于界定景观空间、丰富植物层次, 呈条状或片状分布[37]。然而其数量丰富, 植株之间联系紧密, 群落结构复杂, 具有较高的适应性[38]。另外, 其具有耐修剪、抗风、生长力强等特点, 也促使灌木群落稳定性较高。可见, 用较少的灌木物种就能实现较高的稳定性。因此, 建议在公园植物造景初期就要重视对灌木的运用, 尤其是一些生态本底比较脆弱的公园, 以提高植物群落的稳定性和低维护性。
近海公园总体乡土植物种数占比较高, 但以草本植物为主, 乔灌木植物比例不高。原因可能与深圳对城市公园绿化的经济投入较大[39], 引入了大量如红花玉蕊、象脚丝兰等形态独特、观赏效果较好的外来木本植物有关。乡土乔灌植物的应用有利于缓解同质化效应, 对城市生态系统功能的发挥非常重要[40-41]。但城市人口众多, 人为活动频繁而密集, 不仅需要考虑公园植物群落的生态功能, 也要考虑其美观性、康养效果等其他功能。是否有必要增加乡土乔灌木的比例, 以及维持多大的比例, 需要权衡植物群落多方面的功能与需求, 同时要结合动物生存、人类使用等多重生态系统问题作具体的判断, 有待未来进行更深入的研究。
近海公园的乔灌植物群落能够较好地反映深圳的地域特色, 其指示种多为具有热带性的树种, 如榕树、红花羊蹄甲、小叶榄仁等。值得注意的是, 草本群落的指示种中, 出现了1级入侵物种喜旱莲子草, 其繁殖能力强, 对农田、水域有极大的危害[42]。也发现了入侵等级为5级的地毯草[42], 其入侵危害不明显。因此, 对具有较强入侵特性或潜力的物种进行持续性的调查与防治十分必要[43], 这对城市生态安全具有重要意义。
3.2 近海公园植物多样性与稳定性的关系探讨对于物种多样性与群落稳定性之间的关系长期以来存在较多争议。尽管物种多样性的提高确实有利于构建更为稳定的群落[44], 但越来越多的研究者提出二者的关系不是简单的线性关系[45]。基于研究结果, 将近海公园物种多样性与稳定性数值进行耦合分析, 比较他们在各个公园中的排序与分布规律。草本物种丰富度最高的是荔枝公园, 而稳定性最高的是洪湖公园。此外, 香蜜公园中乔木物种丰富度偏低, 稳定性却最高。因此, 二者并没有明显的大小关系。很可能是因为, 物种多样性与物种的功能群、功能冗余有关[46], 这些因素都可能导致群落稳定性的变化。未来可能还需要对功能多样性、功能性状进行测量, 以便进一步探讨二者的关系。
3.3 公园景观结构对生物多样性保护的启示近海公园乔木物种多样性只与草地最大斑块指数呈负相关。可能是因为在营造满足人为活动、集散需求的开敞空间时, 公园入口、水体等景观周围, 通常分布着许多大面积的草坪, 而不是物种较丰富的乔木群落景观[47]。因此在公园面积一定的情况下, 草地景观面积越大, 乔木植物多样性可能会相对减小。
从公园整体景观格局来看, 斑块密度和林地面积比、林地最大斑块面积比呈显著负相关。公园整体景观破碎度的增加容易导致林地景观破碎度增加, 使得林地核心斑块更容易缩小, 亚种群间的隔离程度增大, 生态流变弱, 从而加快了林地面积的丧失[48]。公园人工表面比例与乡土植物比例呈显著负相关。公园中的人工表面比例增加往往需要牺牲更多原有的绿地生境, 使得这些生境中残存的乡土植物栖息地面积减小或丧失[49], 导致其物种数降低。因此, 在公园设计和改造中, 要更加注重对乡土植物的保护。
植物群落是城市公园中发挥生态作用的主体, 植物群落结构与其景观分布格局对公园中的昆虫、鸟类等动物多样性也具有重要影响[50]。有研究证实, 公园林地最大斑块指数的增加有利于提高鸟类物种数[51-52], 大面积的公园绿地能维持大量的昆虫与土壤动物[53-54]。城市公园的景观组成与分布是公园的景观结构, 其植物群落多样性可看作生物多样性保护功能的潜力指标。基于结构与功能之间的关系, 在了解公园景观格局与植物多样性的相关性规律基础上, 可对城市植物配置进行优化, 这对公园生物多样性的维持和保护具有重要意义。
4 结论深圳近海公园植物调查共记录到植物108科310属471种, 其植物种类丰富, 物种多样性较高。植物群落的指示种多数具有热带性质, 公园中灌木植物种数低于乔木和草本植物, 但群落稳定性较高。半自然型和人工型近海公园相比较, 各类植物中以灌木植物相似性最低。公园林地面积比、林地最大斑块面积与整体景观破碎度呈显著负相关(P < 0.05), 公园景观破碎度的增大可能会导致植被景观面积减小。建议在公园植物群落的构建中, 重视灌木植物的运用, 有助于改善群落稳定性。同时在公园景观格局上, 应尽可能保留大面积的核心植被生态斑块, 更好地发挥其生态功能。
| [1] |
Husté A, Boulinier T. Determinants of local extinction and turnover rates in urban bird communities. Ecological Applications, 2007, 17(1): 168-180. DOI:10.1890/1051-0761(2007)017[0168:DOLEAT]2.0.CO;2 |
| [2] |
Dai X K, Zhu R, Jia P, Shen L, Li S P, Hua Z S, Li M G, Li J T. Factors governing the naturalization-to-invasion transition of exotic plants in Shenzhen, China. Applied Ecology and Environmental Research, 2016, 14(3): 637-677. DOI:10.15666/aeer/1403_637677 |
| [3] |
Sun B, Pan X, Han L B, Fei Y J. Selection of waterlogging-tolerant and water purification herbaceous plants for the construction of a sponge city in Shenzhen, China. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2020, 48(2): 1043-1056. DOI:10.15835/nbha48211908 |
| [4] |
蒙敬杭. 深圳建设国际一流城市绿化的缘由和路径. 风景园林, 2012(3): 172-172. DOI:10.3969/j.issn.1673-1530.2012.03.040 |
| [5] |
苏泳娴, 黄光庆, 陈修治, 陈水森, 李智山. 城市绿地的生态环境效应研究进展. 生态学报, 2011, 31(23): 7287-7300. |
| [6] |
Nagendra H, Gopal D. Tree diversity, distribution, history and change in urban parks: studies in Bangalore, India. Urban Ecosystems, 2011, 14(2): 211-223. DOI:10.1007/s11252-010-0148-1 |
| [7] |
Figueroa J A, Castro S A, Reyes M, Teillier S. Urban park area and age determine the richness of native and exotic plants in parks of a Latin American city: Santiago as a case study. Urban Ecosystems, 2018, 21(4): 645-655. DOI:10.1007/s11252-018-0743-0 |
| [8] |
Baró F, Haase D, Gómez-Baggethun E, Frantzeskaki N. Mismatches between ecosystem services supply and demand in urban areas: A quantitative assessment in five European cities. Ecological Indicators, 2015, 55: 146-158. DOI:10.1016/j.ecolind.2015.03.013 |
| [9] |
郝晟, 王春连, 林浩文. 城市湿地公园生物多样性设计与评估——以六盘水明湖国家湿地公园为例. 生态学报, 2019, 39(16): 5967-5977. |
| [10] |
王娟, 马履一, 王新杰, 刘俊, 张文娟, 万威. 北京城区公园绿地景观格局研究. 西北林学院学报, 2010, 25(4): 195-199. |
| [11] |
付晖, 方纪华, 许先升, 付广. 海口市中心城区公共绿地景观格局分析. 西北林学院学报, 2014, 29(6): 260-265. DOI:10.3969/j.issn.1001-7461.2014.06.50 |
| [12] |
Cheng X Y, Wei B S, Chen G J, Li J X, Song C H. Influence of park size and its surrounding urban landscape patterns on the park cooling effect. Journal of Urban Planning and Development, 2015, 141(3): A4014002.1-A4014002.10. |
| [13] |
Cornelis J, Hermy M. Biodiversity relationships in urban and suburban parks in Flanders. Landscape and Urban Planning, 2004, 69(4): 385-401. DOI:10.1016/j.landurbplan.2003.10.038 |
| [14] |
Penteado H M. Assessing the effects of applying landscape ecological spatial concepts on future habitat quantity and quality in an urbanizing landscape. Landscape Ecology, 2013, 28(10): 1909-1921. DOI:10.1007/s10980-013-9940-7 |
| [15] |
刘瑞雪, 许晓雪, 陈龙清. 基于使用行为的城市公园植物景观空间调查研究——以深圳湾滨海公园为例. 中国园林, 2019, 35(4): 123-128. |
| [16] |
雷金睿, 宋希强, 何荣晓. 滨海城市公园植物物种多样性比较——以海口市为例. 生态学杂志, 2016, 35(1): 118-124. |
| [17] |
林明太, 孙虎, 郭斌. 福建旅游型海岛景观格局变化——以妈祖圣地湄洲岛为例. 生态学杂志, 2010, 29(7): 1414-1419. |
| [18] |
Czortek P, Pielech R. Surrounding landscape influences functional diversity of plant species in urban parks. Urban Forestry & Urban Greening, 2020, 47: 126525. |
| [19] |
胡宏友, 肖亮嫦, 张万旗, 刘俊伟, 董克钻, 李学梅. 厦门城市公园景观格局与植物群落结构的相关性. 生态学杂志, 2010, 29(11): 2229-2234. |
| [20] |
彭羽, 刘雪华. 城市化对植物多样性影响的研究进展. 生物多样性, 2007, 15(5): 558-562. DOI:10.3321/j.issn:1005-0094.2007.05.014 |
| [21] |
深圳市气象局(台). 深圳市社会发展和现代化统计监测年报(2018年全年). (2019-05-14)[2021-05-26]. http://weather.sz.gov.cn/xingxigongkai/tongjishuju/shujujiedu/content/post_3569348.html.
|
| [22] |
赵娟娟, 宋晨晨, 刘时彦. 城市植物种类构成的特征分析——以厦门市为例. 西南大学学报: 自然科学版, 2018, 40(7): 1-8. |
| [23] |
谢世林, 逯非, 曹垒, 周伟奇, 欧阳志云. 北京城区公园景观格局对夏季鸟类群落的影响. 景观设计学, 2016, 4(3): 10-21. |
| [24] |
王静, 周伟奇, 许开鹏, 颜景理. 京津冀地区城市化对植被覆盖度及景观格局的影响. 生态学报, 2017, 37(21): 7019-7029. |
| [25] |
布仁仓, 胡远满, 常禹, 李秀珍, 贺红士. 景观指数之间的相关分析. 生态学报, 2005, 25(10): 2764-2775. DOI:10.3321/j.issn:1000-0933.2005.10.044 |
| [26] |
Wittig R, Becker U. The spontaneous flora around street trees in cities-A striking example for the worldwide homogenization of the flora of urban habitats. Flora-Morphology, Distribution, Functional Ecology of Plants, 2010, 205(10): 704-709. DOI:10.1016/j.flora.2009.09.001 |
| [27] |
张峰, 张金屯, 上官铁梁. 历山自然保护区猪尾沟森林群落植物多样性研究. 植物生态学报, 2002, 26(S1): 46-51. |
| [28] |
尹锴, 崔胜辉, 石龙宇, 吝涛, 郭青海, 吕静. 人为干扰对城市森林灌草层植物多样性的影响——以厦门市为. 生态学报, 2009, 29(02): 563-572. DOI:10.3321/j.issn:1000-0933.2009.02.002 |
| [29] |
Wang C Y, Wu B D, Jiang K, Zhou J W, Liu J, Lv Y N. Canada goldenrod invasion cause significant shifts in the taxonomic diversity and community stability of plant communities in heterogeneous landscapes in urban ecosystems in East China. Ecological Engineering, 2019, 127: 504-509. DOI:10.1016/j.ecoleng.2018.10.002 |
| [30] |
张景慧, 黄永梅. 生物多样性与稳定性机制研究进展. 生态学报, 2016, 36(13): 3859-3870. |
| [31] |
Godron M, Daget P, Poissonet J, Poissonet P. Some aspects of heterogeneity in grasslands of Cantal//Patil G P, Pielou E C, Waters W E, eds. Statistical Ecology: Volume 3: Many Species Populations, Ecosystems, and Systems Analysis. University Park, PA: Pennsylvania State University Press, 1972: 397-145.
|
| [32] |
胡寒剑. 杭州西湖湖西景区湿地植物群落研究[D]. 杭州: 浙江农林大学, 2015.
|
| [33] |
刘瑞雪, 陈龙清. 水淹干扰对水库水滨带植物群落稳定性与种间关系的影响. 生态学报, 2021, 41(16): 1-14. (2021-05-21). https://kns.cnki.net/KCMS/detail/detail.aspx2dbcode=CJFQ&dbname=CAPJLAST&filename=STXB20210520001&v=MDQ3MDlXTTBDTEw3UjdxZFp1Wm5GaTNsVjdyS0lWbz1Oam5UYkxHNEhORE1xbzFGWk9zT1l3OU16bVJuNmo1N1QzZmxx.
|
| [34] |
曾晓阳. 成都市城市森林的近自然植物群落配置模式研究[D]. 成都: 四川农业大学, 2009.
|
| [35] |
黄穗昇, 周劲松, 陈红锋. 深圳乡土地被植物调查及园林应用分析. 中国园林, 2007, 23(9): 81-84. DOI:10.3969/j.issn.1000-6664.2007.09.024 |
| [36] |
李军. 城市植物景观恢复技术与质量评价体系研究——以珠三角的广州、深圳、珠海为例[D]. 长沙: 中南林业科技大学, 2005.
|
| [37] |
杭夏子, 袁喆, 翁殊斐. 广州市公园园林灌木资源及其景观特色调查与分析. 中国园林, 2014, 30(4): 104-107. |
| [38] |
吴瑾, 孙斌, 刘爱容, 于法钦, 钟美晶. 演替视角下深圳近自然社区绿道植物配置实践分析. 生态环境学报, 2015, 24(9): 1461-1465. |
| [39] |
孙逊, 沈华山. 深圳香蜜公园植物保护与景观营造. 中国园林, 2018, 34(S2): 146-150. |
| [40] |
达良俊. 基于本土生物多样性恢复的近自然城市生命地标构建理念及其在上海的实践. 中国园林, 2021, 37(5): 20-24. |
| [41] |
Zerbe S, Maurer U, Schmitz S, Sukopp H. Biodiversity in Berlin and its potential for nature conservation. Landscape and Urban Planning, 2003, 62(3): 139-148. DOI:10.1016/S0169-2046(02)00145-7 |
| [42] |
闫小玲, 刘全儒, 寿海洋, 曾宪锋, 张勇, 陈丽, 刘演, 马海英, 齐淑艳, 马金双. 中国外来入侵植物的等级划分与地理分布格局分析. 生物多样性, 2014, 22(5): 667-676. |
| [43] |
邵志芳, 赵厚本, 邱少松, 杨义标, 彭少麟, 陆宠芳, 陈卓全. 深圳市主要外来入侵植物调查及治理状况. 生态环境, 2006, 15(3): 587-593. DOI:10.3969/j.issn.1674-5906.2006.03.029 |
| [44] |
McCann K S. The diversity-stability debate. Nature, 2000, 405(6783): 228-233. DOI:10.1038/35012234 |
| [45] |
Jiang L, Pu Z C. Different effects of species diversity on temporal stability in single-trophic and multitrophic communities. The American Naturalist, 2009, 174(5): 651-659. DOI:10.1086/605961 |
| [46] |
Wang H D, Zhang L L, Zhu Z H. Effects of clipping and fertilizing on the relationships between species diversity and ecosystem functioning and mechanisms of community stability in alpine meadow. Chinese Journal of Plant Ecology, 2013, 37(4): 279-295. DOI:10.3724/SP.J.1258.2013.00028 |
| [47] |
White J G, Antos M J, Fitzsimons J A, Palmer G C. Non-uniform bird assemblages in urban environments: the influence of streetscape vegetation. Landscape and Urban Planning, 2005, 71(2/4): 123-135. |
| [48] |
Craven D, Eisenhauer N, Pearse W D, Hautier Y, Isbell F, Roscher C, Bahn M, Beierkuhnlein C, Bönisch G, Buchmann N, Byun C, Catford J A, Cerabolini B E L, Cornelissen J H C, Craine J M, De Luca E, Ebeling A, Griffin J N, Hector A, Hines J, Jentsch A, Kattge J, Kreyling J, Lanta V, Lemoine N, Meyer S T, Minden V, Onipchenko V, Polley H W, Reich P B, Van Ruijven J, Schamp Schamp B, Smith M D, Soudzilovskaia N A, Tilman D, Weigelt A, Wilsey B, Manning P. Multiple facets of biodiversity drive the diversity-stability relationship. Nature Ecology & Evolution, 2018, 2(10): 1579-1587. |
| [49] |
Yang F P, Ignatieva M, Wissman J, Ahrné K, Zhang S X, Zhu S Y. Relationships between multi-scale factors, plant and pollinator diversity, and composition of park lawns and other herbaceous vegetation in a fast growing megacity of China. Landscape and Urban Planning, 2019, 185: 117-126. DOI:10.1016/j.landurbplan.2019.02.003 |
| [50] |
Han Y W, Kang W M, Thorne J, Song Y. Modeling the effects of landscape patterns of current forests on the habitat quality of historical remnants in a highly urbanized area. Urban Forestry & Urban Greening, 2019, 41: 354-363. |
| [51] |
Canedoli C, Manenti R, Padoa-Schioppa E. Birds biodiversity in urban and periurban forests: environmental determinants at local and landscape scales. Urban Ecosystems, 2018, 21(4): 779-793. DOI:10.1007/s11252-018-0757-7 |
| [52] |
王本耀, 王小明, 王天厚, 丁由中. 上海闵行区园林鸟类群落嵌套结构. 生态学报, 2012, 32(009): 2788-2795. |
| [53] |
Milano V, Maisto G, Baldantoni D, Bellino A, Bernard C, Croce A, Dubs F, Strumia S, Cortet J. The effect of urban park landscapes on soil Collembola diversity: A Mediterranean case study. Landscape and Urban Planning, 2018, 180: 135-147. DOI:10.1016/j.landurbplan.2018.08.008 |
| [54] |
Huang Y, Zhao Y Z, Li S H, Von Gadow K. The Effects of habitat area, vegetation structure and insect richness on breeding bird populations in Beijing urban parks. Urban Forestry & Urban Greening, 2015, 14(4): 1027-1039. |