 2021, Vol. 41
2021, Vol. 41文章信息
- 薛娟, 魏雪, 何先进, 吴鹏飞
- XUE Juan, WEI Xue, HE Xianjin, WU Pengfei
- 高寒草甸生态系统蚁丘对小型土壤节肢动物群落的影响
- Effects of ant-hills on the community structure of soil microarthropods in an alpine meadow ecosystem
- 生态学报. 2021, 41(4): 1613-1624
- Acta Ecologica Sinica. 2021, 41(4): 1613-1624
- http://dx.doi.org/10.5846/stxb202003030395
-
文章历史
- 收稿日期: 2020-03-03
- 网络出版日期: 2020-12-24





2. 重庆大学环境与生态学院, 重庆 400044
2. College of Environment and Ecology, Chongqing University, Chongqing 400044, China
小型土壤节肢动物作为消费者和分解者, 是草地生态系统的重要的组成成分, 对草地生态系统的物质循环和能量流动起着重要的调控作用[1]。已有研究表明, 小型土壤节肢动物的活动能促进有机质分解、维持土壤结构、指示植被变化、改变土壤理化性质[2-4]。草地生态系统小型土壤节肢动物群落受小型兽类、放牧等因素的影响[5-6]。因此, 小型土壤节肢动物通常也被视为草地生态系统响应干扰的指示类群[7]。
蚂蚁是草地生态系统中分布最广泛的昆虫类群之一, 具有庞大的生物量和改造环境的能力, 被称为“生态系统工程师”[8-9]。蚂蚁的筑丘行为作为草地生态系统不可忽视的干扰因子, 能改变局部的环境特征, 引起生境的异质性[10]。目前关于蚁丘的研究主要集中在对土壤理化性质、植物群落多样性、微生物功能多样性和土壤呼吸等方面[11-19]。例如, 蚂蚁通过搬运、取食、消化及排泄有机残体, 影响了土壤养分的重新分配, 从而对土壤理化性质产生显著影响[11-13], 进一步改变了植物群落的高度、盖度、生物量[14-15]和结构[16]等。蚁丘土能维持较高的土壤温度, 使得土壤微生物的数量和活性有所升高, 从而影响了土壤微生物群落功能多样性[17-18]。此外, 蚂蚁筑巢能够降低土壤容重, 增加土壤孔隙度、持水性和通透性等, 从而对土壤呼吸产生重要影响[19]。而已有的研究表明土壤理化性质[20]、植物群落[21]、土壤微生物群落[22]及土壤呼吸[23]的变化均可以影响小型土壤节肢动物群落。因此, 蚂蚁筑巢行为必然会影响小型土壤节肢动物群落。但目前未见关于蚁丘对小型土壤节肢动物群落多样性影响方面的报道。
青藏高原素有“地球第三极”之称, 是地球上海拔最高、面积最大、形成最晚的高原。高寒草甸是青藏高原上的典型地带性植被, 其生态系统功能较脆弱, 对干扰因子的响应较为强烈[16]。有研究表明, 在青藏高原未退化的高寒草甸中蚁丘密度约为6—9个/hm2, 直径约为30—50 cm[24]。但目前还不清楚高寒草甸蚁丘对小型土壤节肢动物群落多样性的影响。因此, 本文通过调查蚁丘中小型土壤节肢动物, 旨在了解高寒草甸生态系统中蚂蚁筑丘活动对小型土壤节肢动物群落的影响。研究结果不仅可以揭示不同土壤节肢动物类群之间的内在联系, 还可为正确评价蚂蚁在高寒草甸生态系统的功能提供依据。
1 研究区概况若尔盖湿地位于四川省阿坝藏族羌族自治州北部, 地处青藏高原东北边缘, 属于黄河中上游地区。其地理坐标介于102°08′—103°39′E, 32°56′—34°19′N之间。海拔3400—3800 m之间, 平均海拔3500 m。该区属于高原寒带湿润季风气候。最热月为7月, 平均气温10.8℃, 绝对最高气温24.6℃;最冷月为1月, 平均气温-10.6℃, 绝对最低气温-33.7℃。4—7月基本为无霜期, 无绝对无霜期。年降雨量600—800 mm, 其中超过80%集中在4月下旬—10月中旬。年均日照2389 h, 平均日照12 h。年均蒸发量1232 mm。植被以沼泽植被和草甸植被为主;其中草甸植被群落外貌相对单一, 以嵩草属(Kobresia spp.)、蓼属(Polygonum spp.) 植物为主, 禾本科及毒害草伴生。土壤以高原草甸土、高原沼泽土、高原泥炭土为主, 呈中性或微碱性反应, 成土母质多为质地均匀的粉尘和亚粘土。
2 研究方法 2.1 样地设置本研究于2018年5月(春)、7月(夏)、10月(秋), 在若尔盖县唐克乡白河牧场进行。该牧场内分布有大量的广布弓背蚁(Camponotus herculeanus), 喜在落叶和枯枝下营巢。在该牧场内设置一个面积为100 m×100 m的正方形样方作为定位观测样地, 每次在样地内随机选取广布弓背蚁筑巢自由形成的直径大小为30—35 cm, 高度为7—10 cm的蚁丘6个, 蚁丘间隔10 m左右(图 1)。同时在与蚁丘相距2 m处且没有蚁丘扰动的草地选取一个采样点的作为对照。

|
| 图 1 高寒草甸蚁丘 Fig. 1 Ant-hills in alpine meadow ecosystem |
在橡胶手套保护下, 除去蚁丘表层的凋落物等, 使核心部分暴露。用容积为300 cm3环刀(Φ88 mm×h50 mm)快速采集蚁丘内容物, 装入自封袋内。另外, 用300 cm3环刀采集对照土样。采集的样品做相应的标记, 以备室内分离用。3次采样, 共采集土样36份。
在实验室内, 用Tullgren法分离小型土壤节肢动物, 温度控制在38℃左右, 分离时间为48 h。根据《中国土壤动物检索图鉴》[25]、《昆虫分类检索》[26]、《农业螨类学》[27]等文献, 在体式镜(Olympus SZX16)和光学显微镜(Olympus BX53)下对收集到的小型土壤节肢动物进行鉴定。一般鉴定到属, 少数鉴定到科, 并统计个体数量。
2.3 环境因子调查与分析用容积为200 cm3环刀(Φ70 mm×h52 mm)测量蚁丘及对照点的土壤含水量和容重。另取一定量的蚁丘及对照土样用于分析土壤化学性质。
土壤有机质用重铬酸钾氧化—外加热法测定;土壤全氮采用硫酸钾—硫酸铜—硒粉消煮, 定氮仪自动分析法;土壤全磷采用硫酸—高氯酸消煮—钼锑抗比色法;土壤水分含量采用烘干称重法[28]。
2.4 数据分析处理物种优势度的划分:个体数量占总捕获量10%以上的类群为优势类群;1%—10%左右的为常见类群;1%以下的为稀有类群。
群落多样性的计算:采用以下几个参数来分析小型土壤节肢动物群落多样性。
丰富度指数:以小型土壤节肢动物的类群数表示。
Shannon多样性指数(H):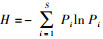
Pielou优势度指数(E):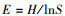
式中, S代表总类群数, Pi为每个生境中第i个类群的个体密度占该生境小型土壤节肢动物总数的比例。
利用主成分分析法(Principal Component Analysis, PCA), 对小型土壤节肢动物群落结构组成进行排序, 分析过程中对其个体密度进行log(x+1)转换。
利用双因素方差分析对蚁丘、对照和月份间的小型土壤节肢动物群落的密度及多样性进行检验。另外, 用单因素方差分析分别对蚁丘和对照的不同月份间的差异性进行检验。分析前对数据进行正态检验, 如果不符合正态分布则进行log(x+1)。对转换后仍不符合正态分布的数据改用非参数检验。获得显著性后, 采用LSD法或Mann — Whitney U test法进行多重比较。采用回归分析小型土壤节肢动物的个体密度、多样性指数等与环境因子之间的关系。
数据分析采用IBM SPSS 20.0和Canoco for windows 4.5软件进行完成。
3 结果与分析 3.1 小型土壤节肢动物群落组成结构差异蚁丘中小型土壤节肢动物群落的组成结构与对照有一定差异(表 1)。蚁丘和对照的小型土壤节肢动物类群数分别为34类、20类, 呈下降趋势。不同生境中, 各类群的个体数量在群落中所占的比例也存在变化。在属层次上, 蚁丘中的优势类群为库跳属(Coloburella)和隐跳属(Cryptopygus), 个体数分别占捕获量的27.29%和20.92%;常见类群包含双瘤吸螨属(Biscirus)和布伦螨属(Brennandania)等14类, 占总捕获量的46.94%;稀有类群包含露尾甲科成虫(Nitidulidae adult)和尖奥甲螨属(Oxyoppia)等18类, 占总捕获量的4.85%。对照中的优势类群为拟缺跳属(Pseudanurophorus)、布伦螨属(Brennandania)、盾螨科(Scutacaridae)和绒螨科( Trombidiidae), 个体数分别占捕获量的13.86%、18.81%、10.89%和14.85%;常见类群包含双瘤吸螨属(Biscirus)和植绥螨科(Phytoseiidae)等9类, 占总捕获量的34.65%;稀有类群包含小大翼甲螨属(Galumnella)和象甲科成虫(Curculionidae adult)等7类, 占总捕获量的6.93%。
| 类群 Groups |
蚁丘Ant-hills | 对照Controls | 总计Total | |||||
| 个体数 Individuals |
% | 个体数 Individuals |
% | 个体数 Individuals |
% | |||
| 弹尾纲Collembola | 1484 | 54.36 | 25 | 24.75 | 1509 | 53.30 | ||
| 长角跳目Entomobryomorph | 1454 | 53.26 | 19 | 18.81 | 1473 | 52.03 | ||
| 库跳属 Coloburella | 745 | 27.29 | 3 | 2.97 | 748 | 26.42 | ||
| 隐跳属Cryptopygus | 571 | 20.92 | 2 | 1.98 | 573 | 20.24 | ||
| 符跳属Folsomia | 112 | 4.10 | — | — | 112 | 3.96 | ||
| 拟缺跳属Pseudanurophorus | 12 | 0.44 | 14 | 13.86 | 26 | 0.92 | ||
| 长跳属Entomobrya | 8 | 0.29 | — | — | 8 | 0.28 | ||
| 鳞跳属Tomocerus | 4 | 0.15 | — | — | 4 | 0.14 | ||
| 近缺跳属Paranurophorus | 1 | 0.04 | — | — | 1 | 0.04 | ||
| 驼跳属Cyphoderus | 1 | 0.04 | — | — | 1 | 0.04 | ||
| 原跳目Poduromorpha | 29 | 1.06 | 4 | 3.96 | 33 | 1.17 | ||
| 奇跳属Xenylla | 11 | 0.40 | 4 | 3.96 | 15 | 0.53 | ||
| 疣跳属Neanura | 12 | 0.44 | — | — | 12 | 0.42 | ||
| 跳虫属Podura | 4 | 0.15 | — | — | 4 | 0.14 | ||
| 副跳属 Paranura | 2 | 0.07 | — | — | 2 | 0.07 | ||
| 愈腹跳目Symphyleona | 1 | 0.04 | 2 | 1.98 | 3 | 0.11 | ||
| 钩圆跳属Bouretiella | 1 | 0.04 | 2 | 1.98 | 3 | 0.11 | ||
| 蛛形纲Arachnida | 1203 | 44.07 | 69 | 68.32 | 1272 | 44.93 | ||
| 前气门亚目Prostigmata | 694 | 25.42 | 55 | 54.46 | 749 | 26.46 | ||
| 布伦螨属Brennandania | 233 | 8.53 | 19 | 18.81 | 252 | 8.90 | ||
| 双瘤吸螨属Biscirus | 234 | 8.57 | 9 | 8.91 | 243 | 8.58 | ||
| 盾螨科Scutacaridae | 65 | 2.38 | 11 | 10.89 | 76 | 2.68 | ||
| 矮蒲螨属Pygmephorus | 66 | 2.42 | 1 | 0.99 | 67 | 2.37 | ||
| 绒螨科Trombidiidae | 35 | 1.28 | 15 | 14.85 | 50 | 1.77 | ||
| 小黑螨属Caligonellidae | 49 | 1.79 | — | — | 49 | 1.73 | ||
| 肉食螨科Cheylrtidae | 9 | 0.33 | — | — | 9 | 0.32 | ||
| 大赤螨科Anystidae | 3 | 0.11 | — | — | 3 | 0.11 | ||
| 甲螨亚目Oribatida | 381 | 13.96 | 3 | 2.97 | 384 | 13.56 | ||
| 微奥甲螨属Microppia | 126 | 4.62 | — | — | 126 | 4.45 | ||
| 下盾甲螨属Hypovertex | 103 | 3.77 | 2 | 1.98 | 105 | 3.71 | ||
| 小大翼甲螨属Galumnella | 83 | 3.04 | 1 | 0.99 | 84 | 2.97 | ||
| 弓奥甲螨属Arcoppia | 48 | 1.76 | — | — | 48 | 1.70 | ||
| 尖奥甲螨属 Oxyoppia | 21 | 0.77 | — | — | 21 | 0.74 | ||
| 中气门亚目Mesostigmata | 128 | 4.69 | 11 | 10.89 | 139 | 4.91 | ||
| 寄螨科Parasitidae | 52 | 1.90 | 4 | 3.96 | 56 | 1.98 | ||
| 植绥螨科Phytoseiidae | 46 | 1.68 | 7 | 6.93 | 53 | 1.87 | ||
| 肛厉螨属Proctolaelaps | 30 | 1.10 | — | — | 30 | 1.06 | ||
| 昆虫纲Insecta | 39 | 1.43 | 7 | 6.93 | 46 | 1.62 | ||
| 鞘翅目Coleoptera | 35 | 1.28 | 2 | 1.98 | 37 | 1.31 | ||
| 露尾甲科成虫Nitidulidae adult | 25 | 0.92 | 1 | 0.99 | 26 | 0.92 | ||
| 露尾甲科幼虫Nitidulidae larvae | 8 | 0.29 | — | — | 8 | 0.28 | ||
| 蚁形甲科成虫Anthicidae adult | 2 | 0.07 | — | — | 2 | 0.07 | ||
| 象甲科成虫Curculionidae adult | — | — | 1 | 0.99 | 1 | 0.04 | ||
| 双翅目幼虫Diptera larvae | 4 | 0.15 | 5 | 4.95 | 9 | 0.32 | ||
| 瘦足蝇科Micropezidae | 4 | 0.15 | — | — | 4 | 0.14 | ||
| 蝇科Muscidae | — | — | 2 | 1.98 | 2 | 0.07 | ||
| 尖眼蕈蚊科Sciaridae | — | — | 1 | 0.99 | 1 | 0.04 | ||
| 扁角蚊科Ceroplatidae | — | — | 1 | 0.99 | 1 | 0.04 | ||
| 摇蚊科Chironomidae | — | — | 1 | 0.99 | 1 | 0.04 | ||
| 双尾纲Diplura | 4 | 0.15 | — | — | 4 | 0.14 | ||
| 双尾目Diplura | 4 | 0.15 | — | — | 4 | 0.14 | ||
| 鳞虫八属Lepidocampa | 4 | 0.15 | — | — | 4 | 0.14 | ||
| 总个体数Total individuals | 2730 | 100.00 | 101 | 100.00 | 2831 | 100.00 | ||
在纲层次上, 弹尾纲(Collembola)个体数在蚁丘中占总捕获量的54.36%, 在对照中占24.75%。蛛形纲(Arachnida)个体数在蚁丘中占总捕获量的44.07%, 在对照中占68.32%。昆虫纲(Insecta)个体数在蚁丘中占总捕获量1.43%, 在对照中占6.93%。双尾纲(Diplura)个体数在蚁丘中占0.15%, 在对照中没有发现。
小型土壤节肢动物群落排序分析结果(图 2)表明, 蚁丘和对照间的小型土壤节肢动物群落组成结构存在明显差异, 且随采样月份变化, 以5、10月的小型土壤节肢动物群落结构差异较明显。此外, 不同月份间, 影响小型土壤节肢动物群落结构的主要类群不同。5、7、10月份, 在PC1轴上影响群落结构的主要类群分别是绒螨科(Trombidiidae)、库跳属(Coloburella)和小黑螨属(Caligonellidae)等, 在PC2轴上影响群落结构的主要类群是隐跳属(Cryptopygus)、下盾甲螨属(Hypovertex)和寄螨科(Parasitidae)等。

|
| 图 2 蚁丘与对照间小型土壤节肢动物群落结构差异 Fig. 2 Differences in soil microarthropod communities between ant-hills and controls |
长角跳目、前气门亚目、甲螨亚目在群落中所占比例在10%以上, 是优势类群(表 1), 因此对这3个类群的变化进行分析。从3个月的整体情况来看, 蚁丘中长角跳目和甲螨亚目的个体密度及个体百分比均高于对照;而前气门亚目的个体密度高于对照, 个体百分比则低于对照(图 3)。双因素方差分析(表 2)结果表明, 蚁丘中3个优势类群的个体密度均显著高于对照(P<0.05), 但蚁丘中仅甲螨亚目个体百分比显著高于对照(P<0.05), 而长角跳目及前气门亚目个体百分比在蚁丘和对照间无显著差异(P>0.05)。

|
| 图 3 优势类群的个体密度及百分比动态(平均值±标准误) Fig. 3 The dynamics in individual density and percentage of the dominant groups (Mean±SE) 不同大写字母表示不同生境间差异显著(P<0.05), 不同小写字母表示相同生境不同月份间差异显著(P<0.05) |
| 来源 Sources |
df | 长角跳目Entomobrvomorph | 前气门亚目Prostigmata | 甲螨亚目Oribatida | |||||||||||
| 密度Density | % | 密度Density | % | 密度Density | % | ||||||||||
| F | P | F | P | F | P | F | P | F | P | F | P | ||||
| 蚁丘Ant-hills | 1, 10 | 4.66 | 0.039 | 1.78 | 0.192 | 10.68 | 0.003 | 4.15 | 0.051 | 7.72 | 0.009 | 8.75 | 0.006 | ||
| 月份Month | 2, 10 | 0.83 | 0.445 | 0.37 | 0.693 | 3.09 | 0.060 | 0.98 | 0.387 | 2.77 | 0.079 | 1.07 | 0.357 | ||
| 蚁丘Ant-hills ×月份Month | 2, 20 | 0.79 | 0.465 | 0.88 | 0.426 | 2.85 | 0.074 | 2.90 | 0.071 | 2.69 | 0.084 | 0.67 | 0.521 | ||
蚁丘中小型土壤节肢动物群落的类群数、个体密度、Shannon多样性指数均高于对照;而Pielou均匀度指数无明显变化(图 4)。月份变化上, 从5月到10月, 蚁丘和对照的小型土壤节肢动物群落个体密度、Shannon多样性指数、类群数均呈增加趋势(图 4)。双因素方差分析结果表明(表 3), 蚁丘中小型土壤节肢动物群落的个体密度、类群数和Shannon多样性指数均显著高于对照(P<0.05);月份变化对、类群数、Shannon多样性指数也有显著影响(P<0.05)。蚁丘中小型土壤节肢动物群落仅有个体密度在不同月份间存在显著差异(P<0.05), 而对照中小型土壤节肢动物群落类群数、Shannon多样性指数、Pielou均匀度指数均随月份变化显著增加(P<0.05)。

|
| 图 4 蚁丘及对照的小型土壤节肢动物群落密度及多样性(平均值±标准误) Fig. 4 The individual density and diversity index of soil microarthropod communities in ant-hills and controls (Mean±SE) 不同大写字母表示不同生境间差异显著(P<0.05), 不同小写字母表示同一生境不同月份间差异显著(P<0.05) |
| 来源 Sources |
df | 个体密度 Individual density |
类群数 Group number |
多样性指数(H) Shannon index |
均匀度指数(E) Pielou index |
|||||||
| F | P | F | P | F | P | F | P | |||||
| 蚁丘Ant-hills | 1, 10 | 10.38 | 0.003 | 23.65 | <0.001 | 4.65 | 0.039 | 0.001 | 0.973 | |||
| 月份Month | 2, 10 | 1.78 | 0.187 | 5.38 | 0.010 | 4.45 | 0.020 | 3.19 | 0.056 | |||
| 蚁丘Ant-hills×月份Month | 2, 20 | 1.65 | 0.210 | 0.53 | 0.592 | 0.28 | 0.752 | 2.38 | 0.109 | |||
蚁丘和对照两个不同生境的土壤理化性质如表 4。蚁丘土壤中的有机质、全氮、全磷含量均显著高于对照土壤(P<0.01或0.05), 但容重、含水量均显著低于对照区土壤(P<0.01)。
| 环境变量Environmental variables | 蚁丘Ant-hills | 对照Controls | t | P |
| 含水量Water content /% | 0.07±0.01 | 0.35±0.02 | 12.51 | <0.001 |
| 容重Bulk density /(g/cm3) | 0.23±0.02 | 0.91±0.06 | 9.22 | <0.001 |
| 有机质Soil organic matter /(g/kg) | 221.02±29.02 | 100.81±9.66 | 4.86 | 0.002 |
| 全氮Total nitrogen /(g/kg) | 8.35±0.56 | 4.66±0.43 | 4.28 | 0.002 |
| 全磷Total phosphorus /(g/kg) | 1.46±0.08 | 0.87±0.05 | 7.49 | 0.001 |
小型土壤节肢动物群落各参数与环境因子的回归分析(表 5)结果表明, 小型土壤节肢动物的类群数、Shannon多样性指数、前气门亚目的密度与有机质、全氮、全磷含量呈显著正相关(P<0.05), 个体密度与有机质含量呈显著正相关(P<0.05), 甲螨亚目的密度与全氮含量呈显著正相关(P<0.05), 个体密度、类群数、Shannon多样性指数、长角跳目、前气门亚目、甲螨亚目的密度与土壤容重呈显著负相关(P<0.05)。
| 环境变量 Environmental variables |
个体密度 Individual density |
类群数 Group number |
多样性指数(H) Shannon index |
均匀度指数(E) Pielou index |
长角跳目 Entomobryomorph |
前气门亚目 Prostigmata |
甲螨亚目 Oribatida |
| 含水量/% Water content |
-0.45 | -0.48 | -0.34 | 0.06 | -0.41 | -0.43 | 0.43 |
| 容重/(g/cm3) Bulk density |
-0.62* | -0.89** | -0.65* | 0.07 | -0.49** | -0.79** | 0.75** |
| 有机质/(g/kg) Soil organic matter |
0.60* | 0.95** | 0.76** | 0.05 | 0.49 | 0.83** | 0.58** |
| 全氮/(g/kg) Total nitrogen |
0.56 | 0.91** | 0.72** | 0.05 | 0.46 | 0.77** | 0.62* |
| 全磷/(g/kg) Total phosphorus |
0.43 | 0.83** | 0.67* | 0.05 | 0.34 | 0.64* | 0.46 |
| *表示显著性水平P<0.05;**表示P<0.01, 双尾检验 | |||||||
本研究结果表明, 蚁丘中小型土壤节肢动物群落结构与对照即高寒草甸土壤明显不同, 以螨类为主要类群的小型土壤节肢动物群落转变为以跳虫为主要类群, 说明蚁丘对小型土壤节肢动物的群落组成影响较大。此外, 蚁丘还能够显著提高小型土壤节肢动物群落的个体密度和多样性, 反映出蚂蚁通过筑丘活动产生与周围不同的微环境, 有利于小型土壤节肢动物生存。
首先, 吴鹏飞等在对高寒草甸生态系统土壤动物群落的研究中发现食物资源的多寡和环境条件改变是影响小型土壤节肢动物群落结构及多样性的主要原因[29]。本研究中, 蚁丘小型土壤节肢动物密度显著高于对照主要是由于弹尾纲中的库跳属、隐跳属和符跳属以及螨类中的布伦螨属、双瘤吸螨属、微奥甲螨属和下盾甲螨属的个体数量显著增加。库跳属、隐跳属和符跳属主要为菌食性[30], 而布伦螨属、双瘤吸螨属、微奥甲螨属和下盾甲螨属主要为腐食性[31]。由此可知, 蚁丘环境有利于菌食性跳虫和腐食性螨类。这可能是因为蚁丘内的环境有利于促进土壤微生物生长和繁殖[18, 32], 能够为菌食性小型土壤节肢动物提供丰富的食物资源[31]。而蚂蚁通过筑巢、取食及排泄等活动可把其他昆虫尸体、植物组织、动物粪便以及自身的排泄物聚集在蚁丘内[33-34]。我们通过对蚁丘土样过筛处理, 发现蚁丘土壤主要是牦牛粪便、鼠兔粪便、细碎粪便与土壤草屑的混合物, 平均比例为1%、53%和46%。大量的脊椎动物粪便不仅可增加土壤有机质、改善土壤结构, 促进微生物生长, 还可为腐食性小型土壤节肢动物提供大量的食物资源[35]。马金豪等在若尔盖沙化草地生态恢复过程中, 也发现向土壤中添加家畜粪便等有机物能影响土壤动物群落结构及多样性[36]。
其次, 土壤理化性质是小型土壤节肢动物生存的条件, 能引起类群数、个体密度的分布差异[37]。本研究发现土壤容重与个体密度、类群数、多样性指数呈显著负相关, 这与崔丽巍等人在川西北冷杉林群落恢复过程中土壤动物群落的研究结果一致[38]。有机质含量与类群数、多样性指数、个体密度显著正相关, 全氮、全磷含量与类群数、多样性指数呈显著正相关, 而吴鹏飞等人在研究土壤动物对高寒草甸退化响应的过程中也有类似的发现[29]。蚁丘内的土壤容重显著低于对照, 这可能与蚁丘土壤主要由草屑、动物粪便组成有关;使土壤处于疏松状态, 通透性好, 有利于小型土壤节肢动物的活动和繁殖[39-40]。而有机质、全氮、全磷含量则显著高于对照, 这种富集作用主要是由于蚂蚁的粪便、排泄物、动物尸体、植物残渣等在蚁巢内聚集[33-34];从而显著改善了土壤结构, 提高土壤养分, 可为小型土壤节肢动物提供丰富的食物资源和更为合适的居住环境[41-42]。本研究中含水量虽然与小型土壤节肢动物的个体密度、类群数、多样性指数、均匀度指数的相关性不显著, 但其变化趋势与小型土壤节肢动物群落多样性动态基本一致, 可能的原因是, 食物等资源充足的情况下小型土壤节肢动物群落内不同个体间的竞争排斥作用较弱[43]。
此外, 李霁航等在研究蚂蚁筑巢对土壤呼吸影响的过程中发现蚁丘内能够维持较高的土壤温度[19]。这可能与蚁巢对温度的物理隔离、蚁巢内蚂蚁数量众多, 蚂蚁呼吸以及蚁巢内微生物的新陈代谢产生较多的热量密切相关[32, 44]。李萌等在贡嘎山东坡典型植被类型土壤动物群落特征的研究中发现, 土壤动物群落的个体密度、多样性指数与土壤温度呈正相关[45]。蒙凤群等在川西北高寒草甸蚁丘对植物群落演替的研究中表明蚂蚁筑巢降低了植物群落密度, 增加了光照强度[33];而太阳辐射量的增加可以提高小型土壤节肢动物个体密度和多样性[46]。因此, 蚁丘中小型土壤节肢动物的个体密度、多样性指数均高于对照。蚂蚁作为地球上分布广泛、种类丰富和数量巨大的社会性昆虫[18], 对生态系统结构和功能的形成有关键的作用[47]。目前已有将蚂蚁用作土地利用与保护[48]、植被恢复[49]、牧场监测[50]、生物多样性[51]等方面的指示生物。鉴于本文发现蚁穴中小型土壤节肢动物密度和多样性显著高于对照, 因此蚂蚁可以作为高寒草甸生态系统小型土壤节肢动物多样性的指示生物。
4.2 蚁丘对小型土壤节肢动物季节动态的影响本研究还发现蚁丘和对照的小型土壤节肢动物季节动态不同, 在蚁丘中小型土壤节肢动物仅个体密度有显著的季节变化, 而对照中的类群数、多样性指数、均匀度指数有显著的季节变化。蚁丘内小型土壤节肢动物密度显著变化主要是因为7月份库跳属、下盾甲螨属的个体数量和10月隐跳属、双瘤吸螨属的个体数量都远大于5月, 而这几类主要为菌食性和腐食性小型土壤节肢动物[30, 52]。在高寒草地, 5—10月份的月均气温8.0 ℃, 月均土温12.0 ℃[53], 5月表层土壤刚解冻, 气温较低, 不利微生物繁殖和腐殖质形成, 而7、10月温湿度相对较高, 有利于蚁穴内微生物繁殖和腐殖质的形成。此外, 在若尔盖地区, 年降雨量600—800 mm, 且主要集中在5—8月[54], 地表水分聚集不利于螨类和跳虫土壤动物的取食和栖息[55], 因此菌食性和腐食性小型土壤节肢动物的个体密度显著增加。作为对照的高寒草甸中小型土壤节肢动物多样性指数季节变化显著是因为小型土壤节肢动物群落的类群数和Shannon多样性指数在5、10月间有显著差异。其原因可能是因为这期间若尔盖高寒草地植物群落的组成种类、高度、盖度以及气候因子的季节变化相对较大, 影响了小型土壤节肢动物的群落组成和多样性。吴鹏飞等在大型土壤动物群落对高寒草甸退化的响应的研究中发现, 降水的季节性增加可导致部分小型土壤节肢动物的多样性下降[29]。, 而蚁丘则因为其植物少, 且有较多的草碎屑等物质, 可以遮挡雨水进入, 因此蚁丘内部的环境相对稳定, 小型土壤节肢动物群落的类群组成结构也相对稳定, 导致多样性指数的季节变化不显著。
5 结论高寒草甸生态系统的蚁丘能明显改变小型土壤节肢动物群落组成结构, 使以螨类为主要类群的高寒草地小型土壤节肢动物群落转变为以跳虫为主要类群。蚁丘还能显著提高小型土壤节肢动物的群落密度及多样性, 尤其是能够显著增加菌食性跳虫和腐食性螨类的个体数量。此外, 蚁丘对小型土壤节肢动物群落组成及多样性的季节动态也有明显的影响。土壤容重和有机质含量分别是影响小型土壤节肢动物群落密度和多样性的主要环境因子。
| [1] |
Wardle D A, Bardgett R D, Klironomos J N, Setälä H, van der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629-1633. DOI:10.1126/science.1094875 |
| [2] |
Liu R T, Zhao H L, Zhao X Y. Effect of vegetation restoration on ant nest-building activities following mobile dune stabilization in the Horqin Sandy Land, Northern China. Land Degradation & Development, 2009, 20(5): 562-571. |
| [3] |
Chen H L, Li B, Fang C M, Chen J K, Wu J H. Exotic plant influences soil nematode communities through litter input. Soil Biology and Biochemistry, 2007, 39(7): 1782-1793. DOI:10.1016/j.soilbio.2007.02.011 |
| [4] |
Goralczyk K. Nematodes in a coastal dune succession: indicators of soil properties?. Applied Soil Ecology, 1998, 9(1/3): 465-469. |
| [5] |
武崎, 吴鹏飞, 王群, 文勇立, 高艳美, 张荣芝, 龙伟. 放牧强度对高寒草地不同类群土壤动物的群落结构和多样性的影响. 中国农业科学, 2016, 49(9): 1826-1834. |
| [6] |
叶国辉, 楚彬, 周睿, 张飞宇, 华铣泽, 花蕊, 王婷, 花立民. 高原鼠兔干扰对高寒草甸大型土壤动物类群的影响. 甘肃农业大学学报, 2019, 54(4): 150-158. |
| [7] |
Knoepp J D, Coleman D C, Crossley Jr D A, Clark J S. Biological indices of soil quality: an ecosystem case study of their use. Forest Ecology and Management, 2000, 138(1/3): 357-368. |
| [8] |
Folgarait P J. Ant biodiversity and its relationship to ecosystem functioning: a review. Biodiversity & Conservation, 1998, 7(9): 1221-1244. |
| [9] |
Dauber J, Niechoj R, Baltruschat H, Wolters V. Soil engineering ants increase grass root arbuscular mycorrhizal colonization. Biology and Fertility of Soils, 2008, 44(5): 791-796. DOI:10.1007/s00374-008-0283-5 |
| [10] |
赵有益, 龙瑞军, 林慧龙, 任继周. 草地生态系统安全及其评价研究. 草业学报, 2008, 17(2): 143-150. |
| [11] |
King T J. The plant ecology of ant-hills in calcareous grasslands: I. Patterns of species in relation to ant-hills in southern England. Journal of Ecology, 1977, 65(1): 235-256. DOI:10.2307/2259077 |
| [12] |
刘任涛, 赵哈林, 赵学勇. 沙地生态系统中蚂蚁活动与地表植被及土壤环境间的互作关系. 干旱区资源与环境, 2011, 25(12): 166-170. |
| [13] |
Verchot L V, Moutinho P R, Davidson E A. Leaf-cutting ant (Atta sexdens) and nutrient cycling: deep soil inorganic nitrogen stocks, mineralization, and nitrification in Eastern Amazonia. Soil Biology and Biochemistry, 2003, 35(9): 1219-1222. DOI:10.1016/S0038-0717(03)00183-4 |
| [14] |
侯继华, 周道玮, 姜世成. 蚂蚁筑丘活动对松嫩草地植物群落多样性的影响. 植物生态学报, 2002, 26(3): 323-329. |
| [15] |
向泽宇, 张莉, 刘雪娟. 蚁丘干扰下高寒草甸植物群落特征和生态位. 生态学杂志, 2016, 35(2): 321-329. |
| [16] |
陈骥, 曹军骥, 张思毅, 樊云龙, 王小菲, 魏永林. 蚂蚁扰动对青海湖北岸高寒草甸草原群落结构影响. 地球环境学报, 2013, 4(5): 1461-1469, 1480-1480. |
| [17] |
杨有芳, 字洪标, 刘敏, 阿的鲁骥, 陈焱, 王长庭. 高寒草甸土壤微生物群落功能多样性对广布弓背蚁蚁丘扰动的响应. 草业学报, 2017, 26(1): 43-53. |
| [18] |
Boulton A M, Amberman K D. How ant nests increase soil biota richness and abundance: a field experiment. Biodiversity & Conservation, 2006, 15(1): 69-82. |
| [19] |
李霁航, 王邵军, 王红, 张哲, 曹润, 陈闽昆, 李少辉, 陈奇伯. 蚂蚁筑巢对高檐蒲桃热带森林群落土壤呼吸的影响. 生态学报, 2018, 38(17): 6033-6042. |
| [20] |
Wu P F, Zhang H Z, Wang Y. The response of soil macroinvertebrates to alpine meadow degradation in the Qinghai-Tibetan Plateau, China. Applied Soil Ecology, 2015, 90: 60-67. DOI:10.1016/j.apsoil.2015.02.006 |
| [21] |
周育臻, 吴鹏飞. 贡嘎山东坡森林小型土壤节肢动物群落多样性与时空分布. 生态学杂志, 2020, 39(2): 586-599. |
| [22] |
李俊, 吴福忠, 杨万勤, 谭波, 彭艳, 王滨, 常晨晖. 高山草甸冬季凋落物分解过程中土壤动物对微生物群落结构的影响. 应用与环境生物学报, 2016, 22(1): 27-34. |
| [23] |
王可洪, 武海涛, 吴东辉. 土壤动物对土壤呼吸的影响研究进展. 土壤通报, 2013, 44(6): 1526-1530. |
| [24] |
Wang C T, Wang G X, Wu P F, Rafique R, Zi H B, Li X Z, Luo Y Q. Effects of ant mounds on the plant and soil microbial community in an alpine meadow of Qinghai-Tibet Plateau. Land Degradation & Development, 2017, 28(5): 1538-1548. |
| [25] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [26] |
李鸿兴, 隋敬之, 周士秀, 周勤, 孙洪国. 昆虫分类检索. 北京: 农业出版社, 1987.
|
| [27] |
忻介六. 农业螨类学. 北京: 农业出版社, 1988.
|
| [28] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
|
| [29] |
吴鹏飞, 张洪芝, 崔丽巍, 钟红梅, 王永. 大型土壤动物群落对高寒草甸退化的响应. 土壤学报, 2013, 50(4): 786-799. |
| [30] |
张阿娟, 张健, 李金金, 刘志刚, 张丹桔. 轮伐期前后不同密度巨桉(Eucalyptus grandis)人工林土壤动物群落结构特征. 生态学报, 2020, 40(3): 808-821. |
| [31] |
周泓杨, 张健, 张丹桔, 张捷, 魏大平, 赵燕波, 赵波, 李川北. 不同郁闭度控制下马尾松(Pinus massoniana)人工林土壤动物群落特征. 生态学报, 2017, 37(6): 1939-1955. |
| [32] |
曹乾斌, 王邵军, 任玉连, 张哲, 陈闽昆, 李少辉, 曹润, 王平, 左倩倩. 蚂蚁筑巢对西双版纳热带森林土壤碳矿化动态的影响. 应用生态学报, 2019, 30(12): 4231-4239. |
| [33] |
蒙凤群, 高贤明, 孙书存. 川西北高寒草甸蚁丘植物群落演替: 种类组成与物种多样性. 植物分类与资源学报, 2011, 33(2): 191-199. |
| [34] |
陈应武, 李新荣, 苏延桂, 窦彩虹, 贾晓红, 张志山. 腾格里沙漠人工植被区掘穴蚁(Formica cunicularia)的生态功能. 生态学报, 2007, 27(4): 1508-1514. |
| [35] |
Scrimgeour G J, Kendall S. Effects of livestock grazing on benthic invertebrates from a native grassland ecosystem. Freshwater Biology, 2003, 48(2): 347-362. |
| [36] |
马金豪, 吴鹏飞. 土壤动物群落对沙化高寒草地生态恢复的响应. 生态学杂志, 2018, 37(12): 3566-3575. |
| [37] |
李雨, 吴鹏飞, 龙伟, 马金豪. 高寒地区种植不同种类牧草对土壤节肢动物群落的影响. 生态学报, 2019, 39(20): 7697-7708. |
| [38] |
崔丽巍, 刘世荣, 刘兴良, 吴鹏飞, 张洪芝, 何先进. 川西北冷杉林恢复过程中土壤动物群落动态. 生态学报, 2012, 32(15): 4772-4783. |
| [39] |
赵乌英嘎, 红梅, 赵巴音那木拉, 刘鹏飞, 美丽, 王文东, 李金龙. 不同耕作方式下黑土区农田中小型土壤动物群落特征. 水土保持通报, 2019, 39(3): 39-45. |
| [40] |
Ge G F, Li Z J, Zhang J, Wang L G, Xu M G, Zhang J B, Wang J K, Xie X L, Liang Y C. Geographical and climatic differences in long-term effect of organic and inorganic amendments on soil enzymatic activities and respiration in field experimental stations of China. Ecological Complexity, 2009, 6(4): 421-431. |
| [41] |
Hu C, Qi Y. Abundance and diversity of soil nematodes as influenced by different types of organic manure. Helminthologia, 2010, 47(1): 58-66. DOI:10.2478/s11687-010-0009-8 |
| [42] |
Cole L, Buckland S M, Bardgett R D. Relating microarthropod community structure and diversity to soil fertility manipulations in temperate grassland. Soil Biology and Biochemistry, 2005, 37(9): 1707-1717. |
| [43] |
Cole L, Buckland S M, Bardgett R D. Influence of disturbance and nitrogen addition on plant and soil animal diversity in grassland. Soil Biology and Biochemistry, 2007, 40(2): 505-514. |
| [44] |
Frouz J. The effect of nest moisture on daily temperature regime in the nests of Formica polyctena wood ants. Insectes Sociaux, 2000, 47(3): 229-235. DOI:10.1007/PL00001708 |
| [45] |
李萌, 吴鹏飞, 王永. 贡嘎山东坡典型植被类型土壤动物群落特征. 生态学报, 2015, 35(7): 2295-2307. |
| [46] |
Salmon S, Artuso N, Frizzera L, Zampedri R. Relationships between soil fauna communities and humus forms: response to forest dynamics and solar radiation. Soil Biology and Biochemistry, 2008, 40(7): 1707-1715. |
| [47] |
Werling B P, Dickson T L, Isaacs R, Gaines H, Gratton C, Gross K L, Liere H, Malmstrom C M, Meehan T D, Ruan L, Robertson B A, Robertson G P, Schmidt T M, Schrotenboer A C, Teal T K, Wilson J K, Landis D A. Perennial grasslands enhance biodiversity and multiple ecosystem services in bioenergy landscapes. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(4): 1652-1657. |
| [48] |
Majer J D. Ants: Bio-indicators of minesite rehabilitation, land-use, and land conservation. Environmental Management, 1983, 7(4): 375-383. |
| [49] |
李巧, 卢志兴, 张威, 马艳滟, 冯萍. 金沙江干热河谷人工林地表的蚂蚁群落. 林业科学, 2015, 51(8): 134-141. |
| [50] |
Andersen A N, Fisher A, Hoffmann B D, Read J L, Richards R. Use of terrestrial invertebrates for biodiversity monitoring in Australian rangelands, with particular reference to ants. Austral Ecology, 2004, 29(1): 87-92. |
| [51] |
Hoffmann B D. Using ants for rangeland monitoring: global patterns in the responses of ant communities to grazing. Ecological Indicators, 2010, 10(2): 105-111. |
| [52] |
黄玉梅, 杨万勤, 张健, 卢昌泰, 刘旭, 王伟, 郭伟. 川西亚高山针叶林土壤动物群落对模拟林下植物丧失的响应. 生态学报, 2010, 30(8): 2018-2025. |
| [53] |
张洪芝, 吴鹏飞, 杨大星, 崔丽巍, 何先进, 熊远清. 青藏东缘若尔盖高寒草甸中小型土壤动物群落特征及季节变化. 生态学报, 2011, 31(15): 4385-4397. |
| [54] |
龙伟, 高艳美, 吴鹏飞. 若尔盖高寒草甸退化对表栖节肢动物群落的影响. 生态学杂志, 2018, 37(1): 128-138. |
| [55] |
Staley J T, Hodgson C J, Mortimer S R, Morecroft M D, Masters G J, Brown V K, Taylor M E. Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates. European Journal of Soil Biology, 2007, 43(3): 189-198. |