 2021, Vol. 41
2021, Vol. 41文章信息
- 韩涛涛, 唐玄, 任海, 王俊, 刘楠, 郭勤峰
- HAN Taotao, TANG Xuan, REN Hai, WANG Jun, LIU Nan, GUO Qinfeng
- 群落/生态系统功能多样性研究方法及展望
- Community/Ecosystem Functional Diversity: measurements and development
- 生态学报. 2021, 41(8): 3286-3295
- Acta Ecologica Sinica. 2021, 41(8): 3286-3295
- http://dx.doi.org/10.5846/stxb201903080442
-
文章历史
- 收稿日期: 2019-03-08
- 网络出版日期: 2021-02-24

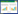




2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;
3. 中国科学院大学, 北京 100049;
4. 美国农业部东部森林威胁评估中心, RTP, NC 27709
2. State Key Laboratory of Urban and Regional Ecology, Chinese Academy of Sciences, Beijing 100085, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Eastern Forest Environmental Threat Assessment Center, USDA Forest Service, RTP, NC 27709, USA
不同物种为什么可以共存?物种多样性如何影响生态系统功能?这些问题是生态学研究中的经典问题, 也是当前急需解决的科学难题[1]。要解决这些问题首先需要对物种多样性进行量化[2]。在以往的研究中, 生态学家主要基于Shannon-Weiner指数, Simpson指数等以物种为基本单元的多样性指数来探讨群落物种多样性。然而, 物种间的差异往往体现在物种对于不同环境的适应, 即植物功能性状(Plant functional traits)。以物种为基本单元的研究很难体现不同群落/生态系统间功能特征的差异[3-4]。而基于植物功能性状、功能多样性的研究可以从植物的生长、定居、生殖、防御、持续性等功能策略方面解释不同群落/生态系统间的物种组成、功能特征, 从而有助于进一步理解不同群落/生态系统的生态过程和功能[2, 5-7]。
近30年来, 随着群落生态学及生态系统生态学的发展, 植物功能性状越来越多的被应用到生态学研究中[8-9]。基于植物功能性状的研究方法也已应用到从植物个体水平到群落水平乃至到生态系统水平等各个尺度的研究中。依据不同尺度规则, 可选取不同类型的性状指标来评估特定尺度下生态系统的内在组分(图 1)[6, 8, 10-11]。功能多样性是指可以影响生态系统功能的物种性状值和范围[6], 是解释和预测群落/生态系统过程和功能的重要指标, 对于深入理解群落构建机制、物种多样性维持机理、生态系统服务功能等具有重要意义。当前功能多样性的相关研究依旧处于理论探讨方面, 野外实证研究相对缺乏[12-16]。对于不同功能多样性概念及研究方法的进一步梳理、比较、探讨, 有助于解决上述问题。

|
| 图 1 利用植物功能性状进行不同尺度的生态学研究框架 Fig. 1 Plant traits vs. functional traits and their general roles in community/ecosystems studies at different levels and scales |
功能多样性是生物集群的重要特征之一, 是解释和预测生态系统过程、功能的有力工具[12-13]。目前大多关于功能多样性的定义并没有有效的实践价值, 对于功能多样性的不同理解也催生了多种功能多样性的定量化研究方法[14-17]。在研究中, 这些方法对于植物功能性状选取、功能多样性计算、生态系统功能评估等差异较大。因此有必要对功能多样性概念进行统一。Diaz和Cabido[5]认为, 功能多样性是指生态系统中有机体所表现出来的功能性状数目、类型和分布;Poos等[18]认为功能多样性是指群落/生态系统中物种间功能性状的差异程度;Tilman[6]认为功能多样性是可以影响生态系统功能的物种或有机体的性状值和范围。由于Tilman的功能多样性概念具有较强的概括性和可操作性, 被普遍接受并被广泛用于计算/评估群落/生态系统结构与功能的各个方面[12-15]。
当前关于群落/生态系统功能多样性的研究方法大致有10种[7, 14, 16-17], 它们不仅可以对群落/生态系统功能进行定性描述, 也可以进行定量分析。这对进一步深化理解群落/生态系统结构与功能具有重要意义。然而, 不同研究方法得出的结果差异较大, 且难以进行多研究间比较研究[16-17]。比较探讨各功能多样性研究方法的优缺点有利于拓展功能多样性内涵, 也有助于功能多样性研究方法的应用及发展。本文旨在探讨如何利用功能性状量化群落/生态系统的功能多样性, 着重分析目前关于功能多样性的主要研究方法。包括植物功能性状选取、性状权重、功能多样性指数计算及应用等问题, 以期为进一步丰富群落/生态系统功能多样性研究提供理论参考。
1 植物功能性状预处理 1.1 植物功能性状类型及选取为便于植物功能性状研究, 有学者对植物性状进行了大致分类, 比如, 形态性状和生理性状;营养性状、防御性状及繁殖性状;地上性状和地下性状;以及“软性状”和“硬性状”等[9, 18-19]。根据全球植物性状数据库(Plant Trait Database), 目前至少有2091种植物功能性状被使用[20]。而现阶段在植物功能性状的选取方面仍缺乏标准方法, 因为这强烈依赖于所要研究的科学问题、生态特性及研究尺度等[21]。例如, 与传粉者、捕食者有关的科学问题, 植物对水环境、营养条件响应相关的科学问题等, 均对性状的选取具有显著依赖性[22-23]。又如, 在研究植物丰度动态与气候变化关系时, 有学者采用比叶面积、叶片质量、根氮浓度、比根长等代表植物资源捕获效率, 叶厚度、叶干物质质量、叶根碳浓度、叶碳氮比等代表植物资源保持策略, 以及种子大小等代表植物生殖策略[24]。在研究植物功能性状与环境因子间相关性随演替变化时, 采用最大光合速率、比叶面积、叶片干物质质量、木材密度、潜在最大高度、叶片氮、磷及总有机碳含量等代表植物竞争力、养分循环策略[25]。
盲目选择及追求过于简单、易测量性状难以对所要研究的科学问题起到较好帮助。有研究表明, 基于生活史的功能型组和基于简易的多样性指数并不能很好的预测地上生物量[13]。又如人为干扰后, 土壤种子库类型比与群落恢复无关的功能性状对于群落恢复更有意义。通常在决定使用多种功能性状进行研究时, 需考虑如下问题:(1)选取何种功能性状, 它们是否对所研究科学问题具有相同权重;(2)若所选性状存在尺度差异, 如何进行性状联结;(3)计算群落/生态系统功能多样性时, 是否对性状进行权重、如何权重[2]。
1.2 功能性状间相关性处理在植物功能性状研究中, 很多功能性状存在一定的相关性[26-27]。即在描述功能多样性时, 所选功能性状可能存在冗余现象。例如, 种子质量与种子大小在自然条件下显著相关, 如果在研究中同时选取两者, 种子性状就会在功能多样性指数中具有双倍权重。然而, 当前关于性状相关性问题的研究仍处于探索阶段[28]。当选取功能性状存在显著相关时, 或所选取性状对于所要研究生态系统的功能影响较小时, 需要对这些性状进行降权处理。
一般可使用基于代数等数学方法对相关性状进行降权, 比如Mason等[14]基于方差-协方差的广义马氏距离处理方法[29]。然而在实际研究中, 一些相关性状其生态学意义可能不尽相同, 比如种子大小与种子数量。另外, 许多不相关性状在一些特殊数据集中也可能相关。Leps等[2]指出使用主成分分析方法可以实现对相关性状的降权处理, 同时也能够解决相关性状在多样性指数中权重过高的问题。但如果相关性状能够反映不同的生态系统功能信息, 在功能多样性计算时, 不应对其进行降权。因此, 在选取性状的过程中, 应选择与所要研究生态学问题相关但彼此间不相关的性状。值得借鉴的方法有Westoby[30]的叶-高度-种子策略(LHS策略)、Wright等[31]的叶经济谱策略(LES策略)、Gillison[11, 32]的叶-生活型-根策略(LLR策略)。这些研究方法所选取的植物功能性状在很大程度上可以反应植物的生存、生活、定植、持续性等策略, 且彼此相关程度不高。
1.3 功能性状数据预处理当前很多关于功能多样性的研究均会使用多重功能性状[28, 33-34]。然而, 由于功能性状间的差异性(如存在不同量纲等现象), 在进行功能多样性计算之前, 有必要对性状数据进行转化或标准化处理。常见转化方式有对数转化、Z转化、利用性状极差转化等[2]。
1.4 功能性状权重方法质量比率假说认为, 生态系统功能在很大程度上由生物量占优势的物种决定[11, 34]。因此在计算群落/生态系统功能多样性时, 需对物种功能性状进行权重调节。目前有3种权重方法:(1)基于物种丰富度(物种数量);(2)基于物种在样方中的出现频率;(3)基于物种生物量(为便于野外工作, 很多学者采用基于植物茎占面积替代)[35-36]。其中, 物种丰富度与种群动态过程相关, 如出生率, 扩散率等;物种出现频率与物种竞争力相关;生物量与生态过程如生态系统生产力、分解力、资源利用率相关。
Garnier等[37]认为仅考虑优势物种功能性状不会影响与生物多样性有关的群落/生态系统整体功能特征, 即仅使用群落中优势物种的功能性状可以反映生态系统的主要特征。因此在进行群落/生态系统功能多样性计算时, 选取群落内优势物种的研究可以在一定程度上弱化因不同权重方法所导致的研究结果差异。此外, 采取何种权重方法也依赖于所要研究的科学问题, 比如, 在研究群落/生态系统生产力时, 使用基于生物量(茎占面积)的权重方法可以更好描述群落/生态系统生产力的真实水平。
1.5 种内功能性状差异当前大多功能多样性研究方法均忽略了种内性状差异[7, 12, 14], 而其对于群落/生态系统的整体功能亦具有重要影响[38-39]。比如在森林中, 成年植株个体与幼年个体在光合作用、水分、营养元素利用方面具有较大差异;又如, 在同一群落中, 由于微环境的差异, 不同植株个体的生态功能也不尽相同。Plourde等[40]在研究热带森林植物木质部密度随森林演替变化时认为, 物种种内差异、功能群差异等均对木质部密度随演替的变异产生重要影响。Kichenin等[41]在研究新西兰高山亚高山植物群落功能特征随环境梯度变化时发现, 物种种内差异与种间差异对于环境变化具有不同的响应。Bolnick等[38]综合前人的研究, 提出6种植物功能性状种内变异对于群落/生态系统结构和功能的影响机制, 认为在经典生态学模型的研究中, 应重新审视种内性状变异对于所要研究问题的重要性。总之, 在预测群落和生态系统过程与功能时, 同时考虑种内种间性状差异有助于更深入认识所要研究的科学问题。同时, 也将拓宽群落生态学的研究范畴, 即从以物种研究为基础向以植物个体研究为基础转变, 从而有助于更具预测性生态学理论的提出[42]。
2 功能多样性研究方法物种的大量功能性状指标, 生境中的大量共存物种, 以及当前对于植物功能性状如何影响生态系统功能认识的不充分性均导致了难以以一种简便方法去衡量群落/生态系统功能多样性。在以往生态学研究中, 生态学家常常采用一些简单易测量的指标替代功能多样性, 比如, (1)物种丰富度或生境中物种数量;(2)功能群组数量;(3)同一功能组内物种多样性[6]。近年来, 随着计算机等技术的快速发展, 越来越多的复杂数学方法被引入到功能生态学研究中, 产生了一些定量化的功能多样性指标[7, 14, 18, 33-34, 43]。这些指标不仅可以研究定量的功能性状数据、也可以研究定性、或定性定量的混合数据;同时也逐渐对数据格式不再有太多严格要求(如允许缺失值存在)。这些研究方法为野外研究提供了极大的实践价值。表 1简述了当前关于功能多样性研究的部分定量化研究方法。
| 指标 Index |
简要描述 Summary |
性状数 Trait number |
考虑种内差异 Intraspecific difference |
考虑物种丰度 Species abundance |
考虑物种库 Species pool |
参考来源 Reference |
| 功能属性多样性指数 Functional attribute diversity (FAD) |
配对物种间的距离总和 | 单个性状 | 否 | 否 | 否 | [12] |
| 功能多样性指数 Functional diversity (FD) |
功能树图总枝长度 | 多个性状 | 否 | 否 | 是 | [7] |
| 功能性状变异范围 Variance of functional diversity (FDvar) |
功能性状的变异程度总和 | 单个性状 | 否 | 是 | 否 | [15] |
| Rao二次熵系数 Rao′s quadratic entropy (FDQ) |
由相对丰度权重的物种对间的距离之和 | 多个性状 | 是 | 是 | 否 | [16] |
| 总功能规则性指数 Overall functional regularity (OFRO) |
所有物种在功能空间分布的均匀性总和 | 多个性状 | 否 | 是 | 否 | [4] |
| 功能丰富度 Functional richness (FRic) |
包含所有物种的最小功能性状空间凸壳体积 | 多个性状 | 否 | 是 | 否 | [33] |
| 功能均匀性 Functional evenness (FEve) |
相对丰度权重的功能树图枝长总和 | 多个性状 | 否 | 是 | 否 | [33] |
| 功能分歧度 Functional divergence (FDiv) |
物种偏离由相对丰度权重的中心的平均距离 | 多个性状 | 否 | 是 | 否 | [33] |
| 改良的功能属性多样性 Modified functional attribute diversity (MFAD) |
功能单元间的平均配对距离之和 | 多个性状 | 否 | 否 | 否 | [43] |
| 功能分散度 Functional dispersion (FDis) |
物种偏离由相对丰度权重的中心的平均距离 | 多个性状 | 是 | 是 | 否 | [45] |
较早关于功能多样性的描述主要采用物种多样性(Species diversity, SD)及功能型组(Functional group, FG)。比如, Shannon Weiner指数, Simpson指数等可以在一定程度上作为功能多样性的间接量度。由于SD计算简便, 并被认为与生态系统功能具有较强关联, 其在早期应用较为广泛[46-49]。然而由于强调物种对生态系统功能贡献的独立性[13], 忽略了物种间功能的相似性及差异特征, SD对于群落/生态系统功能具有较低的解释效力[18, 43, 50]。此外, 将具有相似功能性状的物种划分为同一FG的方法可以使得不同FG间差异显著, 从而简化了计算过程[13]。比如, 在草地生态系统中, 依据不同生长季植物生长的活跃程度, 碳(C)、磷(P)利用途径等将植物划分为C3草本植物、C4草本植物、C3灌木、C4灌木、豆科植物及乔木等[6]。但该方法的划分过于武断, 且当前没有统一的划分标准[16], 同时其也忽略了组内物种间的功能差异及物种丰富度, 因而FG方法也不能很好的指示群落/生态系统功能多样性。
2.2 近期发展的定量化功能多样性研究方法近20年来, 随着计算机技术等的快速发展, 很多复杂的数学公式被引入到功能多样性研究中, 催生了很多定量化功能多样性指标(表 1)[7, 12-16, 28, 33]。在研究植物功能多样性与生态系统功能关系时, Walker等[12]选取植物高度, 成熟植株生物量, 比叶面积, 植株寿命及凋落物质量5个植物功能性状指标, 并提出了功能属性多样性指数(Functional attribute diversity, FAD)。相比于SD、FG方法, 该指数不需要对植物功能进行人为划分, 即既考虑了较大的物种功能差异, 也包含了较小被FG所忽略的差异, 因而其对群落/生态系统功能多样性具有较高的解释效力。该方法缺点是对于所选功能性状具有较高的敏感性, 且没有考虑物种权重, 因此其也不能很好的代表群落/生态系统功能多样性[28]。Petchey等[7]将功能多样性指数(Functional diversity, FD)定义为群落中所有物种组成功能树图的总枝长度, 通过计算物种在性状空间的离散程度进而评估物种性状互补性, 因此其可以在整个生态系统尺度上对群落/生态功能多样性进行计算。另外, Mason等[15]提出了一种功能性状变异指数(Functional diversity variance, FDvar)。与FAD类似, FD和FDvar均不需人为对植物功能进行划分, 从而有效避免了分析过程中的数据损失, 同时FDvar在计算中还考虑了物种权重的影响, 相比于之前的指标, 这两种指标更具优势性。但二者缺点是:FD在计算时会受到构建功能树图所用到的聚类方法的影响[18], 而FDvar在计算时每次只能考虑一种植物功能性状。
将基于分类学、进化分枝树图、物种间遗传距离等方法应用到功能多样性指数计算中来, 用影响群落/生态系统结构与功能的功能性状替代物种间的遗传距离, 从而可以得出一些计算功能多样性的新方法[7, 16]。基于Rao[51]将物种相对丰富度与物种间功能差异结合起来的研究方法, Botta-Dukát[16]提出了Rao二次熵系数指标(Rao′s quadratic entropy, FDQ)。相比于之前的研究方法, 该指标可同时考虑定性、定量性状, 也将物种权重纳入计算, 因而具有较高优势。但缺点是该指标在计算中会受到性状的分布以及协变量的影响, 同时在性状选取时需考虑性状相关性等问题。
与物种多样性可分解为物种丰富度和物种均匀度类似, 可将功能多样性分解为功能丰富度及功能均匀(规则)性(Functional regularity, FRO)[28]。其中功能丰富度表示群落中被物种占据的功能空间的数量。FRO表示群落功能性状空间中功能分布的均匀程度, 仅作为使用物种单个功能性状所代表的群落/生态系统功能多样性指标。总功能规则性指数(Overall Functional Regularity, OFRO)是指对于多重性状的FRO进行综合计算。FRO和OFRO方法的优点是为群落的功能均匀(规则)性提供了一种可测量连续型性状的方法, 较先前研究, 其更能反应功能多样性的某个具体方面, 更具可操作性。
很多学者认为仅采用一种指标难以代表群落/生态系统功能多样性[13-14, 28], 从而将功能多样性进行了细分:(1)功能丰富度(群落中物种占据生态位空间的大小;Functional richness, FRic);(2)功能均匀性(生态位空间中功能性状分布的均匀程度;Functional evenness, FEve);(3)功能分歧度(在何种程度上功能性状分布可以最大化整个群落的功能差异;Functional divergence, FDiv)。理论上讲, 该三种指标相互独立, 能较好指示功能多样性的不同方面。该方法为研究功能多样性提供了一种新的理论框架, 对于定量化群落/生态系统功能多样性、生态系统过程与功能等研究具有较大参考价值[14]。但其缺陷是每次只能研究一种植物功能性状, 难以代表群落/生态系统的整体功能特征, 因此在研究群落/生态系统的整体功能多样性特征时该方法有所欠缺。基于上述理论框架, Villéger等[33]建立了可用于计算多重性状的改进功能多样性三组分研究方法(Modified functional richness, MFRic;Modified functional evenness, MFEve;Modified functional divergence, MFDiv)。该方法通过将几何学中的矩阵与植物功能性状相结合, 为研究多重功能性状提供了一种新的研究框架。但其在选取功能性状时, 仍需避免性状相关性问题。另外, MFRic会随着物种丰富度的升高先升高后趋于稳定, 因此在研究中也需要保证所要研究的物种数目。
Schmera等[43]在研究功能多样性指数时提出两个标准:(1)单调性, 认为当新物种与群落中已有物种的功能性状值有所区别时, 在群落中增加该物种会使得群落功能多样性增加;(2)重合性, 认为如果在群落中增加一个与已有物种功能性状相同或相似的物种, 功能多样性不会发生变化。基于此, Schmera等[43]依据FAD建立了改进后的功能属性多样性指标(Modified functional attribute diversity, MFAD)。该方法优点是:(a)满足性状重合性标准;(b)满足性状单调性标准;(c)当使用相同功能性状时, 可进行不同群落/生态系统间的比较;(d)不需要事先对群落中所有物种均有所了解;(e)相比于FAD, MFAD对物种数量不敏感, 而对功能单元比较敏感;(f)可计算单个物种对群落/生态系统功能多样性的贡献。
Villéger等[33]的功能三组分理论框架虽然可以在多维性状空间中探索功能多样性的不同方面, 但却有很多限制条件, 比如对于定性的性状、半定量的性状、有缺失值的数据集、物种数目大于性状数目的数据集等, 该方法均有局限性。同时, 该方法也不能直接对物种个体进行权重分析[17, 33]。为解决上述问题, Laliberté和Legendre[17]提出了功能分散度指数(Functional Dispersion;FDis), 即物种个体到群落/生态系统中由所有物种组成空间中心点的平均距离。在模拟数据对该方法的检验中, 与其他功能多样性研究方法相比, 其与FDQ相关性最高(r=0.966)。FDis优点包括:(1)可计算定性、半定量性状;(2)允许数据中有缺失值;(3)可对种内性状进行权重差异分析。
3 问题与展望功能多样性是生物多样性研究中的重要组成部分[1, 34]。然而目前对于功能多样性的研究仍处于起步阶段, 虽然已出现部分理论、概念及研究方法, 但仍缺乏广泛认可[45]。同时, 现阶段对于自然群落功能多样性的研究仍相对较少, 对于功能多样性的统一化认识以及功能多样性标准化计算方法的探索仍将是未来功能生态学发展的一个主要方向。此外, 将功能多样性与生态系统功能相联系, 对生态系统功能进行定量化研究将是未来功能生态学发展的热点与难点[52]。
3.1 功能多样性研究主体目前大多学者对于功能多样性的认识主要倾向于Tilman的定义, 即功能多样性是指可影响生态系统功能的物种或有机体的性状值及范围。尽管概念中已经表明了研究主体为影响生态系统功能的有机体, 但目前关于功能多样性的绝大多数研究, 主体仍为植物群落, 与动物群落、微生物群落等的联系较少[14-17, 36]。仅考虑植物群落对生态系统功能的影响是否合理?其他生物如动物、微生物群落等对生态系统功能多样性的影响仍需进一步研究。
3.2 功能多样性计算方法功能多样性提供了一个整合离散、连续型性状的新方法, 为更精细化研究群落/生态系统过程与功能提供了有利途径。目前对于功能多样性研究主要有10种计算方法, 且均以植物功能性状为基本单元。然而在实际研究中, 关于物种选取、物种数目、性状选取、性状数目、性状权重、性状相关性等问题, 每种方法均存在一定局限性[36, 45, 52-55]。
随着群落/生态系统结构与功能的深入研究, 关于植物功能性状的研究越来越多样化, 所用数据逐渐包括定性、半定量以及定性定量的混合数据, 以及存在缺失值的数据。因此在研究群落/生态系统的功能多样性时, 须要求所选多样性指标不仅可以在很大程度上代表群落/生态系统功能, 同时也需对性状数据有较大容忍性。针对当前功能多样性研究的需要, 功能多样性指标应满足如下标准:(1)能基本代表群落/生态系统的功能多样性特征;(2)可计算定性、半定量类型的性状数据;(3)对数据没有较为严格的要求, 如允许缺失值存在, 数据非单调性等;(4)可同时计算多重功能性状数据;(5)考虑群落/生态系统中物种权重的影响;(6)可进行不同群落/生态系统间的比较;(7)可对种内性状进行权重差异分析;(8)计算简便, 易于理解, 实践性强。通过总结当前研究群落/生态系统功能多样性的10种主要研究方法, 本文认为Laliberté和Legendre[17]的功能分散性指数(FDis)以及Botta-Dukát[16]的Rao二次熵系数(FDQ)能基本满足上述标准, 在当前及未来功能生态学研究中具有较高应用潜力。但由于二者仍存在一定的局限性, 标准化功能多样性的计算方法在未来仍需进一步探索。
3.3 利用功能多样性预测生态系统功能尽管功能多样性为研究生态系统功能提供了很好的思路, 但由于方法学等的限制, 应用功能多样性探讨生态系统结构与功能的研究(如功能多样性与生态系统生产力、稳定性、资源动态关系的研究)仍显不足[36, 53-56]。如何将物种水平的功能性状与生态系统功能相联系仍将是未来功能生态学发展的一大热点与难点。He等[57]率先提出了生态系统性状(Ecosystem traits)概念, 为解决由物种尺度到生态系统尺度的功能生态学研究提供了一种新的思路。但受限于大量植物功能性状、土壤动物、微生物群落以及遥感、通量等数据的获取, 该方法也存在一些不足。此外, 由于对功能多样性的认知差异及功能性状的获取限制, 目前大多数学者在研究功能多样性对生态系统过程和功能的影响时, 仍采用物种多样性、功能组等的研究思路[45], 这对于功能多样性的应用、功能生态学的发展造成了比较大的障碍。相比于物种多样性对生态系统功能的影响, 我们有理由相信使用功能多样性的研究方法为全面认识了解生态系统过程、功能提供了一个全新的视角。
3.4 功能多样性在生态保护和恢复中的作用生态保护和恢复的关键目的是维持生态系统功能的健全及稳定[58]。近年来, 随着人类活动干预的加剧, 越来越多的生态系统面临着严峻的退化风险, 而仅基于以物种为基本单元的研究很难真正体现生态系统的资源动态及其稳定程度。功能多样性, 通过整合植物的不同功能策略, 为研究生态系统的过程与功能提供了一种较好的思路[45]。通过测定一系列自然群落的功能多样性指数, 生态学家可以进一步认识生物多样性的时空分布格局, 从而为生态保护优先级的划分(如自然保护区规划)提供理论依据[7]。比如, Devictor等[59]在使用法国高分辨率的鸟类分布数据时, 发现功能多样性的分布和物种丰富度的分布在空间上存在严重不匹配性, 虽现有保护区保护了一些物种丰富度较高的区域, 但对于功能多样性高的区域保护力度仍显不足。
到目前为止, 绝大多数的生物多样性保护工作仍集中于以物种为单元的保护[45, 60]。已有证据表明, 功能多样性受到人类活动的威胁比物种丰富度所受到的威胁更大。比如, 在研究土地利用对物种丰富度和功能多样性的影响研究中发现, 农业活动均导致了物种多样性和功能多样性的显著下降, 但功能多样性下降幅度更大[61]。可见, 仅从物种层面上对生物多样性进行保护显然难以达到维持生态系统健全功能的目的, 从功能多样性的角度出发, 不仅有助于量化生态系统的一些具体功能, 也有助于比较不同生态系统间的功能差异, 从而有助于政府等组织更有针对性的实施生态保护方案。
| [1] |
Courchamp F, Bradshaw C J A. 100 articles every ecologist should read. Nature Ecology & Evolution, 2018, 2(2): 395-401. |
| [2] |
Leps J, De Bello F, Lavorel S, Berman S. Quantifying and interpreting functional diversity of natural communities: practical considerations matter. Preslia, 2006, 78(4): 481-501. |
| [3] |
Naeem S, Duffy J E, Zavaleta E. The functions of biological diversity in an age of extinction. Science, 2012, 336(6087): 1401-1406. DOI:10.1126/science.1215855 |
| [4] |
Mouillot D, Graham N A J, Villéger S, Mason N W H, Bellwood D R. A functional approach reveals community responses to disturbances. Trends in Ecology & Evolution, 2013, 28(3): 167-177. |
| [5] |
Diaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655. |
| [6] |
Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E. The influence of functional diversity and composition on ecosystem processes. Science, 1997, 277(5330): 1300-1302. DOI:10.1126/science.277.5330.1300 |
| [7] |
Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. DOI:10.1111/j.0030-1299.2007.15559.x |
| [8] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学: 生命科学, 2015, 45(4): 325-339. |
| [9] |
Chapin Ⅲ F S, Walker B H, Hobbs R J, Hooper D U, Lawton J H, Sala O E, Tilman D. Biotic control over the functioning of ecosystems. Science, 1997, 277(5325): 500-504. DOI:10.1126/science.277.5325.500 |
| [10] |
Gillison A N. Plant functional types and traits at the community, ecosystem and world level//van der Maarel E, Franklin J, eds. Vegetation Ecology. Oxford: John Wiley & Sons, 2013: 347-386.
|
| [11] |
Walker B, Kinzig A, Langridge J. Original articles: plant attribute diversity, resilience, and ecosystem function: the nature and significance of dominant and minor species. Ecosystems, 1999, 2(2): 95-113. DOI:10.1007/s100219900062 |
| [12] |
Petchey O L, Gaston K J. Functional diversity (FD), species richness and community composition. Ecology Letters, 2002, 5(3): 402-411. DOI:10.1046/j.1461-0248.2002.00339.x |
| [13] |
Petchey O L, Hector A, Gaston K J. How do different measures of functional diversity perform?. Ecology, 2004, 85(3): 847-857. DOI:10.1890/03-0226 |
| [14] |
Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. DOI:10.1111/j.0030-1299.2005.13886.x |
| [15] |
Mason N W H, MacGillivray K, Steel J B, Wilson J B. An index of functional diversity. Journal of Vegetation Science, 2003, 14(4): 571-578. DOI:10.1111/j.1654-1103.2003.tb02184.x |
| [16] |
Botta-Dukát Z. Rao's quadratic entropy as a measure of functional diversity based on multiple traits. Journal of Vegetation Science, 2005, 16(5): 533-540. DOI:10.1111/j.1654-1103.2005.tb02393.x |
| [17] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. DOI:10.1890/08-2244.1 |
| [18] |
Poos M S, Walker S C, Jackson D A. Functional-diversity indices can be driven by methodological choices and species richness. Ecology, 2009, 90(2): 341-347. DOI:10.1890/08-1638.1 |
| [19] |
孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150-165. DOI:10.3321/j.issn:1005-264X.2007.01.019 |
| [20] |
TRY Plant Trait Database. Quantifying and scaling global plant trait diversity: Database version 5 online (2019-03-26).[2020-01-05]. https://www.try-db.org/TryWeb/Home.php
|
| [21] |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. Corrigendum to: new handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2016, 64(8): 715-716. DOI:10.1071/BT12225_CO |
| [22] |
Diaz S, Cabido M, Casanoves F. Plant functional traits and environmental filters at a regional scale. Journal of Vegetation Science, 1998, 9(1): 113-122. DOI:10.2307/3237229 |
| [23] |
SchuldtA, Bruelheide H, Durka W, Eichenberg D, Fischer M, Kröber W, Härdtle W, Ma K P, Michalski S G, Palm W U, Schmid B, Welk E, Zhou H Z, Assmann T. Plant traits affecting herbivory on tree recruits in highly diverse subtropical forests. Ecology Letters, 2012, 15(7): 732-739. DOI:10.1111/j.1461-0248.2012.01792.x |
| [24] |
Soudzilovskaia N A, Elumeeva T G, Onipchenko V G, Onipchenko V G, Shidakov I I, Salpagarova F S, Khubiev A B, Tekeev D K, Cornelissen J H C. Functional traits predict relationship between plant abundance dynamic and long-term climate warming. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(45): 18180-18184. DOI:10.1073/pnas.1310700110 |
| [25] |
卜文圣, 臧润国, 丁易, 张俊艳, 阮云泽. 海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化. 生物多样性, 2013, 21(3): 278-287. |
| [26] |
Wright I J, Reich P B, Cornelissen J H C, Falster D S, Garnier E, Hikosaka K, Lamont B B, Lee W, Oleksyn J, Osada N, Poorter H, Villar R, Warton D I, Westoby M. Assessing the generality of global leaf trait relationships. New Phytologist, 2005, 166(2): 485-496. DOI:10.1111/j.1469-8137.2005.01349.x |
| [27] |
Asanok L, Marod D, Duengkae P, Pranmongkol U, Kurokawa H, Aiba M, Katabuchi M, Nakashizuka T. Relationships between functional traits and the ability of forest tree species to reestablish in secondary forest and enrichment plantations in the uplands of northern Thailand. Forest Ecology and Management, 2013, 296: 9-23. DOI:10.1016/j.foreco.2013.01.029 |
| [28] |
Mouillot D, Mason W H N, Dumay O, Wilson J B. Functional regularity: a neglected aspect of functional diversity. Oecologia, 2005, 142(3): 353-359. DOI:10.1007/s00442-004-1744-7 |
| [29] |
Pavoine S, Vela E, Gachet S, De Bélair G, Bonsall M B. Linking patterns in phylogeny, traits, abiotic variables and space: a novel approach to linking environmental filtering and plant community assembly. Journal of Ecology, 2011, 99(1): 165-175. DOI:10.1111/j.1365-2745.2010.01743.x |
| [30] |
Westoby M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil, 1998, 199(2): 213-227. DOI:10.1023/A:1004327224729 |
| [31] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [32] |
Gillison A. A generic, computer-assisted method for rapid vegetation classification and survey: tropical and temperate case studies. Conservation Ecology, 2002, 6(2): 3. |
| [33] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. DOI:10.1890/07-1206.1 |
| [34] |
Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741-758. DOI:10.1111/j.1461-0248.2006.00924.x |
| [35] |
Lavorel S, Grigulis K, McIntyre S, Williams N S G, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A. Assessing functional diversity in the field-methodology matters!. Functional Ecology, 2008, 22(1): 134-147. |
| [36] |
Zhang H, Chen H Y, Lian J Y, John R, Li R H, Liu H, Ye W H, Berninger F, Ye Q. Using functional trait diversity patterns to disentangle the scale-dependent ecological processes in a subtropical forest. Functional Ecology, 2018, 32(5): 1379-1389. DOI:10.1111/1365-2435.13079 |
| [37] |
Garnier E, Cortez J, Billès G, Navas M L, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint J P. Plant functional markers capture ecosystem properties during secondary succession. Ecology, 2004, 85(9): 2630-2637. DOI:10.1890/03-0799 |
| [38] |
Bolnick D I, Amarasekare P, Araújo M S, Bürger R, Levine J M, Novak M, Rudolf V H M, Schreiber S J, Urban M C, Vasseur D A. Why intraspecific trait variation matters in community ecology. Trends in Ecology & Evolution, 2011, 26(4): 183-192. |
| [39] |
Laughlin D C, Joshi C, van Bodegom P M, Bastow Z A, Fulé P Z. A predictive model of community assembly that incorporates intraspecific trait variation. Ecology Letters, 2012, 15(11): 1291-1299. DOI:10.1111/j.1461-0248.2012.01852.x |
| [40] |
Plourde B T, Boukili V K, Chazdon R L. Radial changes in wood specific gravity of tropical trees: inter-and intraspecific variation during secondary succession. Functional Ecology, 2015, 29(1): 111-120. DOI:10.1111/1365-2435.12305 |
| [41] |
Kichenin E, Wardle D A, Peltzer D A, Morse C W, Freschet G T. Contrasting effects of plant inter-and intraspecific variation on community-level trait measures along an environmental gradient. Functional Ecology, 2013, 27(5): 1254-1261. DOI:10.1111/1365-2435.12116 |
| [42] |
Violle C, Enquist B J, McGill B J, Jiang L, Albert C H, Hulshof C, Jung V, Messier J. The return of the variance: intraspecific variability in community ecology. Trends in Ecology & Evolution, 2012, 27(4): 244-252. |
| [43] |
Schmera D, Erös T, Podani J. A measure for assessing functional diversity in ecological communities. Aquatic Ecology, 2009, 43(1): 157-167. DOI:10.1007/s10452-007-9152-9 |
| [44] |
Mouchet M A, Villéger S, Mason N W H, Mouillot D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 2010, 24(4): 867-876. DOI:10.1111/j.1365-2435.2010.01695.x |
| [45] |
Cadotte M W, Carscadden K, Mirotchnick N. Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 2011, 48(5): 1079-1087. DOI:10.1111/j.1365-2664.2011.02048.x |
| [46] |
Naeem S, Naeem S, Thompson L J, Lawler S P, Lawton J H, Woodfin R M. Empirical evidence that declining species diversity may alter the performance of terrestrial ecosystems. Philosophical Transactions of the Royal Society B: Biological Sciences, 1995, 347(1321): 249-262. DOI:10.1098/rstb.1995.0025 |
| [47] |
Grover JP. Resource Competition. Berlin: Springer Science and Business Media, 1997.
|
| [48] |
Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E. The influence of functional diversity and composition on ecosystem processes. Science, 1997, 277(5330): 1300-1302. DOI:10.1126/science.277.5330.1300 |
| [49] |
Tilman D. The ecological consequences of changes in biodiversity: a search for general principles. Ecology, 1999, 80(5): 1455-1474. |
| [50] |
Li R H, Zhu S D, Chen H Y H, John R, Zhou G Y, Zhang D Q, Zhang Q M, Ye Q. Are functional traits a good predictor of global change impacts on tree species abundance dynamics in a subtropical forest?. Ecology Letters, 2015, 18(11): 1181-1189. DOI:10.1111/ele.12497 |
| [51] |
Rao C R. Diversity and dissimilarity coefficients: a unified approach. Theoretical Population Biology, 1982, 21(1): 24-43. DOI:10.1016/0040-5809(82)90004-1 |
| [52] |
Gower J C. A general coefficient of similarity and some of its properties. Biometrics, 1971, 27(4): 857-871. DOI:10.2307/2528823 |
| [53] |
Mason N W H, de Bello F, Mouillot D, Pavoine S, Dray S. A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. Journal of Vegetation Science, 2013, 24(5): 794-806. DOI:10.1111/jvs.12013 |
| [54] |
Song Y T, Wang P, Li G D, Zhou D W. Relationships between functional diversity and ecosystem functioning: a review. Acta Ecologica Sinica, 2014, 34(2): 85-91. DOI:10.1016/j.chnaes.2014.01.001 |
| [55] |
Li S P, Cadotte M W, Meiners S J, Hua Z S, Jiang L, Shu W S. Species colonisation, not competitive exclusion, drives community overdispersion over long-term succession. Ecology Letters, 2015, 18(9): 964-973. DOI:10.1111/ele.12476 |
| [56] |
Laureto L M O, Cianciaruso M V, Samia D S M. Functional diversity: an overview of its history and applicability. Natureza & Conservação, 2015, 13(2): 112-116. |
| [57] |
He N P, Liu C C, Piao S L, Sack L, Xu L, Luo Y Q, He J S, Han X G, Zhou G S, Zhou X H, Lin Y, Yu Q, Liu S R, Sun W, Niu S L, Li S G, Zhang J H, Yu G R. Ecosystem traits linking functional traits to macroecology. Trends in Ecology & Evolution, 2019, 34(3): 200-210. |
| [58] |
Thorpe A S, Stanley A G. Determining appropriate goals for restoration of imperilled communities and species. Journal of Applied Ecology, 2011, 48(2): 275-279. DOI:10.1111/j.1365-2664.2011.01972.x |
| [59] |
Devictor V, Mouillot D, Meynard C, Jiguet F, Thuiller W, Mouquet N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world. Ecology Letters, 2010, 13(8): 1030-1040. |
| [60] |
Mace G M, Gittleman J L, Purvis A. Preserving the tree of life. Science, 2003, 300(5626): 1707-1709. DOI:10.1126/science.1085510 |
| [61] |
Flynn D F B, Gogol-Prokurat M, Nogeire T, Molinari N, Richers B T, Lin B B, Simpson N, Mayfield M M, DeClerck F. Loss of functional diversity under land use intensification across multiple taxa. Ecology Letters, 2009, 12(1): 22-33. DOI:10.1111/j.1461-0248.2008.01255.x |