 2021, Vol. 41
2021, Vol. 41文章信息
- 张毅锋, 汤孟平
- ZHANG Yifeng, TANG Mengping
- 天目山常绿阔叶林空间结构动态变化特征
- Analysis on spatial structure dynamic characteristics of evergreen broad-leaved forest in Tianmu Mountain
- 生态学报. 2021, 41(5): 1959-1969
- Acta Ecologica Sinica. 2021, 41(5): 1959-1969
- http://dx.doi.org/10.5846/stxb201912312841
-
文章历史
- 收稿日期: 2019-12-31
- 网络出版日期: 2021-01-04





2. 浙江农林大学环境与资源学院, 临安 311300
2. School of Environmental and Resources Science, Zhejiang Agriculture and Forestry University, Lin'an 311300, China
森林空间结构决定林木间的竞争关系及其空间生态位, 很大程度上决定森林的稳定性、发展的可能性和经营空间的大小[1]。在森林经营中, 充分考虑森林空间结构的影响是维持和改善森林质量的基本前提[2]。一般认为, 森林空间结构主要包括林木空间分布格局、树种混交和林木竞争3方面[3-6]。林木空间分布格局描述林木在空间的分布形式, 包括聚集分布、随机分布和均匀分布。树种混交描述不同树种在群落空间上的配置。林木竞争描述不同大小树木之间的竞争态势[7]。随着森林可持续经营对精确信息的需求增加, 森林空间结构研究呈日益增加趋势[8-11]。常绿阔叶林是亚热带地区最复杂、生产力最高、生物多样性最丰富的地带性植被类型之一, 能保护森林生态系统的平衡和相对稳定, 对保护环境、维持全球碳循环平衡和人类可持续发展等都具有重要作用[12]。因此, 掌握常绿阔叶林群落空间结构动态变化规律, 对亚热带地区环境治理、生物多样性保护和维持均具有重要意义。
目前, 国内外学者已对常绿阔叶林的空间结构开展了大量研究, 最初集中在群落优势树种的空间分布格局、种内种间竞争等单一方面的空间结构分析[13-17], 随着混交度概念的提出和应用[18-19], 学界逐渐开展对常绿阔叶林的林木空间分布格局、树种混交和林木竞争等多种空间结构特征研究[20-24]。但是, 现有研究主要采用一次性调查数据, 静态地分析常绿阔叶林的空间结构特征。或者采用空间代替时间的方法, 分析常绿阔叶林优势树种的分布格局动态变化特征[14, 16]。由于空间代替时间的方法缺乏长期定位观测数据, 其研究结果存在一定的局限性[25-26]。近年来, 建立永久固定样地进行长期观测的方法被认为是研究群落结构动态较为可靠的方法, 国内外已开始建立固定样地来系统研究森林生态系统的结构动态[27-28]。然而, 利用固定样地长期观测数据, 分析常绿阔叶林的空间结构动态变化尚少见报道[29-30]。
本研究以浙江省天目山国家级自然保护区内的常绿阔叶林为研究对象, 设置固定样地, 基于2005年、2010年和2015年3期调查数据, 利用优势度分析法、聚集指数、树种多样性混交度和Hegyi竞争指数, 分析群落和优势树种的空间结构动态变化特征, 旨在为常绿阔叶林的可持续经营提供依据。
1 研究区概况研究区位于浙江省杭州市临安区境内的浙江天目山国家级自然保护区(119°23′47″—119°28′27″E, 30°18′30″—30°24′55″N)。该自然保护区总面积1050hm2, 主峰海拔1506m。年平均气温8.8—14.8℃, ≥10℃年积温2500—5100℃, 年降水量1390—1870mm, 相对湿度76%—81%, 主要受海洋暖湿气候影响, 具有中亚热带向北亚热带过渡的特征。保护区内地势陡峭, 海拔上升速度快, 气候差异明显, 植被的分布有着明显的垂直界限, 自山麓到山顶垂直带谱为:海拔870m以下为常绿阔叶林;870—1100m为常绿、落叶阔叶混交林;1100—1380m为落叶阔叶林;1380—1506m为落叶矮林。其中, 该地区常绿阔叶林乔木树种以壳斗科常绿树种为主, 例如青冈(Cyclobalanopsis glauca)、细叶青冈(Cyclobalanopsis gracilis)、短尾柯(Lithocarpus brevicaudatus)等, 此外还常混生有常绿针叶和落叶阔叶树种, 例如杉木(Cunninghamia lanceolata)、榧树(Torreya grandis)、枫香(Liquidambar formosana)、榉树(Zelkova serrata)等[14]。
2 研究方法 2.1 样地设置与调查2005年7—8月, 在自然保护区内选择有代表性的地段, 设置常绿阔叶林固定样地, 样地大小100m×100m。采用相邻网格调查法把固定样地划分为100个10m×10m的调查单元, 在每个单元内进行每木调查, 记录树种、胸径、树高、活枝下高、冠幅等因子数据, 以样地西南角为原点, 采用激光对中全站仪(徕卡TCR702Xrange)测定每株树木的三维坐标。2010年7月和2015年7月, 先后对该固定样地进行复查。
2.2 优势树种的确定优势树种对群落结构和环境的形成和改变有着显著控制作用[14], 采用Ohsawa优势度分析法可以确定群落优势树种[31], 公式如下:
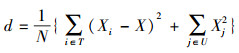
|
(1) |
式中, Xi为前位树种(T)的相对胸高断面积, X为以优势树种数量确定的优势树种理想百分比, Xj为剩余种(U)的百分比, N为总种数。假设群落只有1个优势树种, 那么优势树种的理想百分比为100%。如果有2个优势树种, 那么理想百分比为50%, 依次类推, 分别计算d值。当d值取到最小值时, 前位种的种数(T)即为群落优势种数。
2.3 边缘矫正和森林空间结构单元的确定为了防止样地界外可能是样地内树木的相邻木的情况, 采用8邻域法对样地进行边缘矫正, 只将原样地林木作为对象木进行空间结构指数的计算[32]。
森林空间结构单元是森林空间结构分析的基本单位, 它由样地内任意一株林木作为对象木及其n株相邻木所构成。相邻木数n的取值决定空间结构单元的大小与形状。传统构建空间结构单元的方法是固定n的值为3或4, 但由于现实森林中对象木与其相邻木具有多种分布情形, 任何固定的n值均不能反映真实情况, 会造成空间结构指数的有偏估量[32]。而汤孟平等引入Voronoi图来确定空间结构单元, 可以克服这个问题, 因为根据Voronoi图的特征, 任一对象木的相邻木是确定的[7]。因此, 本研究采用Voronoi图确定空间结构单元, 计算空间结构指数。
2.4 空间结构指数 2.4.1 聚集指数聚集指数可以分析林木的空间分布格局,指的是最近邻单株距离的平均值与随机分布下的期望平均距离之比[33],公式如下:
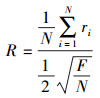
|
(2) |
式中,R为聚集指数;ri为第i株树木到其最近邻木的距离(m);N为样地内的对象木的总株数(株);F为样地面积(m2)。
在(2)式中,R∈[0,2.1491],若R>1,则群落呈均匀分布趋势;若0≤R<1,群落呈聚集分布趋势;若R=1,则群落呈随机分布趋势。在聚集分布(0≤R<1)情况下,根据R的取值范围划分聚集程度,R取值在[0, 0.25), [0.25, 0.5), [0.5, 0.75), [0.75, 1)区间内,分别表示强度、中度、低度、弱度4个林分聚集等级。
根据林分聚集指数公式,解析出单木聚集指数公式,公式如下:
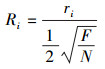
|
(3) |
根据Ri的取值范围划分为以下9个等级:
(1) 聚集分布
Ri取值在[0, 0.25), [0.25, 0.5), [0.5, 0.75), [0.75, 1)区间内,分别表示强聚集、中聚集、低聚集、弱聚集4个等级。
(2) 随机分布Ri=1
(3) 均匀分布
Ri取值在(1, 1.38), [1.38, 1.76), [1.76, 2.14), [2.14, ∞)区间内,分别表示弱均匀、低均匀、中均匀、强均匀4个等级。
2.4.2 树种多样性混交度树种多样性混交度反映对象木与相邻木之间, 以及相邻木相互之间的树种空间隔离程度, 计算公式为[4]:
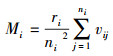
|
(4) |
式中, Mi为样地内林木i处的点混交度;ni为对象木i的相邻木的株数(株);ri为对象木i的ni株相邻木中不同树种个数(个);当对象木i与第j株相邻木属同一树种时, vij=0。当对象木i与第j株相邻木属于不同树种时, vij=1。
林分混交度是点混交度的平均值, 公式为:
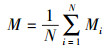
|
(5) |
式中, M是林分混交度;N为样地的对象木的总株数(株)。
M的取值在[0, 0.25), [0.25, 0.5), [0.5, 0.75), [0.75, 1]区间内, 分别表示弱度、低度、中度、强度4个混交等级。
2.4.3 Hegyi竞争指数采用Hegyi竞争指数描述林木与其相邻木之间的竞争态势, 计算公式为[15, 34]:
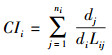
|
(6) |
式中, CIi为林木i的点竞争指数;Lij为对象木i与竞争木j之间的距离(m);di为对象木i的胸径(cm), dj为竞争木j的胸径(cm);ni为对象木i的竞争木的株数(株)。
为了反映群落的平均竞争水平, 取林木点竞争指数的平均值作为林分竞争指数[35-36], 计算公式为:
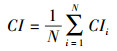
|
(7) |
式中, CI为林分竞争指数;N为样地对象木的总株数(株)。
CI的取值在[0, 6), [6, 12), [12, 18), [18, ∞)区间内, 分别表示弱度、低度、中度、强度4个竞争等级。
2.5 空间结构动态变化分析空间结构动态变化分析就是分析空间结构特征在时间序列上的变化, 具体方法:首先在Excel中对3期样地调查数据进行预处理, 利用Python编程语言结合ArcMap 10.2中Voronoi图分析功能, 计算样地内每株树木的空间结构指数。再通过SPSS 20软件的Kolmogorov-Smirnov检验确定数据是否服从正态分布, 若不服从正态分布, 则使用Wilcoxon-Mann-Whitney秩和检验法, 分析不同调查年份空间结构指数之间的差异性。
2.5.1 Kolmogorov-Smirnov检验Kolmogorov-Smirnov检验(以下简称K-S检验)可检验经验分布是否和理论分布一致。假设F(X)为理论分布累积函数, Fn(X) 表示样本累积分布函数。定义统计量[37]。
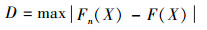
|
(8) |
可用统计量D衡量Fn(X) 与F(X) 的差别。根据给定显著水平α和样本容量n, 确定单样本K-S检验临界值D(α, n), 如果D≥ D(α, n), 则说明经验分布与理论分布有显著差异。
2.5.2 Wilcoxon-Mann-Whitney秩和检验Wilcoxon-Mann-Whitney秩和检验(以下简称W-M-W检验)是检验两个独立样本是否来自同一个总体的非参数检验方法。其方法是把样本(X1, X2, …, Xn)和(Y1, Y2, …, Yn)混合, 并按照升序进行排序, 每个数据有一个对应的秩。分别计算两个样本的秩和记为Wx和Wy, 根据Wx和Wy计算W-M-W检验统计量, 公式如下:
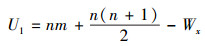
|
(9) |
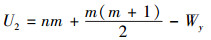
|
(10) |
式中, n和m分别是样本X和样本Y的样本容量。
选择U1和U2中较小者作为最终的检验统计量U。对于两个大样本的W-M-W检验统计量, 其抽样分布接近正态分布, 对应均值和标准差为:
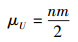
|
(11) |
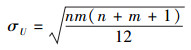
|
(12) |
相应的Z统计量为:
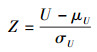
|
(13) |
在给定0.05显著性水平下, 应用标准正态分布进行检验。若原假设Mx=My (Mx和My分别是样本X和样本Y的中位数)成立, 则两组数据总体分布相同;若原假设不成立, 则两组数据具有较大的差异性。
3 结果与分析 3.1 群落空间结构动态变化 3.1.1 群落结构因子统计分析各调查年份林分空间结构指数计算结果及林分株数和树种数统计结果见表 1。可知, 常绿阔叶林林木株数呈逐渐减少趋势, 从2005年的3402株, 减少至2015年的2820株, 减少了582株, 降幅为17.11%。同时, 有13个树种退出群落, 这些树种株数均在1—3株之间, 并不会对群落的物种多样性产生较大影响。2005—2015年, 林分聚集指数处于[0.5, 0.75)区间范围内, 群落呈低度聚集分布格局;林分混交度处于[0.5, 0.75)区间范围内, 群落呈中度混交状态;林分竞争指数处于[12, 18)区间范围内, 群落呈中度竞争态势。
3.1.2 群落林木分布格局变化2005—2015年, 常绿阔叶林林分聚集指数在0.6728—0.6832之间, 3次样地调查群落均表现为低度聚集分布(表 1)。但是, 林分聚集指数存在不断升高的趋势, 有必要对这一趋势进行显著性检验。K-S检验结果表明, 各年林木聚集指数均不符合正态分布(P<0.05)。进而采用W-M-W检验分析各年林分聚集指数之间的差异性。结果表明, 各年林分聚集指数之间差异显著(P<0.05), 即2005—2015年林分聚集指数呈持续上升趋势, 群落聚集程度不断下降。前5年林分聚集指数提高0.52%, 后5年林分聚集指数提高1.02%, 10年间林分聚集指数提高了1.55%, 后5年分布格局的变化程度大于前5年。
| 调查年份 Survey year |
株数 Number of trees |
树种数 Number of tree species |
聚集指数 Aggregation index |
树种多样性混交度 Tree species diversity mingling |
Hegyi竞争指数 Hegyi competition index |
| 2005 | 3402 | 100 | 0.6728 | 0.5259 | 15.2901 |
| 2010 | 3254 | 98 | 0.6763 | 0.5268 | 15.1770 |
| 2015 | 2820 | 87 | 0.6832 | 0.5252 | 14.3175 |
绘制各年指数的频率分布图, 可以更具体地分析指数的分布与变化特征。各年聚集指数频率分布图见图 1。可知, 各年林木聚集指数的频率分布特征基本相同, 70%—71.11%林木表现为聚集分布, 28.89%—30%林木表现为均匀分布。强聚集等级(0≤Ri<0.25)的分布频率值最大, 频率范围在42.23%—44.12%, 其次是弱均匀等级(1<Ri<1.38), 频率范围在12.52%—13.33%。分布频率最小值出现在强均匀等级(2.14≤Ri), 频率范围在3.32%—3.48%。2005—2015年, 低聚集、弱均匀等级的分布频率增加, 强聚集等级的分布频率降低, 而中聚集、弱聚集、低均匀、中均匀和强均匀等级的分布频率基本保持稳定。其中, 各分布等级变化幅度最大的是强聚集分布, 从44.12%降低至42.23%, 其次是低聚集分布, 从8.47%增加至9.33%。

|
| 图 1 各年聚集指数频率分布图 Fig. 1 Frequency distribution histogram of aggregate index for each year |
2005—2015年, 常绿阔叶林林分混交度在0.5252—0.5268之间, 3次样地调查群落均表现为中度混交状态(表 1)。对林分混交度先升后降的趋势进行显著性检验, 通过K-S检验发现, 各年林木混交度均不符合正态分布(P<0.05)。进而采用W-M-W检验分析不同年份林分混交度间的差异性, 结果表明, 各年林分混交度之间无显著差异(P>0.05), 即2005—2015年林分混交度没有发生显著变化, 群落混交程度基本保持稳定。
各年混交度频率分布图见图 2。可知, 各年林木混交度的频率分布特征基本相同, 中度混交等级(0.5≤ Mi<0.75)的分布频率最高, 频率范围在35.83%—36.45%, 其次是低度混交等级(0.25≤ Mi<0.5)的分布频率, 频率范围在30.97%—31.86%。分布频率最小值出现在弱度混交等级(0≤ Mi<0.25), 频率范围在12.93%—13.37%。2005—2010年, 低度混交等级的分布频率降低0.89%, 中度混交等级的分布频率增加0.56%, 弱度和强度混交等级的分布频率基本保持稳定。2010—2015年, 各混交等级分布频率基本保持稳定。2005—2015年, 低度混交等级的分布频率降低0.62%, 中度混交等级的分布频率增加0.62%, 弱度和强度混交等级的分布频率基本保持稳定。

|
| 图 2 各年混交度频率分布图 Fig. 2 Frequency distribution histogram of mingling for each year |
2005—2015年, 常绿阔叶林林分竞争指数在14.3175—15.2901之间, 3次样地调查群落均表现为中度竞争态势(表 1)。对林分竞争指数不断降低的趋势进行显著性检验, K-S检验结果显示, 各年林木竞争指数均不符合正态分布(P<0.05), 进而采用W-M-W检验分析各年林分竞争指数间的差异性, 结果表明, 2005—2015年林分竞争指数变化差异显著(P<0.05), 其中, 前5年林分竞争指数无显著差异(P>0.05), 后5年林分竞争指数差异显著(P<0.05)。由此可知, 2005—2015年群落竞争强度降低, 10年间林分竞争指数下降了6.36%。
各年竞争指数频率分布图见图 3。可知, 各年林木竞争指数的频率分布特征基本相同, 最大频率值都出现在弱度竞争等级(0≤CIi<6)中, 范围在32.51%—35.99%。其次是低度(6≤CIi<12)和强度竞争等级(18≤CIi), 频率范围分别在25.46%—27.21%和23.94%—25.96%。2005—2010年, 强度竞争等级的分布频率降低, 低度和弱度竞争等级的分布频率增加, 中度竞争等级的分布频率基本保持稳定, 分布频率变化幅度最大是低度竞争等级, 从25.78%增加至27.21%。2010—2015年, 除弱度竞争等级的分布频率增加外, 其余竞争等级分布频率均降低, 分布频率变化幅度最大的是弱度竞争等级, 从33.10%增加至35.99%。

|
| 图 3 各年竞争指数频率分布图 Fig. 3 Frequency distribution histogram of Hegyi competition index for each year |
各年优势树种确定情况如表 2所示。2005、2010、2015年群落优势种数分别是14、14、13个, 期间只有黄檀和金钱松进出过群落优势种组成, 细叶青冈一直处于最优势地位, 是群落的建群种。可见, 2005—2015年群落优势种组成变化不大, 保持着多优势种结构特征。将3次调查均确定为优势树种的13个树种作为优势树种研究对象, 为便于比较分析, 将这13个优势树种按树种类别分为常绿阔叶树种、常绿针叶树种和落叶阔叶树种3个优势树种组, 分析其空间结构动态变化特征。常绿阔叶树种有:细叶青冈、短尾柯、青冈、小叶青冈(Cyclobalanopsis myrsinifolia)、豹皮樟(Litsea coreana);常绿针叶树种有:杉木、榧树;落叶阔叶树种有:枫香、榉树、白栎(Quercus fabri)、黄连木(Pistacia chinensis)、蓝果树(Nyssa sinensis)、天目木姜子(Litsea auriculata)。各树种组的基本情况见表 3。可知, 各树种组的株数关系为:常绿阔叶树种>落叶阔叶树种>常绿针叶树种;平均胸径大小关系为:落叶阔叶树种>常绿针叶树种>常绿阔叶树种;群落优势地位关系为:常绿阔叶树种>落叶阔叶树种>常绿针叶树种;2005—2015年, 常绿阔叶树种的胸高断面积占比呈增长趋势, 其群落优势地位进一步提高。
| 优势树种 Dominant tree species |
2005 | 2010 | 2015 | |||||
| Ba/cm2 | % | Ba/cm2 | % | Ba/cm2 | % | |||
| 细叶青冈Cyclobalanopsis gracilis | 47315.8 | 13.6 | 49473.3 | 13.2 | 53405.2 | 14.0 | ||
| 杉木Cunninghamia lanceolata | 33863.8 | 9.8 | 34591.5 | 9.2 | 35618.8 | 9.3 | ||
| 短尾柯Lithocarpus brevicaudatus | 25786.6 | 7.4 | 29073.4 | 7.8 | 30055.8 | 7.9 | ||
| 枫香Liquidambar formosana | 25155.8 | 7.3 | 27404.2 | 7.3 | 28547.1 | 7.5 | ||
| 青冈Cyclobalanopsis glauca | 23623.3 | 6.8 | 25677.7 | 6.9 | 26521.5 | 7.0 | ||
| 白栎Quercus fabri | 22359.9 | 6.4 | 24119.4 | 6.4 | 24405.7 | 6.4 | ||
| 榧树Torreya grandis | 17867.3 | 5.2 | 19459.5 | 5.2 | 20953.9 | 5.5 | ||
| 黄连木Pistacia chinensis | 16514.2 | 4.8 | 18325.6 | 4.9 | 18080.4 | 4.7 | ||
| 小叶青冈Cyclobalanopsis myrsinifolia | 15527.8 | 4.5 | 17401.9 | 4.7 | 16359.0 | 4.3 | ||
| 蓝果树Nyssa sinensis | 10342.7 | 3.0 | 10810.4 | 2.9 | 11933.4 | 3.1 | ||
| 豹皮樟Litsea coreana | 8393.1 | 2.4 | 9351.9 | 2.5 | 10035.0 | 2.6 | ||
| 天目木姜子Litsea auriculata | 8246.2 | 2.4 | 9184.9 | 2.5 | 9977.4 | 2.6 | ||
| 榉树Zelkova serrata | 8031.0 | 2.3 | 9164.5 | 2.5 | 9901.3 | 2.6 | ||
| 黄檀Dalbergia hupeana | 8024.5 | 2.3 | — | — | ||||
| 金钱松Pseudolarix amabilis | — | 7733.6 | 2.1 | — | ||||
| Ba, 胸高断面积Basal area; -, 表示该树种非调查年份的群落优势树种 | ||||||||
| 树种组 Tree species group |
2005 | 2010 | 2015 | ||||||||
| Nt | AD/cm | Wb/% | Nt | AD/cm | Wb% | Nt | AD/cm | Wb% | |||
| 常绿阔叶树种 Evergreen broad-leaved species |
1741 | 7.354 | 34.8 | 1675 | 7.8948 | 35 | 1447 | 8.7018 | 35.3 | ||
| 常绿针叶树种 Evergreen conifer species |
207 | 13.4727 | 14.9 | 202 | 14.1458 | 14.7 | 192 | 15.0662 | 14.8 | ||
| 落叶阔叶树种 Deciduous broad-leaved species |
310 | 15.4245 | 26.1 | 295 | 16.905 | 26.5 | 264 | 18.6296 | 27.4 | ||
| Nt, 株数Number of trees; AD, 平均胸径Average DBH; Wb, 表示胸高断面积占林分总断面积的百分比 | |||||||||||
在种群的发育过程中, 其分布格局不是一成不变的, 而是随着时间发生动态变化的[38]。各树种组的分布格局动态变化如图 4所示。可见, 2005—2015年, 各树种组的聚集指数都在[0.5, 0.75)区间范围内, 均呈低度聚集分布格局。常绿阔叶和常绿针叶树种聚集指数表现出升高的趋势, 落叶阔叶树种聚集指数表现出降低的趋势, 聚集程度大小为:落叶阔叶树种>常绿阔叶树种>常绿针叶树种。对聚集指数的变化趋势进行显著性检验, K-S检验结果表明, 各树种组的各年林木聚集指数均不符合正态分布(P<0.05)。为此, 采用W-M-W检验分析同一树种组的不同年份聚集指数之间的差异性, 结果通过字母标记法在图 4上表示。结果表明:常绿阔叶树种各年聚集指数之间存在显著差异(P<0.05), 常绿针叶和落叶阔叶树种各年聚集指数之间并无显著性差异(P>0.05)。由此可知, 2005—2015年常绿阔叶优势树种的聚集程度不断降低, 常绿针叶和落叶阔叶优势树种的聚集程度保持相对稳定。

|
| 图 4 各年各树种组聚集指数直方图 Fig. 4 Histogram of aggregation index of each tree species group for each year |
树种混交程度取决于树种数量以及树种的空间配置。一般认为, 混交程度越高, 群落稳定性越强[5, 39]。同理, 种群的混交程度越高, 种群在群落中生长稳定性越高。各树种组的混交度动态变化如图 5所示。可见, 2005—2015年, 常绿阔叶树种的混交度在[0.25, 0.5)区间范围内, 呈低度混交状态, 混交度表现出先升后降的趋势;常绿针叶和落叶阔叶树种的混交度在[0.5, 0.75)区间范围内, 均呈中度混交状态, 其中, 常绿针叶树种混交度表现出先降后升的趋势, 而落叶阔叶树种表现出先升后降的趋势。各树种组混交程度大小为:常绿针叶树种>落叶阔叶树种>常绿阔叶树种。对各树种组的混交度变化趋势进行显著性检验, K-S检验结果表明, 各树种组各年林木混交度均不符合正态分布(P<0.05)。为此, 采用W-M-W检验分析同一树种组的不同年份混交度之间的差异性, 结果通过字母标记法在图 5上表示。结果表明:各树种组的各年混交度之间均没有显著性差异(P>0.05)。由此可知, 2005—2015年常绿阔叶、常绿针叶和落叶阔叶优势树种的混交程度均保持相对稳定。

|
| 图 5 各年各树种组混交度直方图 Fig. 5 Histogram of mingling of each tree species group for each year |
树种平均竞争指数能够反映出该树种个体与相邻木的平均竞争状态, 竞争指数越低(遭受到竞争压力越小), 说明树种竞争生长优势越大。各树种组的竞争强度动态变化如图 6所示。可见, 2005—2015年, 常绿阔叶树种的竞争指数在[12, 18)区间范围内, 呈中度竞争态势, 竞争生长优势一般, 竞争指数表现出不断降低的趋势;常绿针叶和落叶阔叶树种的竞争指数都在[6, 12)区间范围内, 均呈低度竞争态势, 具有较高的竞争生长优势, 其中, 常绿针叶树种的竞争指数表现出先升后降的趋势, 落叶阔叶树种的竞争指数表现出不断降低的趋势。对竞争指数的变化趋势进行显著性检验, K-S检验结果表明, 各树种组的各年竞争指数均不符合正态分布(P<0.05)。为此, 采用W-M-W检验分析同一树种组的不同年份竞争指数之间的差异性, 结果通过字母标记法在图 6上表示。结果表明:以10年间隔来看, 常绿阔叶树种的竞争指数存在显著差异(P<0.05);常绿针叶和落叶阔叶树种的各年竞争指数之间无显著差异(P>0.05)。由此可知, 2005—2015年, 常绿阔叶优势树种的竞争强度降低, 常绿针叶和落叶阔叶优势树种的竞争强度保持相对稳定。

|
| 图 6 各年各树种组竞争指数直方图 Fig. 6 Histogram of competition index of each tree species group for each year |
通过对2005—2015年天目山常绿阔叶林空间结构动态变化分析发现, 群落优势种组成变化不大, 常绿阔叶树种的优势地位进一步提升, 群落表现为低度聚集、中度混交和中度竞争状态。期间群落聚集程度和林木竞争强度降低, 树种混交程度保持稳定, 常绿阔叶优势树种与群落的空间结构变化特征基本一致。其原因是由于2005—2015年群落中17.11%的林木枯损死亡, 其中, 常绿阔叶、常绿针叶和落叶阔叶优势树种死亡木分别占死亡木总数的50.2%、2.6%、7.9%。死亡木多与其周围林木聚集生长, 聚集指数均值为0.7091, 导致活立木间距扩大。另外, 常绿阔叶优势树种的林木死亡被认为可能是其生存策略或适应机制[40]。因为常绿阔叶优势树种多为壳斗科植物, 具有聚集繁殖特性, 幼小林木聚集生长, 随着树木年龄增长, 对环境资源的需求加剧, 导致种内种间竞争作用加剧, 从而造成林木自疏或它疏死亡[16], 其死亡木数据显示, DBH<10cm的死亡木占89.1%, 平均竞争指数为18.2813, 平均混交度为0.4263, 证实死亡木确实多为遭受强度竞争压迫且同种聚生的小径级林木, 这一现象与谭雪莲等的研究结果一致[41]。吴鞠等在研究林分混交度对长白山天然风景林树木形态的影响时指出, 当树木处于低混交时, 提高混交度能缓和相邻植株间生态竞争, 优化林分空间利用效率[42]。因此, 近自然经营过程中, 有必要提高局部林木混交度, 同时小树与大树保持较远距离, 降低竞争压迫。值得一提的是汤孟平等在研究常绿阔叶林混交度时指出, 壳斗科植物的聚集繁殖方式虽然加剧了青冈等常绿阔叶优势树种的种内竞争, 降低了其树种混交度, 但也正是这种繁殖方式使得青冈等常绿树种在群落中确立优势地位, 对群落的演替产生重要影响[32]。结合本研究发现, 天目山常绿阔叶林经过10年的演变, 群落中有部分林木死亡, 群落林木分布格局和竞争态势均发生变化, 但混交度依旧保持稳定, 说明自然状态下常绿阔叶林群落树种混交度的变化是一个漫长的过程。因此, 在常绿阔叶林经营或人工林近自然改造过程中, 不可忽视空间结构变化规律和种群生物学特性, 急于通过1次优化改造达到理想结构或盲目追求混交度为1的最高混交状态。应根据群落演替阶段和优势种群生物学特性, 设置阶段性空间结构优化目标, 逐步促进群落进展演替。
林木空间分布格局是多种环境因子共同作用的结果, 也是影响林木发展的重要因素[38]。一般情况下, 天然混交林随着群落的演替, 林木空间分布格局趋于随机分布[43]。已有许多学者通过分析群落分布格局来推测群落的演替阶段及趋势。如肖欣等在对井冈山国家自然保护区内3种典型常绿阔叶林群落空间结构特征分析时发现, 丝栗栲、罗浮栲和红楠群落分布格局都接近随机分布, 由此认为各群落处在演替中后期向顶级群落演替的阶段[23]。陈婷婷等在对武夷山常绿阔叶林空间结构特征分析时发现, 群落呈强度混交的轻度聚集分布格局, 具有较高的树种异质性, 推断群落演替可能已达中后期, 将继续向顶级群落演替[24]。张家诚等在对典型顶级森林群落和群落演替过程分析中也指出, 顶级群落发育过程中, 各优势树种的种群分布表现为由集群向随机的波动性扩散, 最终较为稳定地呈现随机分布格局[44]。本研究中的天目山常绿阔叶林群落具有较高的树种多样性和稳定的种间相互隔离关系, 群落整体与常绿阔叶优势树种趋于随机分布, 其空间结构特征与武夷山和井冈山常绿阔叶林基本接近[23-24], 主要差别在树种混交度上, 细叶青冈、青冈和短尾柯等主要常绿阔叶优势树种的聚集繁殖更新方式是该群落混交度偏低的重要原因[45]。而且, 根据前人对该群落不同优势树种组年龄结构分析表明, 常绿阔叶优势树种年龄结构是稳定性, 落叶优势树种天然更新困难, 其年龄结构是衰退型[14]。因此, 认为该群落正处于进展演替过程中, 可能已达到演替阶段的中后期, 随着演替的进行, 落叶阔叶树种逐渐退出群落, 常绿阔叶树种优势地位不断提高, 群落林木分布聚集程度降低, 逐渐向随机分布转变, 群落最终演替为典型亚热带常绿阔叶林顶级群落。
大型固定监测样地为研究群落演替和种群动态等生态学规律和现象提供了绝佳的机会, 是生态学研究中最具有活力的领域之一[29]。通过在天目山自然保护区内建立固定样地长期监测, 分析典型植被类型常绿阔叶林的空间结构动态变化特征, 不仅可以为亚热带常绿阔叶林的空间结构研究提供借鉴和参考, 还能为常绿阔叶林恢复与重建提供理论依据。但目前有关常绿阔叶林的空间结构动态监测研究较少, 未来建立以空间结构为目标的常绿阔叶林长期固定监测样地网, 对理解其结构与功能关系, 揭示群落形成机理及其稳定性与演替规律等将有重要推动作用。
| [1] |
惠刚盈, 胡艳波. 混交林树种空间隔离程度表达方式的研究. 林业科学研究, 2001, 14(1): 23-27. DOI:10.3321/j.issn:1001-1498.2001.01.004 |
| [2] |
Baskent E Z, Keles S. Spatial forest planning: a review. Ecological Modelling, 2005, 188(2/4): 145-173. |
| [3] |
Aguirre O, Hui G Y, von Gadow K, Jiménez J. An analysis of spatial forest structure using neighbourhood-based variables. Forest Ecology and Management, 2003, 183(1/3): 137-145. |
| [4] |
汤孟平, 唐守正, 雷相东, 周国模, 谢志新. 两种混交度的比较分析. 林业资源管理, 2004(4): 25-27. DOI:10.3969/j.issn.1002-6622.2004.04.007 |
| [5] |
汤孟平, 唐守正, 雷相东, 李希菲. 林分择伐空间结构优化模型研究. 林业科学, 2004, 40(5): 25-31. DOI:10.3321/j.issn:1001-7488.2004.05.004 |
| [6] |
汤孟平. 森林空间经营理论与实践. 北京: 中国林业出版社, 2007.
|
| [7] |
汤孟平, 陈永刚, 徐文兵, 赵明水. 森林空间结构分析. 北京: 科学出版社, 2013: 21-35.
|
| [8] |
汤孟平. 森林空间结构研究现状与发展趋势. 林业科学, 2010, 46(1): 117-122. |
| [9] |
湛斌, 闫新利, 陈斌, 关庆伟. 紫金山针阔混交林空间结构特征分析. 西北林学院学报, 2016, 31(5): 224-229. DOI:10.3969/j.issn.1001-7461.2016.05.37 |
| [10] |
Szmyt J, Tarasiuk S. Species-specific spatial structure, species coexistence and mortality pattern in natural, uneven-aged Scots pine (Pinus sylvestris L.)-dominated forest. European Journal of Forest Research, 2018, 137(1): 1-16. DOI:10.1007/s10342-017-1084-x |
| [11] |
朱光玉, 徐奇刚, 吕勇. 湖南栎类天然次生林林分空间结构对灌木物种多样性的影响. 生态学报, 2018, 38(15): 5404-5412. |
| [12] |
包维楷, 刘照光, 刘朝禄, 袁亚夫, 刘仁东. 中亚热带湿性常绿阔叶次生林自然恢复15年来群落乔木层的变化动态. 植物生态学报, 2000, 24(6): 702-709. DOI:10.3321/j.issn:1005-264X.2000.06.011 |
| [13] |
金则新, 周荣满. 木荷种内与种间竞争的数量关系. 浙江林学院学报, 2003, 20(3): 259-263. |
| [14] |
汤孟平, 周国模, 施拥军, 陈永刚, 吴亚琪, 赵明水. 天目山常绿阔叶林优势种群及其空间分布格局. 植物生态学报, 2006, 30(5): 743-752. DOI:10.3321/j.issn:1005-264X.2006.05.004 |
| [15] |
汤孟平, 陈永刚, 施拥军, 周国模, 赵明水. 基于Voronoi图的群落优势树种种内种间竞争. 生态学报, 2007, 27(11): 4707-4716. DOI:10.3321/j.issn:1000-0933.2007.11.039 |
| [16] |
刘保双, 付登高, 吴晓妮, 王洪娇, 王琪, 段昌群. 滇中次生常绿阔叶林优势树种的空间格局. 生态学杂志, 2013, 32(3): 551-557. |
| [17] |
Nguyen H H, Uria-Diez J, Wiegand K. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam. Journal of Vegetation Science, 2016, 27(2): 318-327. DOI:10.1111/jvs.12361 |
| [18] |
Von Gadow K, Füldner K. Zur Methodik der Bestandesbeschreibung. Klieken: Vortrag Anlaesslich der Jahrestagung der AG Forsteinrichtung in Kliekenb, 1992. |
| [19] |
惠刚盈, 胡艳波, 赵中华. 结构化森林经营研究进展. 林业科学研究, 2018, 31(1): 85-93. |
| [20] |
黄庆丰, 宫守飞, 许剑辉. 天然落叶与常绿阔叶林林分的空间结构. 东北林业大学学报, 2011, 39(10): 1-3. DOI:10.3969/j.issn.1000-5382.2011.10.001 |
| [21] |
赵洋毅, 王克勤, 陈奇伯, 王玉杰, 王云琦, 段旭. 西南亚热带典型天然常绿阔叶林的空间结构特征. 西北植物学报, 2012, 32(1): 187-192. DOI:10.3969/j.issn.1000-4025.2012.01.030 |
| [22] |
刘益曦, 刁军. 浙江天然常绿阔叶林空间结构特征分析. 西北林学院学报, 2014, 29(4): 155-159. DOI:10.3969/j.issn.1001-7461.2014.04.28 |
| [23] |
肖欣, 丁松, 欧阳勋志. 井冈山国家自然保护区典型常绿阔叶林林分空间结构分析. 西北农林科技大学学报: 自然科学版, 2016, 44(3): 133-140. |
| [24] |
陈婷婷, 徐辉, 杨青, 陈水飞, 葛晓敏, 吴军, 崔鹏, 方炎明, 丁晖. 武夷山常绿阔叶林空间结构参数分布特征. 生态学报, 2018, 38(5): 1817-1825. |
| [25] |
周灿芳. 植物群落动态研究进展. 生态科学, 2000, 19(2): 53-59. DOI:10.3969/j.issn.1008-8873.2000.02.010 |
| [26] |
朱华, 许再富, 王洪, 李保贵. 西双版纳片断热带雨林30多年来植物种类组成及种群结构的变化. 云南植物研究, 2001, 23(4): 415-427. DOI:10.3969/j.issn.2095-0845.2001.04.002 |
| [27] |
马克平. 大型固定样地: 森林生物多样性定位研究的平台. 植物生态学报, 2008, 32(2): 237. DOI:10.3773/j.issn.1005-264x.2008.02.001 |
| [28] |
Feeley K J, Davies S J, Perez R, Hubbell S P, Foster R B. Directional changes in the species composition of a tropical forest. Ecology, 2011, 92(4): 871-882. DOI:10.1890/10-0724.1 |
| [29] |
彭辉. 基于BCI 50公顷大样地的森林空间结构特征研究[D]. 北京: 中国林业科学研究院, 2017.
|
| [30] |
赵中华, 刘文桢, 石小龙, 李安民, 郭小龙, 张弓乔, 惠刚盈. 小陇山锐齿栎天然林结构动态分析. 林业科学研究, 2015, 28(6): 759-766. DOI:10.3969/j.issn.1001-1498.2015.06.001 |
| [31] |
Ohsawa M. Differentiation of vegetation zones and species strategies in the subalpine region of Mt. Fuji. Vegetatio, 1984, 57(1): 15-52. DOI:10.1007/BF00031929 |
| [32] |
汤孟平, 周国模, 陈永刚, 赵明水, 何一波. 基于Voronoi图的天目山常绿阔叶林混交度. 林业科学, 2009, 45(6): 1-5. DOI:10.3321/j.issn:1001-7488.2009.06.001 |
| [33] |
Clark P J, Evans F C. Distance to nearest neighbor as a measure of spatial relationships in population. Ecology, 1954, 35(4): 445-453. DOI:10.2307/1931034 |
| [34] |
Hegyi F. A simulation model for managing jack-pine stands//Fries J. Growth Models for Tree and Stand Simulation. Stockholm: Royal College of Forestry, 1974: 74-90.
|
| [35] |
邹春静, 韩士杰, 张军辉. 阔叶红松林树种间竞争关系及其营林意义. 生态学杂志, 2001, 20(4): 35-38. DOI:10.3321/j.issn:1000-4890.2001.04.010 |
| [36] |
詹步清. 乳源木莲混交林种内及种间竞争研究. 福建林学院学报, 2002, 22(3): 274-277. DOI:10.3969/j.issn.1001-389X.2002.03.020 |
| [37] |
王兆军, 邹长亮. 数理统计教程. 北京: 高等教育出版社, 2014.
|
| [38] |
操国兴, 钟章成, 刘芸, 谢德体. 缙云山川鄂连蕊茶种群空间分布格局研究. 生物学杂志, 2003, 20(1): 10-12. DOI:10.3969/j.issn.2095-1736.2003.01.003 |
| [39] |
邢晖. 大兴安岭落叶松白桦混交林林分空间结构优化技术研究[D]. 哈尔滨: 东北林业大学, 2014.
|
| [40] |
Hubbell S P. Neutral theory and the evolution of ecological equivalence. Ecology, 2006, 87(6): 1387-1398. DOI:10.1890/0012-9658(2006)87[1387:NTATEO]2.0.CO;2 |
| [41] |
谭雪莲, 敬小丽, 阚蕾, 苏志尧, 张璐. 亚热带常绿阔叶林锥和木荷枯立木点格局分析. 西南林业大学学报, 2019, 39(1): 132-138. |
| [42] |
吴鞠, 陈瑜, 刘海轩, 许丽娟, 金桂香, 徐程扬. 林分密度及混交度对长白山天然风景林树木形态的影响. 林业科学, 2018, 54(12): 12-21. DOI:10.11707/j.1001-7488.20181202 |
| [43] |
胡艳波, 惠刚盈, 戚继忠, 安慧君, 郝广明. 吉林蛟河天然红松阔叶林的空间结构分析. 林业科学研究, 2003, 16(5): 523-530. DOI:10.3321/j.issn:1001-1498.2003.05.002 |
| [44] |
张家城, 陈力, 郭泉水, 聂道平, 白秀兰, 蒋有绪. 演替顶极阶段森林群落优势树种分布的变动趋势研究. 植物生态学报, 1999, 23(3): 256-268. DOI:10.3321/j.issn:1005-264X.1999.03.008 |
| [45] |
方国景. 天目山常绿阔叶林空间结构研究[D]. 杭州: 浙江林学院, 2008.
|