 2021, Vol. 41
2021, Vol. 41文章信息
- 卢福浩, 沙衣班·吾布力, 刘深思, 徐贵青
- LU Fuhao, Shayiban·Wubuli, LIU Shensi, XU Guiqing
- 根深决定不同个体大小梭梭对夏季干旱生理响应的差异
- Rooting depth determined physiological response of Haloxylon ammodendron to summer drought
- 生态学报. 2021, 41(8): 3178-3189
- Acta Ecologica Sinica. 2021, 41(8): 3178-3189
- http://dx.doi.org/10.5846/stxb201909282038
-
文章历史
- 收稿日期: 2019-09-28
- 网络出版日期: 2021-02-24





2. 自治区林业生态监测总站 乌鲁木齐 830000;
3. 中国科学院大学, 北京 100049;
4. 中国科学院新疆生态与地理研究所, 荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011;
5. 中国科学院阜康荒漠生态实验站, 阜康 831505
2. Forestry Ecological Monitoring Station, Xinjiang Urumqi 830000, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Xinjiang Institute of Ecology and Geography, Chinese Academy Science, Urumqi 830011, China;
5. Fukang Station of Desert Ecology, Chinese Academy of Sciences, Fukang 831505, China
随着全球气候变暖加剧, 全球或局部降水格局(降水量、降水强度及降水的季节分配)将发生调整[1]。基于GCM模式的气候变化情景数据, 采用帕尔默干旱指数评估我国干旱事件发生的时空变化特征, 表明21世纪中后期我国将面临广泛的干旱化趋势;尤其是新疆的核心沙漠区和内蒙古西部的戈壁沙漠区[2]。因此, 荒漠区植被将受到更为频繁和持续的干旱胁迫, 导致植物生长不良, 甚至大面积死亡[3-4]。荒漠植被群落中的优势种或建群种, 是群落生产力或生物量的主要贡献者, 其大面积的死亡将会改变生态系统的碳水循环过程, 使生态系统从碳汇变成碳源, 进一步反馈到区域乃至全球气候, 最终导致荒漠植被生态系统的服务功能丧失[5-6], 产生一系列难以预见与逆转的后果。
在树木生长发育过程中, 其大小和结构复杂性都在增加, 不同年龄的个体对生境资源利用存在差异, 会导致其对资源可利用性变化产生不同的生理响应[7-9]。在资源丰富的生境(如热带雨林), 植物对资源的竞争一般是非对称的, 如光照通常优先被高大的植物所获取[10]。而在养分和水分不足的生境中, 如干旱荒漠区, 植物对地下资源竞争通常是大小对称的, 且竞争主要集中在根部;尤其在水分极度匮乏下, 植物对资源的竞争则是依赖个体大小的对称资源分割, 即: 植物可利用资源的数量与其个体大小成比例的增加[9-12]。这意味着, 对于干旱荒漠区植物而言, 在植物个体发育过程中, 伴随生物量分配变化发生的根系维度调整(根系大小、分布和根深)可能在决定植物的存活上发挥着重要作用[13]。
梭梭作为古尔班通古特沙漠的主要建群种, 长期适应干旱荒漠环境的过程中, 在生活史(花后生殖休眠)、生理(渗透调节)和形态结构(叶片退化成同化枝执行光合功能)等方面发展了一系列适应性策略[14-17]。同其他植物相比, 梭梭对水分条件的变化具有更强的缓冲和适应能力, 在维持其组成群落的结构和功能中处于主导地位, 具有着重要的生态价值[18]。然而近年在人类活动和气候变化影响下, 梭梭种群数量减少, 面积收缩[19];在种子萌发季节, 降雨模式的不利变化, 致使准噶尔盆地南缘梭梭种群幼苗补员受阻, 幼苗大量夭亡, 梭梭种群年龄结构普遍呈现衰退, 群落显现逆行演替特征[20]。
在全球变化背景下, 不同年龄或大小的梭梭对干旱胁迫的响应特征和适应机制存在什么区别?是什么因素主导着这种差异?相关研究不足, 限制了对自然更新过程中梭梭种群动态变化的认识。我们前期的研究表明, 梭梭的抗旱策略是将光合产物优先分配到地下生长根系以提高水分获取的能力[21-22]。对于幼树而言, 由于根系较浅且不发达, 所能利用到的土壤水分极其有限, 同成年植株相比, 其在水分获取中处于劣势。而在水分匮乏的旱季, 梭梭种群内不同大小个体的根系深度可能在植物水分可利用性上起重要作用。综上所述, 我们推测:不同大小梭梭根系深度在决定越夏生长期间的植物水分状况、生理性状表达起重要作用, 并支配不同大小个体梭梭对干旱胁迫的差异性响应。
1 试验地概况研究区位于古尔班通古特沙漠南缘之中的实验样地。该区属于温带大陆性荒漠气候, 冬季寒冷, 夏季干热, 昼夜温差较大。年均温度6.6℃, 年降水量不超过150 mm, 年潜在蒸发量2000 mm。降水季节分配均匀, 冬季积雪78.8%—92%转化为土壤水, 为春季荒漠植被生长提供了良好的水分条件[23]。实验样地主要以固定和半固定沙丘为主, 土壤以风沙土为主。
研究区主要植物种有梭梭(Haloxylon ammodendron)、白梭梭(Haloxylon persicum)、角果藜(Ceratocarpus arenarius)、对节刺(Horaninovia ulicina)、地白蒿(Artemisia terrae-albae)、盐生假木贼(Anabasis salsa)、沙蓬(Agriophyllum squarrosum)及多种短命、类短命植物[19]。梭梭是该研究区最为重要的建群种。由于梭梭存在假年轮现象, 难以通过年轮确定树龄, 因此以基茎表征梭梭个体大小[24]。选取未受人为因素干扰、分布相对均匀的梭梭种群作为实验小区。在小区内随机选取生长健康、没有受病原体影响、没有动物啃食迹象的梭梭。使用数字游标卡尺测量其基茎, 按照基茎大小, 划分为0—1 cm, 1—2 cm, 2—4 cm, 4—8 cm, >8 cm五组, 每组5株梭梭, 并挂小牌标记, 用于植物生理性状的测量;另选五组, 用于根系调查。其中0—1 cm组, 基茎(0.59±0.1) cm SE, 树高(0.36±0.1) m SE;1—2 cm组, 基茎(1.44±0.1) cm SE, 树高(0.64±0.1) m SE;2—4 cm组, 基茎(2.92±0.1) cm SE, 树高(1.12±0.1) m SE;4—8 cm组, 基茎(5.44±0.4) cm SE, 树高(1.52±0.3) m SE;>8 cm组, 基茎(10.25±0.7) cm SE, 树高(2.1±0.2) m SE。基茎为0—1 cm和1—2 cm为幼龄小梭梭, 基茎2—4 cm为中龄梭梭, 基茎为4—8 cm和>8 cm为成龄大梭梭。
2 研究方法 2.1 土壤含水量测定在6—9月, 每个月中旬, 利用土钻在每株梭梭下方0—300 cm深度内采取土样。在0—20 cm土层每隔10 cm取样, 20—100 cm土层每隔20 cm取样, 100—300 cm的土层每隔40 cm取样, 每个深土层取5个重复。然后, 将装有土样的铝盒(W1)置于烘箱中, 在105℃条件下烘干12 h至恒重, 冷却后称重(W2)。最后每组求其平均值。
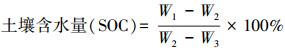
|
式中, W1为湿土加空铝盒质量;W2为干土加铝盒质量;W3是空铝盒质量。
2.2 根系分布调查与根深测算6月初, 采用全根系挖掘法, 对5个基茎组, 共25棵生长健康的梭梭进行根系调查。在每棵选定的梭梭周围挖掘一个环形工作沟, 环形沟内径距梭梭基茎的距离在2—20 m, 内径大小取决于树高和初期调查估算的梭梭个体根区内水平根的拓展范围。根系周围的土壤以0.1 m的间隔被移除以逐步暴露侧根, 收集所有直径大于0.1 mm的根, 并用钢卷尺和游标卡尺测量根的长度和直径。详细的根系挖掘调查方法和根表面积的计算方法参考早期的相关文献[21, 25]。主根深度(即根深)用钢卷尺测量确定。
2.3 同化枝水势的测定与土壤含水量测定同步, 每个月中旬日出前1 h, 测量5个基茎组梭梭黎明前同化枝水势。在每株梭梭的向阳面随机选取3个样枝, 用Model 1000压力室水势仪(PMS Albany USA)测量黎明前同化枝水势(Ψpd)。每株测定5个重复样, 以每次测定水势的平均值作为该组梭梭每月的黎明同化枝水势值。
2.4 压力-容积(P-V)曲线的测定压力-容积曲线采用干燥法测定。8月中旬傍晚, 在梭梭树冠中上层采集生长健康的同化枝, 每株取3枝, 枝条长度5—10 cm。取下后立即放入盛有去离子水的烧杯中, 将其基部浸入水中, 快速带回实验室置于黑暗避光的封闭环境, 使小枝充分饱和吸水12 h。当同化枝达到饱和状态, 在水中切一平口, 然后取出用吸水纸吸干小枝表面多余水分, 迅速用万分之一克天平称其饱和鲜重;随后放入水势仪中缓慢加压测定其水势。在室内自然干燥后测定其质量和水势(初期每隔1—3 min测定一次, 后期每隔10—30 min测定一次), 重复上述过程直至同化枝严重萎蔫。最后将同化枝置于烘干箱, 在105℃条件下烘干至恒重, 称其干重。将一系列同化枝鲜重、干重和水势值输入到压力容积曲线分析程序(PV_Curve_Fitting_5.6 1), 计算出相关水分参数:饱和膨压渗透势(π100)、膨压消失点渗透势(ΨTLP)和细胞壁最大弹性模量(εmax)。
2.5 叶绿素荧光动力学参数和最大气孔导度的测定与同化枝水势测定同步, 每月中旬日出前一小时, 利用植物效率分析仪(Pocket PEA, PE 32 1JL, Hansatech Instruments Ltd., King′s Lynn Norfolk, UK), 测定叶绿素荧光参数。光照强度设定为2500 μmol m-2 s-1, 暗适应20 min后测定初始荧光产量(F0)。随后强闪光(5000 μmol m-2 s-1), 脉冲时间0.7 s, 测定光下荧光产量(Fm)。计算出PSⅡ最大光能转化效率: Fv/Fm=(Fm-F0)/Fm。每月中旬早上10:00—12:00, 利用气孔导度仪(Model SC-1, Decagon)测量5个基茎组梭梭的最大气孔导度(Gs), 每组5个重复。
2.6 数据处理与分析利用SPSS 16.0统计软件进行数据统计分析(SPSS Inc., Chicago, IL, USA), 用单因素方差分析(One-way ANOVA)检验5个基茎组梭梭Ψpd、Fv/Fm、Gs和ΨTLP之间的显著性;利用线性回归拟合分析梭梭生理特性与根深之间的关系, 利用Origin 9.0制图(Origin Lab Corp., Northampton, MA, USA)。
3 结果与分析 3.1 土壤含水量图 1, 6—9月土壤含水量在0.1%—7.9%范围内波动;梭梭土壤含水量随深度的垂直变化规律基本一致, 呈现出先增加后减小的趋势。土壤含水量排序为: 6月>9月>8月>7月。6月, 0—90 cm土层, 土壤含水量随土壤深度逐渐增加, 在90 cm的土层达到峰值(7.9%); 明显是由于融雪和雨水补给的结果。6—7月, 气温逐渐升高, 干旱逐渐加剧, 而梭梭处于旺盛生长期, 短命植物也大量生长发育, 土壤水分蒸散量较高, 从而造成土壤贮水量处于亏损阶段[26]。相较于6月, 7月份土壤含水量已大幅度降低。8月份土壤含水量有所增加, 可能是绿洲用水量的减少和洪水的补给从而引起地下水位的增高造成的[27]。9月份土壤含水量大于7、8月, 则是由于梭梭处于生长末期, 其对水分的生理需要减少, 因此土壤水分状况有所改善[28]。

|
| 图 1 不同深度土壤含水量季节变化 Fig. 1 Seasonal variation of gravimetric soil water content at different depths |
图 1, 土壤深度在0—300 cm范围内, 整体土壤含水量分层变化比较明显。0—90 cm土层, 土壤含水量受融雪、降雨、表面蒸发等因素影响显著, 降水补给能使该层土壤含水量升的很高[29]。90—160 cm土层, 水分在该层下渗过程中, 随着湿润锋下移, 土壤含水量随深度逐渐递减。160—240 cm土层远离地面, 受外界条件影响小, 土壤环境相较稳定, 且不在毛管上升水的影响范围内, 因此水分波动性相对缓和。而240—300 cm土层土壤含水量稍有增加, 则是毛细管水上升的结果。
3.2 根深与根表面积在土壤植物大气连续体中, 根系能够率先感知干旱信号, 协调地上部分的行为, 从而促使植物适应干旱环境[30]。而植根深度有着非常重要的生态学意义, 因为它决定了植被可利用土壤的体积, 植根深度的不同可以导致水分供应以及干旱胁迫的差异[21, 31]。自然生境下, 基茎小的幼年梭梭根系普遍较浅, 根表面积和根冠比较小;而随着梭梭生长发育, 其根系向着深土层发育, 根表面积和根冠比逐渐增加。当成年梭梭基茎为11 cm时, 根深则达到最大值8.4 m(图 2)。

|
| 图 2 不同大小梭梭根系分布与根深 Fig. 2 Root distribution and root depth of H. ammodendron with different basal stem diameter BSD: 基茎basal stem diameter;TA:总根表面积total surface area of the lateral roots;RS:根冠比root-shoot ratio |
π100为饱和含水时的渗透势, 它指示树木在某个生长阶段中可溶物质所能达到的最大浓度[32]。植物π100越低, 说明细胞液浓度就越大, 植物从土壤中吸收水分的潜势越大, 其维持最大膨压的能力就越强[33]。如图 3所示, 基茎为0—1 cm的小梭梭π100与其他基茎组梭梭π100存在显著性差异(P<0.05)。8月份梭梭π100值排序为:0—1 cm < 2—4 cm < 1—2 cm < 4—8 cm<(>8 cm)。这说明同处于干旱胁迫下, 个体小的梭梭同化枝细胞内溶质含量较高, 能保持较低的渗透势, 其保持膨压的能力较强。

|
| 图 3 不同基茎组梭梭饱和膨压、膨压消失点的渗透势和模比系数 Fig. 3 Osmotic potential at full turgor(π100), osmotic potential at turgor lost point (Ψtlp), cell elastic modulus (εmax) of H. ammodendron across different basal stem diameter group(cm) |
初始质壁分离时的渗透势(ΨTLP)亦称为零膨压点时的渗透势, 其大小可反映植物维持最低压力势的极限渗透势[34]。ΨTLP越低, 说明树木维持膨压能力越强。如图 3所示, 梭梭ΨTLP与π100变化趋势基本相同, 且都与基茎的大小呈现出正相关关系。不同基茎组梭梭ΨTLP排序为:>8 cm>4—8 cm>2—4 cm>1—2 cm>0—1 cm。这说明同处于干旱胁迫, 基茎为0—1 cm个体小的梭梭组织细胞能够在较低的渗透势下发生质壁分离, 有利于其叶片在干旱胁迫下保持正常膨压, 推迟萎蔫进程, 避免造成不可逆的伤害。因此从PV水分参数π100、ΨTLP来看, 小梭梭细胞维持膨压能力强于大梭梭。
细胞弹性模量ε是表示细胞壁弹性的参数;随着组织含水量和水势的下降, 弹性较大的组织具有比弹性小组织更大的保持膨压能力[35]。但是ε不是一个常数, 在植物组织失水过程中, 随叶片的相对体积和水势变化而变化, 因此一般采用水分饱和状态下的最大弹性模量εmax表示细胞壁的弹性模量[36]。
在多年生植物生长初期, 其组织幼嫩, 细胞壁刚刚形成, 纤维素含量少, 细胞弹性不高。而随着植物组织成熟, 其细胞壁木质化程度加深, εmax明显升高[27]。当达到一定值后便开始降低[27]。如图 3所示, εmax在基茎为2—4 cm时达到最大值, 之后εmax又开始降低。5个基茎组梭梭的细胞弹性模量εmax排序为: >8 cm < 4—8 cm < 1—2 cm < 0—1 cm < 2—4 cm。这表明在相同生境下, >8 cm和4—8 cm两组基茎较大的梭梭, 其细胞通过改变细胞壁弹性维持膨压的能力较强, 而0—1 cm, 1—2 cm两组基茎较小梭梭的细胞弹性较差, 其细胞通过改变细胞壁弹性保持膨压的能力则相对较弱。
3.4 生理性状叶水势代表植物水分运动的能量水平, 是组织水分状况的直接表现, 反映植物在生长季各种生理活动受环境水分条件的制约程度[37]。黎明前同化枝水势Ψpd则可以很好地反映植物的水分亏损状况;与午后同化枝水势相较, 有受大气状况变化的影响更小、较稳定(低标准差)、直接与土壤水分变化相关的优点[38]。如图 4所示, 6—9月黎明前同化枝水势与土壤水分状况随季节变化趋势具有一致性: 当土壤水分含量高时, 黎明前同化枝水势也相对高。6月, 土壤含水量较高, 各基茎组梭梭黎明前同化枝水势Ψpd均维持较高的水平。6—7月, 随着温度升高, 蒸散作用加剧, 土壤水分降低, 梭梭黎明前同化枝水势呈现显著性降低(P < 0.05);梭梭与土壤之间形成水分梯度, 有利于其从土壤中吸收水分。其中基茎为0—1 cm, 1—2 cm, 2—4 cm组梭梭黎明前同化枝水势降低幅度比较大, 表明基茎小的梭梭受到明显的水分胁迫; 而基茎为4—8 cm和>8 cm组梭梭黎明前同化枝水势降低幅度比较小, 表明基茎较大的梭梭受到干旱胁迫影响程度小。7—8月, 不同基茎组梭梭黎明前同化枝水势基本不变, 都处于相对较低的水平。9月, 随着蒸散作用下降, 土壤水分状况有所改善, 因此不同基茎组梭梭黎明前同化枝水势均有一定幅度上升, 且4—8 cm和>8 cm基茎组梭梭黎明前同化枝水势上升幅度比较小。处于生长季, 5个基茎组平均黎明前同化枝水势值排序依次为:>8 cm>4—8 cm>2—4 cm>0—1 cm>1—2 cm。以上均表明小梭梭夜间水分恢复力不及大梭梭, 所受干旱胁迫强度也大。

|
| 图 4 不同基茎组梭梭黎明前同化枝水势、最大气孔导度和光量子效率季节变化(均值±标准误, n=5) Fig. 4 Seasonal variation of predawn leaf water potential (Ψpd), maximum stomatal conductance (Gs) and maximum photochemical efficiency (Fv/Fm) of H. ammodendron across basal stem diameter class |
气孔导度是植物遭受环境胁迫的敏感指标之一, 可以用于预测植物遭受水、热等因子胁迫的程度[39]。如图 4所示, 在每个月的同一测量日, 各基茎组梭梭的最大气孔导度(Gs)无显著性差异(P>0.05)。6—7月, 基茎为0—1 cm和4—8 cm组梭梭的最大气孔导度(Gs)都呈现上升趋势, 7月0—1 cm基茎组梭梭的最大气孔导度(Gs)达到142.28 mmol m-2 s-1, 4—8 cm基茎组梭梭的最大气孔导度(Gs)达到167.52 mmol m-2 s-1。7—8月, 除了2—4 cm和>8 cm基茎组梭梭的气孔导度(Gs)呈现增加的趋势外, 其他基茎组梭梭的最大气孔导度(Gs)均降低;8—9月份只有基茎>8 cm组梭梭的最大气孔导度(Gs)显著降低(P<0.05)。6—9月, 5个基茎组梭梭的平均最大气孔导度排序为:4—8 cm(115.445 mmol m-2 s-1) >>8 cm(93.575 mmol m-2 s-1)>0—1 cm(91.145 mmol m-2 s-1)>2—4 cm(90.885 mmol m-2 s-1) >1—2 cm(83.225 mmol m-2 s-1)。4—8 cm基茎组梭梭有较高的平均最大气孔导度, 可能是其以失水为代价维持较高的气孔导度使之能在干旱胁迫下进行最大限度的碳积累。7月, 0—1 cm基茎组梭梭的最大气孔导度(Gs)也较高, 可能是其通过提高气孔导度加快蒸腾, 以减少面临干旱和高温的双重胁迫[40]。
Fv/Fm——是PSⅡ最大光化学量子产量, 反映PSⅡ反应中心内禀光能转化效率或最大PSⅡ的光能转化效率[41]。如图 4所示, 6—9月, 梭梭的Fv/Fm值范围在0.71—0.79之内, 且梭梭的Fv/Fm值整体呈现下降趋势。6—7月, 胁迫初期的梭梭Fv/Fm值无显著差异(P>0.05), 均呈现极小幅度降低的趋势。7—8月, 基茎为0—1 cm, 1—2 cm, 2—4 cm和>8 cm梭梭Fv/Fm值均呈现显著性下降(P<0.05), 且基茎为0—1 cm、1—2 cm、2—4 cm梭梭Fv/Fm值下降幅度大于基茎为4—8 cm, >8 cm梭梭的降幅。8—9月, 随着水分胁迫缓和, 梭梭Fv/Fm都有小幅度回升。
3.5 植物性状与根深关系如图 5和图 6, 对不同大小梭梭根深与生理参数之间进行相关性分析。8—9月, 五个基茎组梭梭的Ψpd、Fv/Fm、PV水分参数π100、ΨTLP都与根深呈现显著的正相关, 即: 拥有深根的大梭梭往往有较高Ψpd、Fv/Fm和PV水分参数π100、ΨTLP。Fv/Fm、PV水分参数π100、ΨTLP与Ψpd也呈现较高的的正相关, 即: 拥有较高Ψpd的大梭梭往往有较高的Fv/Fm和PV水分参数π100、ΨTLP。

|
| 图 5 根深与植物性状关系 Fig. 5 Relationship between root depth and plant traits |

|
| 图 6 黎明前同化枝水势与植物性状的关系 Fig. 6 Relationship between predawn leaf water potential and plant traits |
植物抗旱机制是复杂多样的, 同一种植物可能有几种抗旱方式, 随着水分、物候、年龄等条件的变化, 其抗旱机制也是变化的[42]。水分逆境下, 植物通过“渗透调节”和“弹性调节”共同维持膨压[43-44]。而渗透势在|π100-ΨTLP |变化范围内, 其差值则可以直接反映植物组织的渗透调节能力;差值越大, 则表明渗透调节的能力越强[32, 42]。在本实验中, |π100-ΨTLP|的差值排序依次为: 0—1 cm(2.15 MPa)>1—2 cm(1.97 MPa)>4—8 cm(1.7 MPa)>2—4 cm(1.53 MPa)>>8 cm(1.45 MPa)。小梭梭拥有较高的|π100-ΨTLP|差值, 表明小梭梭通过积累渗透物质以降低细胞渗透势, 从而用来增加自身吸水的能力较强。而基茎大的梭梭π100、ΨTLP值较高, εmax值最小, 表明大梭梭在较高的渗透势下发生质壁分离, 其细胞壁柔软且有较大的弹性, 能在环境胁迫下能较快的收缩[35], 从而有利于其在低水势下仍能保持高膨压并防止细胞破裂。因此, 在梭梭生长发育不同阶段, 对于通过调节膨压以适应水分胁迫可能有着不同策略, 即: 生长发育初期小梭梭细胞主要以渗透调节维持膨压, 而随着生长发育, 大梭梭主要以弹性调节来维持膨压。
有关研究同样表明一些树种的水分生理参数可能随植株大小和不同发育阶段而改变[45-46]。树种的气孔导度与净光合速率一般随大小或树龄的增加而降低被广泛报道[46-47]。然而也有一些研究与此不同。如,在Nabeshima等研究中,三种槭属树木的气孔导度和黎明前叶水势随其个体大小增加而下降,净光合速率则基本保持不变[48]。在Hazandy等研究中,欧洲白蜡树(Fraxinus excelsior)的净光合速率随其大小增加而降低,气孔导度则是先升高后降低[49]。但是在本研究中, 小梭梭生长季的黎明前同化枝水势与平均最大气孔导度均低于大梭梭,这与Bao等对柠条(Caragana korshinskii)的研究结果相反[50]。
植物在未遭受环境胁迫, 并经过充分暗适应的植物叶片PSⅡ最大的潜在量子效率是比较恒定的, 一般为0.80—0.85[51]。而处于胁迫条件下, 其叶片吸收过剩的光能就会引起Fv/Fm的降低而发生光抑制[52], 因此Fv/Fm常被用来衡量植物PSⅡ是否因发生光抑制而造成不可逆损伤。8—9月, 干旱胁迫下梭梭Fv/Fm与根深呈正相关(图 5)。以上表明浅根系小梭梭光合机能受到干旱损伤引起光抑制的程度高, 生理活动受水分条件的制约程度也较大。
4.2 根深在不同大小梭梭应对夏季干旱胁迫中的重要作用对于荒漠植物, 水分是干旱区生态系统中最主要的限制性因子[53]。在强烈或者长期干旱条件下, 荒漠植物生物量的分配模式使其所受水分资源限制最小化, 而水分资源获取和净初级生产力(NPP)最大化[54]。根系成为荒漠植物生物量配置的主要部位, 植物优先向根系分配新的生物量用于构建根系, 以增加对土壤水分和养分摄取[55]。荒漠灌木根系分布、同化器官生长与环境水分有效性之间的关系决定着植物的存活[56]。因此在水分匮乏的生境中, 根深对于不同植株幼苗的建成、水分养分的吸收、水力结构和植物的生存是至关重要的[21, 57]。在本研究中, 梭梭根深和生理性状之间显著相关关系(图 5)表明:根深对于不同大小梭梭安全渡过干旱胁迫中起到重要作用。
成年梭梭根系发达, 植根较深, 且根系具有“二态性”。由于冬、春季降水的补给, 当浅层土壤含水量高时, 分布在表层土壤中的梭梭水平侧根可以较好的利用浅层土壤水, 利用比例达62%—95%。随着干旱胁迫的加剧, 强烈的蒸散发作用使得浅层土壤水分亏缺得不到及时补充(图 1);成年梭梭则减少对浅层土壤水分利用, 其分布在深层土壤中的根系增加对深层土壤水及地下水的利用(图 2), 且对地下水的利用比例高达68%—100%[58-60]。对于成年梭梭而言, 深根使之能在干旱胁迫下保持更加有利水分条件(其黎明前同化枝水势:(-3.80±0.18) MPa高于浅根小梭梭黎明前同化枝水势:(-6.45±0.44) MPa), 从而更好的适应干旱胁迫。而幼龄梭梭向根系分配生物量相对少, 根深较浅且侧根主要分布在浅土层(图 2), 其水分状态受到浅层土壤水分影响显著(图 1);旱季主要利用小概率事件的降水, 易受到干旱胁迫影响。因此处于生长期的小梭梭对干旱胁迫敏感, 以较高死亡率为代价抢占更多资源, 通过逐渐构建根系来提高捕捉深层土壤水或者地下水的能力[61]。此外, 小梭梭有着相对较低的平均最大气孔导度, 其根深与π100、ΨTLP也有着显著正相关关系(图 5), 这表明小梭梭采取了更为积极的渗透调节与气孔控制抵御干旱。但是8、9月, 小梭梭Ψpd较低(图 4), 且生理性状与Ψpd之间的显著正相关关系(图 6)仍表明小梭梭受到更为严重的水分胁迫。基于上述分析, 不同大小梭梭水分状况的差异是导致其夏季干旱期间生理性状不同的原因, 梭梭植根深度决定了其水分状况, 进而决定了梭梭的生理性状表现。
由于测定条件、测量方法和树木种类的差异等因素, 本发现可能并不适用于其他生境下的树种。然而该研究工作却促进对处于不同发育阶段或不同大小的荒漠植物水分动态变化响应机制的认识, 有利于在气候变化背景下预测梭梭种群动态变化。鉴于当前全球气温升高伴随着降水格局及氮沉降量的改变, 同时CO2浓度升高、温度升高和干旱对植物还有着交互或协同作用[62-63], 因此建议在以后的研究中要综合考虑多种因素, 从个体发育阶段乃至种群空间分布格局上对梭梭进行长期动态观测和统计学检验, 对本结果进行更加系统深入的验证。
5 结论在越夏生长期间, 不同发育阶段或不同个体大小的梭梭在生理性状表现出明显的差异;尽管梭梭幼苗可以通过渗透调节和气孔控制策应夏季干旱胁迫, 然而并不足以使幼苗保持好的水分状况, 因此梭梭幼苗受到了更为严酷的干旱胁迫。研究结果也表明:根系深度决定了不同个体大小梭梭的水分状况, 进而主导其越夏生长期间的生理性状响应的差异。对于幼龄梭梭而言, 抗旱策略(根系)发育不完全, 当发生干旱时, 易受水分胁迫。而对成年梭梭而言, 持久的优先地下生物量分配使其根系发达, 能通过吸收深层土壤水和地下水来维持较好的自身水分状况从而规避夏季干旱的影响。
| [1] |
李世兰. 降水变化对长白山红松阔叶林土壤微生物特征的影响[D]. 哈尔滨: 东北林业大学, 2014: 1-1.
|
| [2] |
莫兴国, 胡实, 卢洪健, 林忠辉, 刘苏峡. GCM预测情景下中国21世纪干旱演变趋势分析. 自然资源学, 2018, 33(7): 1244-1256. |
| [3] |
张婷, 曹扬, 陈云明, 刘国彬. 生长季末期干旱胁迫对刺槐幼苗非结构性碳水化合物的影响. 水土保持学报, 2016, 30(5): 297-304. |
| [4] |
周倩, 曹家畅, 崔明昆, 龚明. 非结构性糖在植物对干旱胁迫响应与适应中的作用. 安徽农业科学, 2018, 46(30): 24-28. DOI:10.3969/j.issn.0517-6611.2018.30.008 |
| [5] |
段洪浪, 吴建平, 刘文飞, 廖迎春, 张海娜, 樊后保. 干旱胁迫下树木的碳水过程以及干旱死亡机理. 林业科学, 2015, 51(11): 113-120. |
| [6] |
Breshears D D, López-Hoffman L, Graumlich L J. When ecosystem services crash: preparing for big, fast, patchy climate change. Ambio, 2011, 40(3): 256-263. DOI:10.1007/s13280-010-0106-4 |
| [7] |
He J S, Zhang Q B, Bazzaz F A. Differential drought responses between saplings and adult trees in four co-occurring species of New England. Trees, 2005, 19(4): 442-450. DOI:10.1007/s00468-004-0403-2 |
| [8] |
Rohner B, Bigler C, Wunder J, Brang P, Bugmann H. Fifty years of natural succession in Swiss forest reserves: changes in stand structure and mortality rates of oak and beech. Journal of Vegetation Science, 2012, 23(5): 892-905. DOI:10.1111/j.1654-1103.2012.01408.x |
| [9] |
Zang C, Pretzsch H, Rothe A. Size-dependent responses to summer drought in Scots pine, Norway spruce and common oak. Trees, 2012, 26(2): 557-569. DOI:10.1007/s00468-011-0617-z |
| [10] |
Reynolds J H, Ford E D. Improving competition representation in theoretical models of self-thinning: a critical review. Journal of Ecology, 2005, 93(2): 362-372. DOI:10.1111/j.1365-2745.2005.00976.x |
| [11] |
平晓燕, 周广胜, 孙敬松. 植物光合产物分配及其影响因子研究进展. 植物生态学报, 2010, 34(1): 100-111. DOI:10.3773/j.issn.1005-264x.2010.01.013 |
| [12] |
陈仁飞, 姬明飞, 关佳威, 邓建明. 植物对称性竞争与非对称性竞争研究进展及展望. 植物生态学报, 2015, 39(5): 530-540. |
| [13] |
Xie J B, Xu G Q, Jenerette G D, Bai Y F, Wang Z Y, Li Y. Apparent plasticity in functional traits determining competitive ability and spatial distribution: a case from desert. Scientific Reports, 2015, 5(1): 12174. DOI:10.1038/srep12174 |
| [14] |
俞阗. 荒漠植物梭梭对地表高温胁迫的响应及"花后生殖休眠"现象的研究[D]. 南京: 南京农业大学, 2012: 4-8.
|
| [15] |
Song J, Feng G, Tian C Y, Zhang F S. Osmotic adjustment traits of Suaeda physophora, Haloxylon ammodendron and Haloxylon persicum in field or controlled conditions. Plant Science, 2006, 170(1): 113-119. DOI:10.1016/j.plantsci.2005.08.004 |
| [16] |
Xu H, Li Y, Xu G Q, Zou T. Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation. Plant, Cell & Environment, 2007, 30(4): 399-409. |
| [17] |
秦海波, 郑新军, 李彦, 马杰, 李嵩, 唐立松. 荒漠灌木梭梭同化枝相对生长速率对温度升高响应的不确定性. 中国沙漠, 2012, 32(5): 1335-1341. |
| [18] |
李彦, 许皓. 梭梭对降水的响应与适应机制——生理、个体与群落水平碳水平衡的整合研究. 干旱区地理, 2008, 31(3): 313-323. |
| [19] |
张立运, 陈昌笃. 论古尔班通古特沙漠植物多样性的一般特点. 生态学报, 2002, 22(11): 1923-1932. DOI:10.3321/j.issn:1000-0933.2002.11.018 |
| [20] |
黄培祐, 李启剑, 袁勤芬. 准噶尔盆地南缘梭梭群落对气候变化的响应. 生态学报, 2008, 28(12): 6051-6059. DOI:10.3321/j.issn:1000-0933.2008.12.033 |
| [21] |
Xu G Q, McDowell N G, Li Y. A possible link between life and death of a xeric tree in desert. Journal of Plant Physiology, 2016, 194: 35-44. DOI:10.1016/j.jplph.2016.02.014 |
| [22] |
谢江波, 彭历芝, 李艳明, 李彦. 两种梭梭属幼苗在土壤N和P梯度下的竞争能力研究. 干旱区地理, 2018, 41(1): 83-91. |
| [23] |
周宏飞, 李彦, 汤英, 周宝佳, 徐宏伟. 古尔班通古特沙漠的积雪及雪融水储存特征. 干旱区研究, 2009, 26(3): 312-317. |
| [24] |
宋于洋, 楚光明, 胡晓静. 古尔班通古特沙漠梭梭种群径级与龄级关系的研究. 西北植物学报, 2011, 31(4): 808-814. |
| [25] |
Xu H, Li Y. Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events. Plant and Soil, 2006, 285(1/2): 5-17. |
| [26] |
王泽锋, 胡顺军, 李浩. 古尔班通古特沙漠南缘丘间地梭梭群落蒸散特征. 干旱区地理, 2018, 41(6): 1303-1309. |
| [27] |
曾凡江, 李向义, 张希明, Foetzki A, Arndt S K, Runge M. 极端干旱条件下多年生植物水分关系参数变化特性. 生态学杂志, 2010, 29(2): 207-214. |
| [28] |
解婷婷, 张希明, 梁少民, 单立山, 杨小林, 花永辉. 不同灌溉量对塔克拉玛干沙漠腹地梭梭水分生理特性的影响. 应用生态学报, 2008, 19(4): 711-716. |
| [29] |
朱海, 胡顺军, 刘翔, 李浩, 李宜科. 不同龄阶梭梭根区土壤水分时空变化特征. 生态学报, 2017, 37(3): 860-867. |
| [30] |
张林森, 张海亭, 胡景江, 权静, 胥生荣, 韩明玉, 马锋旺. 两种苹果砧木根系水力结构及其PV曲线水分参数对干旱胁迫的响应. 生态学报, 2013, 33(11): 3324-3331. |
| [31] |
Chapin Ⅲ F S, Matson P A, Mooney H A. 陆地生态系统生态学原理. 李博, 赵斌, 彭容豪, 译. 北京: 高等教育出版社, 2005: 71-71.
|
| [32] |
韩刚, 李彦瑾, 孙德祥, 赵忠. 4种沙生灌木幼苗PV曲线水分参数对干旱胁迫的响应. 西北植物学报, 2008, 28(7): 1422-1428. DOI:10.3321/j.issn:1000-4025.2008.07.023 |
| [33] |
韩若霜, 王林和, 张国盛, 高峻, 刘雁南. 13种园林树木PV曲线水分参数的比较. 干旱区资源与环境, 2015, 29(7): 134-139. |
| [34] |
冯玉龙, 巨关升, 朱春全. 杨树无性系幼苗光合作用和PV水分参数对水分胁迫的响应. 林业科学, 2003, 39(3): 30-36. DOI:10.3321/j.issn:1001-7488.2003.03.005 |
| [35] |
谭永芹, 柏新富, 朱建军, 王仲礼, 刘林德. 干旱区五种木本植物枝叶水分状况与其抗旱性能. 生态学报, 2011, 31(22): 6815-6823. |
| [36] |
石松利, 王迎春, 周红兵, 周健华. 濒危种四合木与其近缘种霸王水分关系参数和光合特性的比较. 生态学报, 2012, 32(4): 1163-1173. |
| [37] |
付爱红, 陈亚宁, 李卫红, 张宏锋. 干旱、盐胁迫下的植物水势研究与进展. 中国沙漠, 2005, 25(5): 744-749. DOI:10.3321/j.issn:1000-694X.2005.05.021 |
| [38] |
曹永慧, 周本智, 倪霞, 葛晓改, 王小明. 模拟干旱下毛竹叶片水势的动态变化. 林业科学研究, 2018, 31(4): 183-191. |
| [39] |
喻方圆, 徐锡增. 植物逆境生理研究进展. 世界林业研究, 2003, 16(5): 6-11. DOI:10.3969/j.issn.1001-4241.2003.05.002 |
| [40] |
周洪华, 李卫红, 木巴热克·阿尤普, 徐茜. 荒漠河岸林植物木质部导水与栓塞特征及其对干旱胁迫的响应. 植物生态学报, 2012, 36(1): 19-29. |
| [41] |
张守仁. 叶绿素荧光动力学参数的意义及讨论. 植物学通报, 1999, 16(4): 444-448. DOI:10.3969/j.issn.1674-3466.1999.04.021 |
| [42] |
狄晓艳, 王孟本, 陈建文, 张伟峰. 杨树无性系PV曲线水分参数的研究. 西北植物学报, 2007, 27(1): 98-103. DOI:10.3321/j.issn:1000-4025.2007.01.017 |
| [43] |
闫子娟, 赵鑫, 郝蕾, 王颖, 白玉荣, 张国盛, 李小龙. 利用PV曲线水分参数评价16种植物抗旱性. 西部林业科学, 2017, 46(1): 112-116, 133-133. |
| [44] |
马瑞昆, 蹇家利, 刘淑贞, 李香菊. 改进的作物水分关系PV曲线的压力室分析技术. 河北农业大学学报, 1989, 12(3): 54-59. |
| [45] |
Colangelo M, Camarero J J, Borghetti M, Gazol A, Gentilesca T, Ripullone F. Size matters a lot: drought-affected Italian oaks are smaller and show lower growth prior to tree death. Frontiers in Plant Science, 2017, 8: 135. |
| [46] |
Steppe K, Niinemets U, Teskey R. Tree size-and age-related changes in leaf physiology and their influence on carbon gain. Meinzer F, Lachenbruch B, Dawson T eds. Size- and Age-Related Changes in Tree Structure and Function. Dordrecht: Springer SBM, 2011: 235-253.
|
| [47] |
Bond B J. Age-related changes in photosynthesis of woody plants. 2000, 5(8): 349-353.
|
| [48] |
Nabeshima E, Hiura T. Size-dependency in hydraulic and photosynthetic properties of three Acer species having different maximum sizes. Ecological Research, 2008, 23(2): 281-288. DOI:10.1007/s11284-007-0374-z |
| [49] |
Hazandy A H, Maurizio M. Age- and size-related changes in physiological characteristics and chemical composition of Acer pseudoplatanus and Fraxinus excelsior trees. Tree Physiology, 2009, 29(1): 27-38. |
| [50] |
Bao J T, Wang J, Li X R, Zhang Z S, Su J Q. Age-related changes in photosynthesis and water relations of revegetated Caragana korshinskii in the Tengger desert, Northern China. Trees, 2015, 29(6): 1749-1760. DOI:10.1007/s00468-015-1255-7 |
| [51] |
许大全. 光合作用学. 北京: 科学出版社, 2013: 94-94.
|
| [52] |
沈亮, 陈君, 刘赛, 徐荣, 徐常青, 刘同宁. 脱水胁迫和光合日变化对梭梭和白梭梭叶绿素荧光参数的影响. 应用生态学报, 2015, 26(8): 2321-2328. |
| [53] |
李志军, 罗青红, 伍维模, 韩路. 干旱胁迫对胡杨和灰叶胡杨光合作用及叶绿素荧光特性的影响. 干旱区研究, 2009, 26(1): 45-52. |
| [54] |
Chapin Ⅲ F S, Matson P A, Mooney H A. 陆地生态系统生态学原理. 李博, 赵斌, 彭容豪, 译. 北京: 高等教育出版社, 2005: 113-114.
|
| [55] |
周朝彬, 王梦瑶, 龚伟. 梭梭射线组织特征与非结构性碳水化合物含量的关系. 干旱区研究, 2018, 35(5): 1105-1110. |
| [56] |
许皓, 李彦, 邹婷, 谢静霞, 蒋礼学. 梭梭(Haloxylon ammodendron)生理与个体用水策略对降水改变的响应. 生态学报, 2007, 27(12): 5019-5028. DOI:10.3321/j.issn:1000-0933.2007.12.010 |
| [57] |
Alvarez-Flores R, Winkel T, Nguyen-Thi-Truc A, Joffre R. Root foraging capacity depends on root system architecture and ontogeny in seedlings of three Andean Chenopodium species. Plant and Soil, 2014, 380(1/2): 415-428. DOI:10.1007/s11104-014-2105-x |
| [58] |
傅思华, 胡顺军, 李浩, 王泽锋. 古尔班通古特沙漠南缘梭梭(Haloxylon ammodendron)群落优势植物水分来源. 中国沙漠, 2018, 38(5): 1024-1032. |
| [59] |
戴岳, 郑新军, 唐立松, 李彦. 古尔班通古特沙漠南缘梭梭水分利用动态. 植物生态学报, 2014, 38(11): 1214-1225. |
| [60] |
彭丽萍, 戴岳, 师庆东. 新疆准东荒漠区5种典型植物水分来源. 干旱区研究, 2018, 35(5): 1146-1152. |
| [61] |
田媛, 唐立松, 乔瑞平. 梭梭幼苗个体生长规律与死亡率关系研究. 植物研究, 2016, 36(1): 84-89. |
| [62] |
张维, 赵文勤, 谢双全, 张莉, 吕新华, 贺亚玲. 模拟降水和氮沉降对准噶尔盆地南缘梭梭光合生理的影响. 中国生态农业学报, 2018, 26(1): 106-115. |
| [63] |
李树鑫, 卢元兵, 段宝利, 张远彬. 杨树(Populus)生理生态特性对增温、大气CO2浓度升高和干旱响应的Meta分析. 山地学报, 2017, 35(5): 636-644. |