 2021, Vol. 41
2021, Vol. 41文章信息
- 刘丹丹, 武海涛, 芦康乐, 杨萌尧, 管强, 赵文元, 邵乐夫
- LIU Dandan, WU Haitao, LU Kangle, YANG Mengyao, GUAN Qiang, ZHAO Wenyuan, SHAO Lefu
- 空间和环境因子对黄河口自然和淡水恢复湿地底栖动物群落的差异影响
- Effects of spatial and environmental factors on benthic invertebrate communities in natural and freshwater restored wetlands of the Yellow River Delta
- 生态学报. 2021, 41(17): 6893-6903
- Acta Ecologica Sinica. 2021, 41(17): 6893-6903
- http://dx.doi.org/10.5846/stxb202009052312
-
文章历史
- 收稿日期: 2020-09-05
- 网络出版日期: 2021-06-11





2. 东北林业大学野生动物与自然保护地学院, 哈尔滨 150040;
3. 中国科学院大学, 北京 100049;
4. 辽宁省湿地保护管理中心, 辽阳 111010
2. College of Wildlife and Protected Area, Northeast Forestry University, Harbin 150040, China;
3. University of Chinese Academy of Science, Beijing 100049, China;
4. Liaoning Municipal Wetland Protection and Management Center, Liaoyang 111010, China
群落构建机制是群落生态学研究的核心内容[1-2]。其中, 生态位理论(确定性过程)和中性理论(随机性过程)是目前群落构建机制最重要的两种观点[3], 分别强调了环境(生境特征和生物间相互作用)和空间过程(扩散限制)在群落构建中的重要性[4]。环境过滤和扩散作用通常会导致群落结构发生重大变化[5-6], 且越来越多的证据表明两者对群落构建过程均具有不可忽视的作用[7-8]。因此, 判定环境和空间因子的相对重要性对生物群落中物种分布和共存模式的理解至关重要[9-10]。
由于气候、区域间的生态模式及其形成机制的差异, 不同生态系统驱动底栖动物群落构建过程受到了生态学家的极大关注[11]。在陆地、淡水和海洋生态系统中, 空间和环境梯度变化对群落组成的影响已得到广泛的研究[12-13]。有研究发现湖泊的面积、深度、pH值、营养物质和生物相互作用(如捕食)等是影响底栖动物群落结构的关键环境因素, 也由于扩散和隔离的阻碍, 导致湖泊的空间位置可能影响了物种的定殖和共存[14]。河口湿地中, 底栖动物直接参与生态系统的养分循环和能量代谢, 而且对外界环境变化和干扰反应敏感, 经常被用作指示生物进行研究[15-16]。目前, 底栖动物群落构建研究主要集中在湖泊、溪流等生态系统中, 在河口湿地生态系统中, 有关于环境和空间因子对底栖动物群落的影响研究有限。
河口湿地处于陆地和海洋的交接过渡地带, 具有复杂多样的空间格局, 独特的构造过程以及不同于湖泊、河流和海洋的生物地球化学循环和生物群落[17-20]。黄河三角洲河口湿地是中国暖温带地区保存最广阔、最完善、最年轻的湿地, 其地理位置独特, 生态类型多样, 生物多样性资源丰富[21-23]。近年来, 受自然因素和人类活动的多重影响, 黄河三角洲河口湿地面积减少和退化, 生物多样性受损, 生态系统结构功能面临严峻挑战和威胁[24]。为了抑制湿地生态系统进一步退化, 保护区管理部门对退化湿地进行了淡水补水恢复工程, 使得湿地退化和生物多样性降低得到一定的改善。然而, 蓄水堤坝的建设阻碍了恢复湿地与海洋之间的水文连通, 同时水体理化性质的改变导致水生生物群落结构发生变化[18]。因此, 评估环境和空间因子的调控作用对保护湿地生物多样性具有重要意义。本文以黄河三角洲自然保护区为研究区域, 通过对比分析淡水恢复湿地和自然湿地中底栖动物群落结构, 探究空间因子和环境过程对底栖动物群落变化的相对贡献及关键环境因子, 为提高黄河三角洲底栖动物的生物多样性提供线索。
1 材料和方法 1.1 研究区域概况黄河三角洲位于渤海湾南岸和莱州湾西岸之间(37°34.768′—38°12.310′N, 118°32.981′—119°20.450′E)。受欧亚大陆和太平洋的共同影响, 属暖温带半湿润大陆性季风气候。年平均气温在11.7—12.6℃之间, 最高气温一般出现在7月份, 年平均降水量551.6mm。自2002年以来, 黄河三角洲自然保护区管理局对退化的河口湿地进行了大规模的淡水补给工程, 重塑湿地水文情势[18, 25]。恢复措施包括修建堤坝、建造蓄水池, 每年6月至7月通过现行的清水沟流路将黄河水引入退化湿地[17, 21, 24]。该工程成功的扩大了芦苇湿地的面积, 改善了水盐条件, 增加了黄河三角洲湿地的生物多样性[26]。

|
| 图 1 黄河三角洲淡水恢复湿地和自然湿地采样点分布图 Fig. 1 Sample sites of freshwater restored wetlands and natural wetlands in the Yellow River Delta |
底栖动物采用D型网(1mm网眼、直径35cm), 从水体向岸边水平拖动1m并进行四次重复采集底栖动物[27]。混合四个重复样品为一个采样点的样品, 用95%酒精保存, 贴上标签带回实验室进行分类和鉴定, 并统计1m×0.35m面积内的底栖动物数量。底栖动物鉴定检索使用《中国经济动物志: 淡水软体动物》[28]、《中国海岸带大型底栖动物资源》[29]、《Aquatic Insects of China Useful for Monitoring Water Quality》[30]等将绝大部分物种鉴定到种或属, 样品在体式显微镜下统计数量。
采用便携式水质分析仪(YSI-556MPS, 美国)现场测定了采样点水体的pH值、电导率(Electrical conductivity, Cond)、盐度(Salinity, SAL)、氯离子(Cl-)、氧化还原电位(Oxidation-reduction potential, ORP), 并采集1L混合水样, 带回实验室参照《地表水环境质量标准》(GB3838—2002)中的方法, 测定水样中的总氮(Total nitrogen, TN)、总碳(Total carbon, TC)、总有机碳(Total organic carbon, TOC)、无机碳(Inorganic carbon, IC)、溶解有机碳(Dissolved organic carbon, DOC)、硫酸盐离子(SO42-)、碳酸盐离子(CO32-)、碳酸氢盐离子(HCO3-)。
1.3 数据分析底栖动物群落多样性指数采用Shannon-Weiner多样性指数(H′)、辛普森指数(Simpson index, λ)和Pielou均匀度指数(Js)进行分析, 计算公式如下:
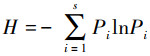
|
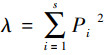
|
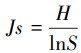
|
式中, H′为多样性指数, Pi为第i个种的个体数占群落总个体数的比率, S为类群数;个体数量占全部捕获量的10%以上为优势类群, 1%—10%为常见类群, 小于1%为稀有类群, 文章中将优势类群和常见类群归为主要类群。底栖动物群落多样性指数通过R软件中“picante”包完成。
采用每个样点的物种丰度数据进行非度量多维标度排序(Non-matric multidimentional scaling, NMDS)以识别不同湿地底栖动物群落结构空间分布的差异性;并通过相似性分析(Analysis of similarities, ANOSIM)进行显著性检验。对淡水恢复湿地和自然湿地的环境变量进行了对数转换, Kolmogorov-Smirnov检验表明符合正态分布;使用独立样本T检验进行环境数据差异性分析。NMDS和ANOSIM分别通过R软件中vegan包中的“metaMDS”函数和“anosim”函数实现, Kolmogorov-Smirnov检验和T检验采用SPSS Statistics Version 22完成。
采用Mantel检验和偏Mantel检验底栖动物群落物种组成距离、地理距离和环境距离之间的Spearman秩相关系数, 评估底栖动物群落与空间距离和环境距离的相关性。群落物种组成距离以物种丰度的Bray-curtis相异度进行测量, 样点间的地理距离利用采样点经纬度坐标计算, 环境距离是以不同采样点环境变量的欧几里得(Euclidean)距离测量。Mantel检验及偏Mantel检验通过R软件中的vegan包实现。
为确定底栖动物群落特征的关键环境因子, 首先对群落丰度数据进行除趋势对应DCA(Detrended correspondence analysis)分析, 结果显示“Axis lengths”最大值大于4, 因此选择典范对应CCA(Canonical correspondence analysis)分析, 解析底栖动物类群与环境因子的关系。为优化底栖动物群落数据, 剔除样点出现频度≤2的偶见种。环境数据(除pH外)和底栖动物群落数据都进行log(x+1) 转化。CCA排序中, 利用基于双终止准则(校正的R2)的前向选择(Forward selection)筛选显著环境变量, 其中所有环境变量都进行蒙特卡洛置换检验, 直到P≥0.05时终止选择。DCA和CCA通过R软件中vegan包实现。
对采样点的经纬度坐标进行邻体矩阵主坐标分析(Principal coordinates of neighboring matrices, PCNM)[31], 保留正相关的空间变量, 分析扩散作用对底栖动物群落结构的影响。PCNM分析主要是通过构建取样点间欧式距离矩阵, 模拟所有可能存在的空间结构;然后利用样点间最小连接值作为阈值对距离矩阵规模进行削减, 并计算削减后距离矩阵的主坐标分析(Principal coordinate analysis, PCoA), 得到n个由取样点-主坐标矩阵表示的空间变量;最后筛选正空间相关的特征变量, 作为解释变量用于后续变差分解分析[32-34]。PCNM分析通过R软件中“PCNM”包实现。
采用变差分解(Variation partitioning analysis, VPA)[35]评估环境变量和空间因子在解释群落结构组成中的相对贡献。将上述CCA和PCNM分析中前向选择筛选出的环境变量和正相关空间变量作为解释变量和转化后的底栖动物数据作为响应变量进行VPA分析;并通过偏冗余分析(partial RDA, pRDA)计算空间因子与环境变量单独部分解释量。VPA结果可以分为四个单独的部分[4, 36], 即纯环境解释量([E])、纯空间解释量([S])、纯环境和纯空间变量的共同解释量([E∩S])、未解释部分(1-[E∩S])。解释量的显著性使用蒙特卡罗置换检验实现(Monte carlo permutation test)。pRDA和VPA分析通过R软件中的vegan包实现。
2 结果与分析 2.1 环境因子特征独立样本T检验表明, 淡水恢复湿地和自然湿地的Cond、SAL、SO42-、Cl-、TN和pH等环境因子间均呈显著性差异(表 1, P < 0.05)。其中, Cond、SAL、SO42-和Cl-等表示盐度的环境指标差异极显著(P < 0.001), 这些参数值在淡水恢复湿地中相对较低;而HCO3-、CO32-、TOC、TC、IC、DOC和ORP等环境因子则在两类湿地之间的差异均不显著(P>0.05)。CCA分析结果显示, Cond、SAL和ORP是淡水恢复湿地的主要影响因子, pH和IC是自然湿地的主要影响因子(表 3)。由上可知, 盐度是区分两类湿地环境特征的重要指标。
| 环境因子 Environmental variables |
淡水恢复湿地 Freshwater restored wetlands |
自然湿地 Natural wetland |
t | P |
| 电导率Cond/(μS/cm) | 4.03±2.12 | 37.29±10.66 | -15.619 | < 0.001 |
| 盐度SAL/(g/L) | 2.27±1.34 | 26.25±7.53 | -16.826 | < 0.001 |
| 硫酸盐离子SO42-/(mg/L) | 222.08±82.95 | 705.42±469.66 | -5.548 | < 0.001 |
| 氯离子Cl-/(mg/L) | 250.12±94.04 | 8348.93±2019.12 | -28.48 | < 0.001 |
| 总氮TN/(mg/L) | 0.88±0.40 | 1.64±1.11 | -2.937 | 0.007 |
| pH | 8.12±0.35 | 7.72±0.24 | 3.479 | 0.002 |
| 碳酸氢盐离子HCO3-/(mg/L) | 145.60±37.73 | 173.09±40.55 | -1.761 | 0.089 |
| 碳酸盐离子CO32-/(mg/L) | 1.24±2.56 | 1.79±2.84 | -0.926 | 0.362 |
| 总有机碳TOC/(mg/L) | 22.63±14.83 | 17.87±8.45 | 1.23 | 0.229 |
| 总碳TC/(mg/L) | 50.50±18.20 | 42.64±9.36 | 1.344 | 0.19 |
| 无机碳IC/(mg/L) | 27.87±6.82 | 24.78±4.14 | 1.065 | 0.296 |
| 溶解有机碳DOC/(mg/L) | 17.58±11.17 | 15.59±7.64 | 0.566 | 0.576 |
| 氧化还原电位ORP/mv | 50.94±19.16 | 43.98±7.23 | 0.556 | 0.582 |
共采集鉴定底栖动物96个分类单元, 隶属于3门7纲20目59科。节肢动物门有3纲11目40科68种(占总物种数的70.83%), 其中昆虫纲占总分类单元的60.42%。软体动物门有2纲6目14科20种(21.51%), 由双壳纲和腹足纲组成, 分别占总分类单元数的7.29%和13.54%;环节动物门有2纲3目5科8种(8.33%), 由寡毛纲和多毛纲组成, 占总分类单元数的2.08%和6.25%。钉螺(Oncomelania hupensis)和中国毛虾(Acetes chinensis)为优势物种, 相对多度分别为21.4%和34.28%, 秀丽白虾(Chinese white prawn)和中国毛虾(Acetes chinensis)为出现频率最高的物种, 均分布在65.63%以上的采样点(表 2, 图 2)。
| 主要类群 Dominant species |
湿地类型Wetland types | F | P | |
| 淡水恢复湿地 Freshwater restored wetlands |
自然湿地 Natural wetlands |
|||
| 划蝽科的一种Corixidae sp. | 22.50±32.65 | 0.04±0.14 | 8.93 | 0.003 |
| 直突摇蚊亚科Orthocladiinae sp. | 3.44±6.20 | - | 10.47 | 0.001 |
| 蟌科的一种Agriocnemis sp. | 4.72±11.17 | - | 10.47 | 0.001 |
| 蟌科的一种Cercion sp. | 11.47±13.37 | - | 18.38 | 0.000 |
| 蟌科的一种Enallagma sp. | 3.25±3.48 | - | 13.34 | 0.000 |
| 大蜾蠃蜚Corophium major | 0.28±0.83 | 5.50±13.46 | 2.48 | 0.115 |
| 窄异跳钩虾Allorchestes angustus | 4.42±11.67 | 0.08±0.29 | 9.78 | 0.002 |
| 秀丽白虾Exopalaemon modestus | 15.44±16.08 | 0.67±1.37 | 18.81 | < 0.001 |
| 中国毛虾Acetes chinensis | 26.75±53.90 | 54.25±163.79 | 1.15 | 0.283 |
| 萝卜螺属Radix swinhoei | 8.94±15.67 | - | 11.86 | 0.001 |
| 狭萝卜螺Radix lagotis | 6.33±13.67 | - | 5.75 | 0.016 |
| 尖口圆扁螺Hippeutis cantori | 5.75±13.35 | - | 9.17 | 0.002 |
| 钉螺Oncomelania hupensis | - | 151.21±345.83 | 10.70 | 0.001 |
| 光滑狭口螺Stenothyra glabra | 1.89±3.84 | 6.33±8.24 | 0.82 | 0.365 |
| 丰富度指数(Richness index) | 15.50±6.90 | 6.17±3.76 | 14.09 | < 0.001 |
| 多样性指数(Shannon-Weiner index) | 2.56±0.82 | 1.14±0.93 | 11.76 | 0.001 |
| 辛普森指数(Simpson index) | 0.70±0.19 | 0.37±0.29 | 9.29 | 0.002 |
| 均匀性指数(Pielou evenness index) | 0.66±0.16 | 0.53±0.32 | 1.21 | 0.271 |

|
| 图 2 淡水恢复湿地和自然湿地底栖动物群落结构组成 Fig. 2 Benthic invertebrate community structure in freshwater restored wetlands and natural wetlands |
NMDS(Stress=0.08)分析结果显示淡水恢复湿地和自然湿地的底栖动物群落结构具有显著差异。另外, ANOSIM分析结果表明, 两类湿地的底栖动物群落间存在差异显著性(R=0.68, P=0.001, 图 3)。由表 2可知, 黄河三角洲淡水恢复湿地底栖动物的物种丰富度、Shannon-Weiner多样性指数和Simpson指数均明显高于自然湿地的物种组成, 且差异显著(P < 0.05);均匀度无明显差异。

|
| 图 3 淡水恢复湿地和自然湿地底栖动物的NMDS图 Fig. 3 NMDS of benthic invertebrate communities′ composition of the Yellow River Delta based on the Bray-Curtis similarity |
Mantel检验结果表明, 在淡水恢复湿地中, 底栖动物群落非相似性距离随环境距离的增加而增加(R=0.262, P=0.017), 与地理距离无显著相关性(R=0.106, P=0.103);而在自然湿地中, 底栖动物群落非相似性距离随地理距离的增加而增加(R=0.631, P=0.001), 与环境距离无显著相关性(图 4, 表 3;R=0.14, P=0.19)。

|
| 图 4 底栖动物群落非相似性距离与环境距离、地理距离的相关性散点图 Fig. 4 Scatter plots of relationships between community dissimilarity, geographic distance and environmental distance based on Spearman′s rank correlation “R”和“P”分别表示线性回归的相关系数和显著性 |
| 距离矩阵 Distance matrix |
淡水恢复湿地 Freshwater restored wetlands |
自然湿地 Natural wetlands |
|||
| R | P | R | P | ||
| 环境距离Environmental distance | 0.262 | 0.017 | 0.14 | 0.19 | |
| 地理距离Geographical distance | 0.106 | 0.103 | 0.631 | 0.001 | |
| 环境距离/地理距离Environmental distance/Geographical distance | 0.25 | 0.03 | 0.131 | 0.22 | |
| 地理距离/环境距离Geographical distance/Environmental distance | 0.067 | 0.196 | 0.63 | 0.001 | |
| 环境距离/地理距离表示, 在控制地理距离时, 底栖动物群落非相似性和环境距离的相关性;地理距离/环境距离表示, 在控制环境距离时, 底栖动物群落非相似性和地理距离的相关性 | |||||
偏Mantel检验进一步表明, 当排除地理距离因素影响时, 淡水恢复湿地底栖动物群落非相似性距离与环境距离仍具有显著相关性(R=0.25, P=0.03), 而与自然湿地的底栖动物群落相关性不显著(R=0.131, P=0.22);在排除环境距离的影响下, 淡水恢复湿地底栖动物群落非相似性距离与空间距离无显著关系(R=0.067, P=0.196), 而与自然湿地底栖动物群落显著相关(表 3;R=0.63, P=0.001)。偏Mantel检验结果与Mantel检验结果一致。然而, 样点间的地理距离不能代表与物种扩散相关的空间结构, 需加入空间变量进一步分析。
2.4 环境和空间因子的相对贡献根据CCA分析中的前向选择过程和PCNM分析筛选的正空间相关变量结果, 共有5个环境变量和3个空间变量与底栖动物群落的变化显著相关(表 4, P < 0.05)。其中, 3个环境变量(SAL、ORP和Cond)和2个空间变量(PCNM2和PCNM4)对淡水恢复湿地底栖动物群落变化有显著影响(P < 0.05);2个环境变量(pH和IC)和1个空间变量(PCNM1)对自然湿地底栖动物群落变化有显著影响(P < 0.05)。
| 湿地类型 Wetland types |
因子 Factors |
R2 | F | P |
| 淡水恢复湿地 Freshwater restored wetlands |
盐度SAL | 0.125 | 3.395 | 0.002 |
| 氧化还原电位ORP | 0.252 | 3.75 | 0.001 | |
| 电导率Cond | 0.3 | 2.013 | 0.032 | |
| PCNM2 | 0.135 | 2.491 | 0.03 | |
| PCNM4 | 0.122 | 2.469 | 0.015 | |
| 自然湿地 Natural wetlands |
pH | 0.405 | 3.236 | 0.008 |
| 无机碳IC | 0.264 | 4.859 | 0.001 | |
| PCNM1 | 0.287 | 2.076 | 0.045 | |
| PCNM: 主轴邻距法产生的空间变量Principal coordinates of neighbor matrices | ||||
VPA结果表明, 环境和空间变量共同解释了淡水恢复湿地43%和自然湿地50.6%的解释率(图 5)。在淡水恢复湿地, 纯环境变量[E](27.2%)的解释率大于纯空间变量[S](11%), 且均达到显著程度(F=3.39, P=0.001; F=2.35, P=0.003);在自然湿地中, 纯空间变量[S](17.7%)解释率大于纯环境变量[E](7.3%), 其中纯空间变量达到显著程度(F=4.22, P=0.001)。纯环境和纯空间变量共同解释([E∩S])4.8%的淡水恢复湿地和25.6%的自然湿地底栖动物群落的解释率。

|
| 图 5 环境变量和空间变量对底栖动物群落结构相对贡献韦恩图 Fig. 5 Venn diagram of variation partitioning, showing the contributions of spatial and environmental variables on the variation of freshwater restored wetlands and natural wetlands |
由于淡水释放造成的环境异质性, 导致底栖动物群落组成发生变化。NMDS显示, 黄河三角洲河口湿地底栖动物的空间分布表现出明显的区域性差异, 表明淡水恢复湿地和自然湿地的底栖动物群落结构具有显著差异。与天然湿地相比, 淡水恢复湿地具有更高的底栖动物多样性和丰富度, 这一结论与唐娜等[36]的研究结果一致。已有研究表明淡水输入可以使植物物种丰富度增加, 进一步恢复并扩大鸟类栖息生境, 使恢复湿地环境发生了极大的改变, 间接或直接影响底栖动物群落物种丰富度和多样性[36]。在淡水恢复湿地, 主要以半翅目、鞘翅目、双翅目和蜻蜓目等水生昆虫及腹足纲的底栖动物为主, 以往研究也表明鞘翅目和一些半翅目昆虫对盐度变化敏感[37]。这种差异可能是由于生境和环境因子发生的梯度变化导致的[15, 38]。
自黄河三角洲退化湿地生态补水项目实施以来, 淡水补给缓解了退化湿地日益加剧的水盐胁迫[21, 24, 39]。而蓄水堤坝的建设, 切断了淡水补给区域与海洋的水文连通性, 阻碍了生物群落的自然迁移, 栖息地生境发生极大改变。进一步的研究显示, 环境因子Cond、SAL、SO42-、Cl-、TN和pH在两类湿地之间的差异极显著。总氮是衡量水质营养状态的重要指标, 水体富营养化往往会导致底栖动物群落生物多样性的降低[39]。电导率是影响底栖动物的生存和群落结构稳定性主要因素[40]。本研究中, 底栖动物群落在自然湿地中的生物多样性和丰度小于恢复湿地, 这可能是由于随着Cond和TN的升高, 底栖动物对生境质量下降和水质污染敏感导致的[41]。在恢复湿地中, 水体pH呈弱碱性(7.77—8.47), 这与杨海强等[40]研究结果一致, 偏碱性水体可能更适合大多数底栖动物生存[42]。盐度是河口环境中的一个重要环境因素, 而且底栖动物对盐度变化敏感[43], 许多类群只适应生存于特定的盐度范围内[20]。我们的研究也表明了, 盐度是淡水恢复湿地和自然湿地底栖动物群落组成差异的关键因子。
3.2 环境及空间因子对底栖动物群落结构的相对调控作用空间因子和环境异质性对于理解生态学研究中群落的调控机制具有重要意义[5]。在本研究中, 环境过滤对淡水恢复湿地底栖动物群落起主导作用, 这可能是淡水恢复湿地的底栖动物群落对环境异质性具有更高的敏感性导致的。进一步分析发现空间和环境变量都能显著解释底栖动物群落变化。虽然环境过滤和扩散作用共同调控了淡水恢复湿地底栖动物群落结构的变化, 但这些因素的相对强度可能因物种生境差异和物种组成而有所不同[44-45]。在淡水恢复区域中, 共记录到62个稀有类群, 其中鞘翅目、双翅目、蜻蜓目、蜉蝣目和毛翅目等水生昆虫占稀有类群的45%。与优势类群和常见类群相比, 稀有类群的分布更依赖于栖息地生境[5, 46-47], 因此在淡水恢复区域中的底栖动物群落可能与环境变化的关系更为密切。另外, 黄河三角洲湿地生态恢复保护工程导致恢复湿地向海连通性遭到严重破坏。与空间更为开放的沿海水域不同, 在连通性被阻隔的水生系统中, 环境过滤作用对底栖动物群落的调控更为重要[48]。
空间因子对自然湿地底栖动物群落具有主要的调控作用。这与在浅滩[48]、潮间带区域[49-50]和潮下带区域[4]等水生系统对底栖动物群落的研究结果基本一致。与淡水恢复湿地不同, 自然湿地与海洋连接, 水文连通性良好, 因此空间因素对底栖动物群落的调控作用更为重要。然而, 环境和空间因子共同部分的解释率(25.6%)比纯空间因素(17.7%)略高, 说明环境和空间因子的相互作用在决定底栖动物群落结构中也起着至关重要的作用[51-52]。此外, 底栖动物群落的调控机制也可能是由栖息地的水文连通性和扩散潜力相互作用的结果[53]。由于恢复湿地被堤坝阻隔, 在低水文连通性的生境中, 物种通常扩散能力较弱。相反, 在自然湿地中物种往往具有较强的扩散能力, 但与环境变异相关性较弱。水文条件也是决定底栖动物群落结构的重要因素[54]。淡水恢复湿地与自然湿地的底栖动物群落结构不相似也可能是由于水动力条件不同导致的。鉴于水文连通性是驱动群落在生境间的定殖与扩散的主导因子[55], 关于补水后连通性的变化对底栖动物群落构建机制的影响有待进一步的研究。
| [1] |
May F, Huth A, Wiegand T. Moving beyond abundance distributions: neutral theory and spatial patterns in a tropical forest. Proceedings of the Royal Society B: Biological Sciences, 2015, 282(1802): 20141657. DOI:10.1098/rspb.2014.1657 |
| [2] |
Gibson N, Prober S, Meissner R, Van Leeuwen S. Implications of high species turnover on the South-western Australian sandplains. PLoS One, 2017, 12(2): e0172977. DOI:10.1371/journal.pone.0172977 |
| [3] |
Gravel D, Canham C D, Beaudet M, Messier C. Reconciling niche and neutrality: the continuum hypothesis. Ecology Letters, 2006, 9(4): 399-409. DOI:10.1111/j.1461-0248.2006.00884.x |
| [4] |
Rao Y Y, Cai L Z, Chen B W, Chen X W, Zheng L M, Lin S J. How do spatial and environmental factors shape the structure of a coastal macrobenthic community and meroplanktonic larvae cohort? Evidence from Daya Bay. Marine Pollution Bulletin, 2020, 157: 111242. DOI:10.1016/j.marpolbul.2020.111242 |
| [5] |
Heino J, Melo A S, Siqueira T, Soininen J, Valanko S, Bini L M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshwater Biology, 2015, 60(5): 845-869. DOI:10.1111/fwb.12533 |
| [6] |
黄建雄, 叶万辉, 练琚愉, 曹洪麟. 谱系结构、环境因子及空间因子对群落动态变化的影响. 科学通报, 2014, 59(35): 3471-3478. |
| [7] |
Leibold M A, McPeek M A. Coexistence of the niche and neutral perspectives in community ecology. Ecology, 2006, 87(6): 1399-1410. DOI:10.1890/0012-9658(2006)87[1399:COTNAN]2.0.CO;2 |
| [8] |
Ofiţeru I D, Lunn M, Curtis T P, Wells G F, Criddle C S, Francis C A, Sloan W T. Combined niche and neutral effects in a microbial wastewater treatment community. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(35): 15345-15350. DOI:10.1073/pnas.1000604107 |
| [9] |
Heino J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biological Reviews, 2013, 88(1): 166-178. DOI:10.1111/j.1469-185X.2012.00244.x |
| [10] |
Daniel J, Gleason J E, Cottenie K, Rooney R C. Stochastic and deterministic processes drive wetland community assembly across a gradient of environmental filtering. Oikos, 2019, 128(8): 1158-1169. DOI:10.1111/oik.05987 |
| [11] |
Tonkin J D, Merritt D M, Olden J D, Reynolds L V, Lytle D A. Flow regime alteration degrades ecological networks in riparian ecosystems. Nature Ecology & Evolution, 2018, 2(1): 86-93. |
| [12] |
Soininen J, McDonald R, Hillebrand H. The distance decay of similarity in ecological communities. Ecography, 2007, 30(1): 3-12. DOI:10.1111/j.0906-7590.2007.04817.x |
| [13] |
Heino J, Tolonen K T. Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnology and Oceanography, 2017, 62(6): 2431-2444. DOI:10.1002/lno.10577 |
| [14] |
Jackson D A, Peres-Neto P R, Olden J D. What controls who is where in freshwater fish communities-the roles of biotic, abiotic, and spatial factors. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(1): 157-170. |
| [15] |
李正飞, 王军, 谢志才, 丁城志, 蒋小明. 南腊河底栖动物多样性与环境因子的关系. 生态学杂志, 2016, 35(12): 3364-3373. |
| [16] |
芦康乐, 杨萌尧, 武海涛, 管强, 张科. 黄河三角洲芦苇湿地底栖无脊椎动物与环境因子的关系研究——以石油开采区与淡水补给区为例. 生态学报, 2020, 40(5): 1637-1649. |
| [17] |
Yang W, Li X X, Sun T, Pei J, Li M. Macrobenthos functional groups as indicators of ecological restoration in the northern part of China's Yellow River Delta Wetlands. Ecological Indicators, 2017, 82: 381-391. DOI:10.1016/j.ecolind.2017.06.057 |
| [18] |
Yang M Y, Lu K L, Batzer D P, Wu H T. Freshwater release into estuarine wetlands changes the structure of benthic invertebrate assemblages: a case study from the Yellow River Delta. Science of the Total Environment, 2019, 687: 752-758. DOI:10.1016/j.scitotenv.2019.06.154 |
| [19] |
解雪峰, 孙晓敏, 吴涛, 蒋国俊, 濮励杰, 项琦. 互花米草入侵对滨海湿地生态系统的影响研究进展. 应用生态学报, 2020, 31(06): 339-348. |
| [20] |
Cloern J E, Jassby A D, Schraga T S, Nejad E, Martin C. Ecosystem variability along the estuarine salinity gradient: examples from long-term study of San Francisco Bay. Limnology and Oceanography, 2017, 62(S1): S272-S291. |
| [21] |
裴俊, 杨薇, 王文燕. 淡水恢复工程对黄河三角洲湿地生态系统服务的影响. 北京师范大学学报: 自然科学版, 2018, 54(1): 104-112. |
| [22] |
靳宇弯, 杨薇, 孙涛, 李铭. 围填海活动对黄河三角洲滨海湿地生态系统的影响评估. 湿地科学, 2015, 13(6): 682-689. |
| [23] |
Yang W, Li X X, Sun T, Yang Z F, Li M. Habitat heterogeneity affects the efficacy of ecological restoration by freshwater releases in a recovering freshwater coastal wetland in China's Yellow River Delta. Ecological Engineering, 2017, 104: 1-12. DOI:10.1016/j.ecoleng.2017.04.007 |
| [24] |
杨薇, 裴俊, 李晓晓, 孙涛, 王文燕. 黄河三角洲退化湿地生态修复效果的系统评估及对策. 北京师范大学学报: 自然科学版, 2018, 54(1): 98-103. |
| [25] |
Chi Y, Zheng W, Shi H H, Sun J K, Fu Z Y. Spatial heterogeneity of estuarine wetland ecosystem health influenced by complex natural and anthropogenic factors. Science of the Total Environment, 2018, 634: 1445-1462. DOI:10.1016/j.scitotenv.2018.04.085 |
| [26] |
Li M, Yang W, Sun T, Jin Y W. Potential ecological risk of heavy metal contamination in sediments and macrobenthos in coastal wetlands induced by freshwater releases: a case study in the Yellow River Delta, China. Marine Pollution Bulletin, 2016, 103(1/2): 227-239. |
| [27] |
Wu H T, Guan Q, Lu X G, Batzer D P. Snail (Mollusca: Gastropoda) assemblages as indicators of ecological condition in freshwater wetlands of northeastern China. Ecological Indicators, 2017, 75: 203-209. DOI:10.1016/j.ecolind.2016.12.042 |
| [28] |
刘月英, 张文珍, 王跃先, 王恩义. 中国经济动物志: 淡水软体动物. 北京: 科学出版社, 1979.
|
| [29] |
李宝泉, 李新正, 陈琳琳, 王少青. 中国海岸带大型底栖动物资源. 北京: 科学出版社, 2019.
|
| [30] |
Morse J C, Yang L F, Tian L. Aquatic Insects of China Useful for Monitoring Water Quality. Nanjing: Hohai University Press, 1994.
|
| [31] |
Borcard D, Legendre P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecological Modelling, 2002, 153(1/2): 51-68. |
| [32] |
高梅香, 何萍, 孙新, 张雪萍, 吴东辉. 环境筛选、扩散限制和生物间相互作用在温带落叶阔叶林土壤跳虫群落构建中的作用. 科学通报, 2014, 59(24): 2426-2438. |
| [33] |
Borcard D, Gillet F, Legendre P. 数量生态学: R语言的应用. 赖江山, 译. 北京: 高等教育出版社, 2014.
|
| [34] |
Borcard D, Gillet F, Legendre P. 数量生态学: R语言的应用(第二版). 赖江山, 译. 北京: 高等教育出版社, 2020.
|
| [35] |
Peres-Neto P R, Legendre P, Dray S, Borcard D. Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology, 2006, 87(10): 2614-2625. DOI:10.1890/0012-9658(2006)87[2614:VPOSDM]2.0.CO;2 |
| [36] |
唐娜, 崔保山, 赵欣胜. 黄河三角洲芦苇湿地的恢复. 生态学报, 2006(08): 2616-2624. DOI:10.3321/j.issn:1000-0933.2006.08.026 |
| [37] |
Piscart C, Moreteau J C, Beisel J N. Biodiversity and structure of macroinvertebrate communities along a small permanent salinity gradient(Meurthe River, France). Hydrobiologia, 2005, 551(1): 227-236. DOI:10.1007/s10750-005-4463-0 |
| [38] |
Jiang X M, Xiong J, Song Z Y, Morse J C, Jones F C, Xie Z C. Is coarse taxonomy sufficient for detecting macroinvertebrate patterns in floodplain lakes?. Ecological Indicators, 2013, 27: 48-55. DOI:10.1016/j.ecolind.2012.11.015 |
| [39] |
殷旭旺, 韩洁, 王博涵, 金文, 杨璐, 陈海, 吴丹. 太子河流域底栖动物群落结构及其与环境因子的关系. 水产学杂志, 2017, 30(3): 40-44. DOI:10.3969/j.issn.1005-3832.2017.03.008 |
| [40] |
杨海强, 潘保柱, 朱朋辉, 蒋万祥, 蒋小明, 侯易明. 渭河干流和秦岭北麓典型支流底栖动物群落结构及水质生物评价. 湖泊科学, 2020, 32(6): 1793-1805. |
| [41] |
张又, 蔡永久, 张颖, 高俊峰, 李宽意. 河流大型底栖动物对环境压力的响应: 以太湖、巢湖流域为例. 湖泊科学, 2021, 33(1): 204-217. |
| [42] |
Buss D F, Baptista D F, Silveira MP, Nessimian J L, Dorvillé L F M. Influence of water chemistry and environmental degradation on macroinvertebrate assemblages in a river basin in south-east Brazil. Hydrobiologia, 2002, 481(1/3): 125-136. DOI:10.1023/A:1021281508709 |
| [43] |
李姗泽, 崔保山, 谢湉, 张树岩, 刘伟华, 付守强. 黄河三角洲沼泽中大型底栖动物的分布特征. 湿地科学, 2015, 13(06): 759-764. |
| [44] |
Heino J. Does dispersal ability affect the relative importance of environmental control and spatial structuring of littoral macroinvertebrate communities?. Oecologia, 2013, 171(4): 971-980. DOI:10.1007/s00442-012-2451-4 |
| [45] |
Heino J. Environmental heterogeneity, dispersal mode, and co-occurrence in stream macroinvertebrates. Ecology and Evolution, 2013, 3(2): 344-355. DOI:10.1002/ece3.470 |
| [46] |
Josefson A B. Species sorting of benthic invertebrates in a salinity gradient-importance of dispersal limitation. PLoS One, 2016, 11(12): e0168908. DOI:10.1371/journal.pone.0168908 |
| [47] |
Leibold M A, Holyoak M, Mouquet N, Amarasekare P, Chase J M, Hoopes M F, Holt R. D, Shurin J B, Law R, Tilman D, Loreau M, Gonzalez A. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters, 2004, 7(7): 601-613. DOI:10.1111/j.1461-0248.2004.00608.x |
| [48] |
Rodil I F, Lucena-Moya P, Jokinen H, Ollus V, Wennhage H, Villnäs A, Norkko A. The role of dispersal mode and habitat specialization for metacommunity structure of shallow beach invertebrates. PLoS One, 2017, 12(2): e0172160. DOI:10.1371/journal.pone.0172160 |
| [49] |
Checon H H, Amaral A C Z. Taxonomic sufficiency and the influence of rare species on variation partitioning analysis of a polychaete community. Marine Ecology, 2017, 38(1): e12384. DOI:10.1111/maec.12384 |
| [50] |
Corte G N, Goncalves-Souza T, Checon H H, Siegled E, Coleman R, Amaral A C Z. When time affects space: dispersal ability and extreme weather events determine metacommunity organization in marine sediments. Marine Environmental Research, 2018, 136: 139-152. DOI:10.1016/j.marenvres.2018.02.009 |
| [51] |
Cottenie K, Michels E, Nuytten N, De MeesterL. Zooplankton metacommunity structure: regional vs. local processes in highly interconnected ponds. Ecology, 2003, 84(4): 991-1000. DOI:10.1890/0012-9658(2003)084[0991:ZMSRVL]2.0.CO;2 |
| [52] |
Grönroos M, Heino J, Siqueira T, Landeiro V L, Kotanen J, Bini L M. Metacommunity structuring in stream networks: roles of dispersal mode, distance type, and regional environmental context. Ecology and Evolution, 2013, 3(13): 4473-4487. DOI:10.1002/ece3.834 |
| [53] |
Sarremejane R, Mykrä H, Bonada N, Aroviita J, Muotkä T. Habitat connectivity and dispersal ability drive the assembly mechanisms of macroinvertebrate communities in river networks. Freshwater Biology, 2017, 62(6): 1073-1082. DOI:10.1111/fwb.12926 |
| [54] |
Rennella A M, Quirós R. The effects of hydrology on plankton biomass in shallow lakes of the Pampa Plain. Hydrobiologia, 2006, 556(1): 181-191. DOI:10.1007/s10750-005-0318-y |
| [55] |
Obolewski K. Macrozoobenthos patterns along environmental gradients and hydrological connectivity of oxbow lakes. Ecological Engineering, 2011, 37(5): 796-805. DOI:10.1016/j.ecoleng.2010.06.037 |