 2021, Vol. 41
2021, Vol. 41文章信息
- 邓晨晖, 白红英, 马新萍, 黄晓月, 赵婷
- DENG Chenhui, BAI Hongying, MA Xinping, HUANG Xiaoyue, ZHAO Ting
- 2000-2017年秦岭山地植被物候变化特征及其南北差异
- Variation characteristics and its north-south differences of the vegetation phenology by remote sensing monitoring in the Qinling Mountains during 2000-2017
- 生态学报. 2021, 41(3): 1068-1080
- Acta Ecologica Sinica. 2021, 41(3): 1068-1080
- http://dx.doi.org/10.5846/stxb202002140250
-
文章历史
- 收稿日期: 2020-02-14
- 网络出版日期: 2020-12-14

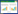




2. 陕西师范大学地理科学与旅游学院, 西安 710119;
3. 西北大学城市与资源环境学院, 西安 710127
2. College of Geography and Tourism, Shaanxi Normal University, Xi'an 710119, China;
3. College of Urban and Environmental Science, Northwest University, Xi'an 710127, China
气候变化对陆地生态系统产生了重要影响, 已成为国内外学者关注的重大科学问题。气候变化表现出显著的区域差异性, 生态脆弱带、生态敏感区, 尤其是山地及中高纬地区的地理过渡带等区域被认为是全球变暖信号的放大器[1-5], 山地过渡带是响应气候变化的典型区域。山地生态系统重要组成部分的植被被认为是响应气候变化的敏感指示器, 植被物候对气候波动非常敏感, 其变化不仅可以揭示山地植被的生长动态, 而且能够直观反映山地生态系统对全球环境变化的响应与适应过程[6-9], 被认为是表征植被-气候相互关系变化的重要指标之一, 以及生物圈对气候变暖反应最敏感也最明显的指示[10-14], 已成为目前全球气候评价与自然环境变化的重要指标[15-16]。因此, 研究气候变化背景下山地过渡带植被物候的变化规律, 探讨山地过渡带地理环境变化的规律和区域差异, 对于深入了解植被—气候关系及揭示山地过渡带对气候变化的响应方式具有重要意义。
植物物候监测主要通过一定的方法和手段对植物生长节律变化进行监测[17], 传统的地面物候观测是从个体水平对物种进行监测;在大尺度物候变化时空格局研究中, 亦有学者采用对物候影响占主导地位的温度数据进行提取气候生长期开展研究[18-22];近年来, 遥感技术为物候研究提供了新的手段, 遥感物候则是通过卫星在群落和生态系统水平上监测地表植被指数, 该物候期是在像元尺度上, 单个像元涵盖很多物种, 空间覆盖广, 连续性强, 便于区域乃至全球尺度物候变化研究, 成为传统物候观测方法的有效补充[23]。因此, 遥感监测的植被指数近年来受到众多物候研究者的青睐[24-27], 其中归一化植被指数(NDVI)被广泛用于反演植被物候期[28-32]。然而, 有学者指出NDVI存在某些缺陷, 尤其是在高植被覆盖区存在大气噪声、土壤背景、饱和度等问题[33]。亦有学者发现, 在解析不同土壤背景下的植被信息时增强型植被指数2(EVI2)要优于NDVI[34], EVI2不仅改进了NDVI的缺陷, 同时在高生物量地区具有更高的敏感性, 与叶面积指数(LAI)线性相关度更高;且当地表湿度较高时, EVI2比NDVI的效果要好很多[35], 能更好地反映出高植被覆盖区的植被生长状况。此外, 在提取物候信息时, 理论上数据的时间分辨率越高所获得的物候反演数据越精确。但是, 高时间分辨率数据往往需要附加数据和降噪技术来完成数据重构, 而过粗的时间分辨率数据又可能会错过一些关键的物候变化时期[36-38]。由此来看, EVI2对于物候研究而言其时空间分辨率(250m, 8d)目前相对更适合。
秦岭山地被称为“中国的脊梁”, 是我国重要的地理分界线与气候分界线, 秦岭南北气候差异明显, 植被垂直分带明显。秦岭也被认为是我国中部地区典型的生态过渡带、生态敏感区, 以及重要的气候敏感区[39-40]。近半个世纪以来, 秦岭地区气温呈显著上升趋势, 自1980s以来增温尤为显著, 且秦岭南北增温表现出北高南低的区域差异[41, 14]。气候变暖背景下, 开展秦岭山地植被物候变化特征及其南北差异研究具有重要意义。然而, 秦岭山地地面观测物候数据缺失, 虽有学者采用遥感植被指数NDVI、EVI对秦岭山地植物物候变化做了初步研究[42-43, 33], 但以植被指数EVI2开展秦岭植被物候变化南北差异对比研究的尚不多见。鉴于此, 本研究基于2000—2017年MODIS EVI2数据, 反演了秦岭山地的植被物候期参数并建立了其遥感物候数据集, 分析了近18年来秦岭山地植被物候的时空格局变化特征及其南北差异。试图回答:气候变化背景下, 近18年来秦岭山地植被物候有何变化特征?南北坡的植被物候变化表现出怎样的差异性?植被物候沿南北坡的海拔梯度呈现怎样的变化规律?以及南北坡的不同植被垂直带上植被物候变化又有何特征?研究旨在揭示气候变化背景下秦岭山地过渡带对气候变化的响应方式, 以期为秦岭山地的生态环境保护、生态安全评价以及应对气候变化策略提供科学依据和决策支持, 为深入理解全球变化的事实及其对山地植被生态系统的影响等科学问题提供基础资料。
1 数据与方法 1.1 研究区概况及其植被垂直带分布秦岭是全球生物多样性关键地带之一, 是我国最具有自然地理分界意义的山脉, 亦是我国重要的生态安全屏障, 其在我国具有无可比拟的特殊地位及重要意义。由于秦岭山体对北进的东亚暖湿气流及南侵的北方干冷气流具有巨大的阻挡作用, 南北气候差异显著, 表现为北干南湿, 北坡属于暖温带半湿润半干旱气候, 1月年均温低于0℃, 广布暖温带针阔混交林与落叶阔叶林;而南坡为北亚热带湿润气候, 1月年均温则高于0℃, 以常绿落叶阔叶混交林为基带。本研究以秦岭的核心组成部分陕西秦岭为研究区(图 1), 以山脊线作为秦岭的南北分界线。

|
| 图 1 研究区地理位置及高程分布 Fig. 1 Geographical location and elevation distribution of the study area |
秦岭植被类型丰富, 植被垂直带谱完整而复杂。依据学者们对秦岭植被垂直带谱划分的相关研究结果[44], 秦岭植被垂直带的空间分布如图 2所示。秦岭植被垂直带谱南北坡差异明显, 自下而上, 北坡依次为落叶阔叶林(600—1500m)——针阔混交林(1500—2400m)——针叶林(2400—3100m)——高山灌丛草甸和落叶针叶林(>3100m);南坡依次为常绿、落叶阔叶混交林(600—1300m)——针阔混交林(1300—2200m)——针叶林(2200—3000m)——高山灌丛草甸(>3000m)。秦岭山脊两侧呈现南草北树的景观差异, 是由秦岭南北气候差异造成的。

|
| 图 2 秦岭植被垂直带空间分布 Fig. 2 Spatial distribution of the vertical vegetation belts in the Qinling Mountains |
本研究选取时间分辨率为8d及空间分辨率250m的MODIS EVI2植被指数作为监测遥感物候变化的数据源, 其来源于NASA的MODIS数据产品中MOD09Q1地表反射率数据集(https://ladsweb.nascom.nasa.gov), 时段为2000—2017年。
首先采用最大值合成法(MVC)消除太阳高度角、大气、云、气溶胶等对EVI2数据的影响, 并通过ENVI 5.3软件的波段运算工具计算得到该地区的增强植被指数EVI2(enhanced vegetation index 2), 其计算公式如下:
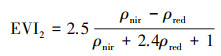
|
式中:ρnir和ρred分别为近红外反射率和红外波段反射率。
之后, 采用时间序列谐波分析法(HANTS)进一步对EVI2时序数据进行去噪、平滑滤波处理, 以去除亚像元内残余云及云霾或其他负面影响。
1.2.2 遥感物候反演方法时序数据能够反映植被在一年中的变化状况, 与植被的物候特征相联系, 因此可以通过提取植被指数EVI2的关键指标来确定植被物候变化。研究以物候始期(Growing Season Start, GSS)、末期(Growing Season End, GSE)及生长期(Growing Season Length, GSL)三个参数作为研究区植被物候的判识指标。采用年日序(Day of Year, DOY)表示物候期出现的时间, 即以当年1月1日开始的实际日数。
目前利用遥感提取物候的常用方法有:滑动平均法、阈值法、最大比率法、阈值法和最大比率法结合等, 但尚无一种方法被广泛接受。本研究采用阈值法与最大比率法相结合的方法进行物候期参数的提取。过程如下:①确定GSS与GSE的阈值。计算HANTS平滑后日EVI2时间序列的多年均值, 可获得每个像元上EVI2多年平均的年内变化曲线;计算基于像元尺度两相邻时间点上的EVI2差值, 即相对变化值, 利用相对变化值的最大绝对值确定GSS与GSE的EVI2阈值。②提取每年的GSS与GSE。在TIMESAT软件中选取双Logistic模型曲线拟合法重建EVI2时间序列拟合曲线, 以确定的EVI2阈值分别提取每年的GSS与GSE。③设定置信区间剔除异常值并计算GSL。在ArcGIS软件中, 设置1%的置信区间剔除提取的GSS与GSE异常值, 进一步计算每年GSS与GSE的差值, 即为该年的GSL。
1.3 分析方法空间尺度的一元线性回归趋势分析法, 是采用最小二乘法逐像元拟合EVI2所确定的GSS、GSE及GSL的斜率, 其能够模拟每个栅格的变化趋势, 综合反映区域植被的时空格局演变[45], 公式为:
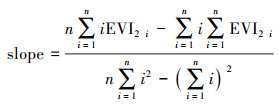
|
式中:n为时间序列(年);EVI2i为第i年的GSS、GSE及GSL值;当slope>0时,表明GSS与GSE呈推迟趋势,GSL呈延长趋势; 当slope<0时,表明GSS与GSE呈提前趋势,GSL呈缩短趋势。采用t检验变化趋势的显著性, 并将结果划分为极显著(P≤0.01)、显著(0.01 < P≤0.05)、弱显著(0.05 < P≤0.1)和不显著(P>0.1)变化4个等级。
4 结果与分析 4.1 秦岭山地遥感GSS与GSE的EVI2阈值确定及精度验证植物物候期识别所需的各项参数均应根据其多年的时间演化规律进行确定, 这样能够提高物候遥感识别方法的鲁棒性。图 3为2000—2017年秦岭山地EVI2年均值最大相对变化值确定的GSS与GSE阈值。结果显示, GSS的EVI2阈值为0.264, 其对应一年中93DOY, 即秦岭山地物候始期平均发生于4月初;GSE的EVI2阈值为0.289, 其对应一年中277DOY, 即秦岭山地物候末期平均发生于10月上旬。

|
| 图 3 2000—2017年秦岭山地EVI2多年均值确定的GSS与GSE阈值 Fig. 3 Thresholds of the GSS and GSE determined by the multi-year mean of EVI2 in the Qinling Mountains from 2000 to 2017 GSS:物候始期 Growing season start; GSE: 物候末期 Growing season end |
秦岭地区的地面物候观测数据甚少, 目前可用于验证以上结论的相关研究仅是学者们采用中国物候观测网中秦岭北麓西安站的木本植物观测数据分析得到的结论[33, 42-43, 46-48]。此外, 研究者本人以研究区日均温10℃气温阈值反演的气候生长期并采用ANUSPLIN法进行空间插值获得的2000—2015年相关结果(见图 4);以及学者们采用遥感监测的植被指数NDVI、EVI反演的物候期分析结果[33, 42-43], 以上三类不同意义的物候研究情况见表 1。

|
| 图 4 2000—2015年秦岭山地气候生长期GSS与GSE的多年均值空间分布 Fig. 4 Spatial distribution of the multi-year mean of the GSS and GSE represented by climate in the Qinling Mountains from 2000 to 2015 |
| 物候监测类型 Types of phenology monitored |
数据类型 Types of data |
作者 Scholars |
研究区域 Study area |
时期 Periods |
始期发生时间 Time of start phenophases |
末期发生时间 Time of end phenophases |
| 传统地面观测 Traditional ground observation |
观测树种 | 邓晨晖等[46] | 西安站 | 1985—2015 | 97DOY 展叶盛期 |
316DOY 叶全变色期 |
| 2000—2015 | 93DOY 展叶盛期 |
322DOY 叶全变色期 |
||||
| 李淑娟和刘雅莉[47] | 西安植物园及曲江风景区各大公园 | 1988—2010 | 3月(37.7%) 4月(61.6%) 开始展叶期 | 9月(67.6%) 10月(32.4%) 叶开始变色期 | ||
| 贺映娜[42] | 西安站 | 1963—1989 | 99.5DOY 展叶盛期 |
301.6DOY 叶全变色期 |
||
| 1990—2008 | 97.4DOY 展叶盛期 |
308.6DOY 叶全变色期 |
||||
| 陈效逑等[48] | 西安站 | 1982—1996 | 2月底—3月67DOY变绿期 | 9月初—10月初257DOY凋零期 | ||
| 气温反演 Estimation based on temperature data |
10℃气温 | 邓晨晖 | 陕西秦岭 | 2000—2015 | 88.44DOY | 298.30DOY |
| 遥感监测 | MODIS NDVI | 马新萍等[43] | 陕西秦岭 | 2000—2010 | 第7—9旬 | 第32—34旬 |
| Remote sensing monitoring | MODIS EVI | 夏浩铭v[33] | 广义秦岭 | 2001—2010 | 81—120DOY | 270—311DOY |
| SPOT NDVI | 贺映娜等[42] | 陕西秦岭 | 1998—2009 | 96.8DOY | 309.0DOY |
由表 1可知, 秦岭地区三类不同意义的研究结果显示, 物候始末期发生时间较为一致, 始期主要发生于4月前后, 末期主要发生于10月前后。但是, 由于存在数据源、时间尺度、物候期时期, 以及提取物候期方法等不同, 学者们对于秦岭地区的物候研究结果也存在一定差异。通过对比发现, 本研究所提取的遥感物候期与以上研究所得的物候始末期发生时间也较为一致, 表明基于MODIS EVI2数据反演的秦岭山地物候期数据对于该区域的物候监测研究具有一定的参考价值。
4.2 植被物候空间分布及年际变化特征 4.2.1 植被物候空间分布特征图 5为2000—2017年秦岭山地植被物候生长期参数的多年均值空间分布。由图 5可见, 近18年来, 秦岭山地植被物候变化表现出明显的地形和气候地域分异规律。全区GSS主要发生于70—130DOY, 平均为107.0DOY, 但在秦岭北坡的地势低平区早于70DOY;而在一些海拔和纬度较高的地区GSS则晚于130DOY。北坡GSS平均发生于102.1DOY, 而南坡为108.3DOY, 北坡较南坡平均约早6.2d。全区GSE主要发生于270—310DOY, 平均为292.3DOY, 但在南北坡地势低平人口密集区却早于270DOY, 也表现出一定的海拔及纬度差异。北坡GSE平均发生于287.7DOY, 南坡为293.5DOY, 北坡较南坡平均约早5.8d。全区GSL主要集中在150—210d, 平均为185.3d, 北坡约114.6—249.7d, 南坡约91.4—231.0d。南北坡GSL平均长度基本相同, 但就总体来看, 北坡较南坡的长, 北坡较南坡约长18.7—23.2d。

|
| 图 5 2000—2017年秦岭山地植被物候期参数的多年均值空间分布 Fig. 5 Spatial distribution of the multi-year mean of vegetation phenological parameters in the Qinling Mountains from 2000 to 2017 |
图 6为2000—2017年秦岭山地植被物候期参数的年际变化趋势及显著性。由图 6可见, 近18年来, 秦岭山地植被物候年际变化存在明显的空间差异。

|
| 图 6 2000—2017年秦岭山地植被物候期参数的年际变化趋势及显著性 Fig. 6 Spatial distribution of interannual change trends and significance in the vegetation phenological parameters in the Qinling Mountains from 2000 to 2017 |
GSS全区以提前趋势为主, 平均速率为-3.7d/10a, 其中83.29%的区域呈提前趋势, 16.71%的区域呈推迟趋势;且以提前0—5d/10a(44.46%)为主, 提前5—10d/10a(28.60%)次之。t检验结果显示, 提前趋势达到显著性(P < 0.1, 下同)的区域仅占25.41%, 主要位于山脊线附近的高海拔区及南坡的西部地区;推迟趋势达到显著性的区域仅占3.57%, 主要位于北坡的低海拔区, 这可能与近些年频发的“倒春寒”气候对北坡影响较大有关。北坡提前趋势达到显著性的区域仅11.65%与不显著的区域为53.00%;而南坡分别为29.03%、59.18%。
GSE全区变化趋势不明显, 其中50.17%的区域呈推迟趋势, 49.83%的区域呈提前趋势;且推迟趋势以0—5d/10a(25.51%)、5—10d/10a(15.44%)为主, 提前趋势亦以0—5d/10a(22.82%)、5—10d/10a(14.81%)为主。推迟趋势达到显著性的区域仅7.49%, 主要位于北坡人口密集的低海拔区, 可能与城市热岛效应的有关;提前趋势达到显著性的区域仅5.08%。北坡推迟趋势达到显著性的区域为20.45%与不显著的区域为57.40%;而南坡分别为4.10%、38.83%。
GSL全区以延长趋势为主, 平均速率为3.7d/10a, 其中65.34%的区域呈延长趋势, 34.66%的区域呈缩短趋势, 延长在0—5d/10a(19.28%)、5—10d/10a(20.71%)及10—15d/10a(14.12%)均有分布, 缩短趋势以0—5d/10a(14.83%)与5—10d/10a(10.15%)的占比相对较多。延长趋势达到显著性的区域仅占12.06%, 主要位于山脊线附近的高海拔区, 这与学者们近年来研究发现高海拔区增温较低海拔区更为显著的结论一致[49];缩短趋势达到显著性的区域仅占2.17%。北坡延长趋势达到显著性的区域为12.74%与不显著的区域为63.66%, 而南坡分别为11.89%、50.57%。
综上可知, GSL延长主要是GSS提前的结果, 由此可知秦岭山地植被生长期延长主要归因于物候始期的提前, 物候始期对气候变化的响应程度显著大于物候末期。GSS的提前趋势南坡较北坡显著, GSE的推迟趋势北坡较南坡显著, 可知秦岭南北坡的植被物候变化不仅表现出显著的区域差异而且存在显著的季节性差异。与此同时, 秦岭山地植被物候在高海拔区的变化趋势更为显著。
4.3 植被物候沿海拔梯度变化特征2000—2017年秦岭山地植被物候期多年均值与海拔的关系显示(图 7), GSS、GSE及GSL随海拔升高呈规律性变化, 表现出显著的海拔敏感性。随着海拔升高, GSS逐渐推迟, GSE逐渐提前, GSL逐渐缩短。在海拔≤600m平原区, 三者随海拔变化的波动幅度较大, 规律不明显, 这可能与人类活动有关, 低海拔区受城市热岛效应的影响区域增温差异明显;而在海拔≥2700m高海拔区, 三者随海拔变化的波动幅度相对更为剧烈, 这与气候条件密切相关, 这一区域为冰缘气候所控制, 气温低且多强风, 山顶效应明显。

|
| 图 7 2000—2017年秦岭山地及其南北坡植被物候期参数多年均值与海拔的关系 Fig. 7 Changing trends in the multi-year mean of vegetation phenological parameters along altitude in the Qinling Mountains and its north and South Slopes from 2000 to 2017 |
秦岭南北坡2000—2017年植被物候期多年均值与海拔的关系显示(图 7), 北坡GSS与GSL随海拔上升的变化幅度大于南坡, 而GSE则表现为南坡大于北坡。海拔每上升100m, 北坡GSS推迟1.76d, GSE提前0.25d, GSL缩短2.01d;南坡GSS推迟1.50d, GSE提前0.44d, GSL缩短1.94d, 这可能与秦岭山地北坡陡短而南坡缓长的地形差异有关。就GSS而言, 在海拔 < 900m地区北坡的变化幅度波动较大且发生时间早于南坡;而在≥2600m地区南北坡均随海拔上升推迟趋势减缓, 且北坡早于南坡;在900—2600m地区南北坡的发生时间相差不大且均随海拔上升呈规律性推迟趋势。就GSE而言, 南北坡的发生时间随海拔变化波动均较大, 在海拔≥900m地区表现出随海拔上升呈规律性提前趋势;而在≥3000m地区变化趋势发生转折, 均表现出随海拔上升呈明显的推迟, 且在3300m之后推迟趋势均有所减缓;在≤1000m及2300—3000m地区则是南坡的发生时间晚于北坡。就GSL而言, 海拔≤600m地区北坡的变化幅度波动大且较南坡长;在600—2800m地区表现出随海拔上升呈规律性缩短趋势且南北坡生长期相差不大;而在≥2800m地区则随海拔上升呈缓慢的延长趋势且北坡GSL长于南坡。
4.4 植被垂直带上物候变化特征依据秦岭植被垂直带的划分情况, 进一步分析了秦岭南北坡的不同植被垂直带上遥感物候变化特征(见表 2)。表 2显示, 落叶阔叶林带与高山灌丛草甸带GSS北坡较南坡分别平均早4.2d、3.5d, 而针阔混交林带与针叶林带GSS则是北坡较南坡分别平均晚6.0d、1.6d。就GSE而言, 除南北坡的≤600m植被带及高山灌丛草甸带外, 落叶阔叶林带、针阔混交林带及针叶林带北坡较南坡依次分别平均早4.7d、4d、1.2d, 而高山灌丛草甸带北坡较南坡平均晚2.9d;落叶阔叶林带、针阔混交林带、针叶林带GSL北坡较南坡依次分别平均短0.4d、10.0d、2.8d, 而高山灌丛草甸带北坡较南坡平均长6.4d。
| 植被带 Vegetation belts |
海拔范围/m Altitude range |
北坡 Northern slope |
植被带 Vegetation belts |
海拔范围/m Altitude range |
南坡 Southern slope |
|||||
| GSS/ DOY | GSE/ DOY | GSL/ days | GSS/ DOY | GSE/ DOY | GSL/ days | |||||
| ≤600 | 82.8 | 282.8 | 200.0 | ≤600 | 103.2 | 288.2 | 185.0 | |||
| 落叶阔叶林 Deciduous broadleaved forest |
600—1500 | 99.8 | 292.3 | 192.6 | 常绿落叶阔叶混交林 Evergreen deciduous broadleaved mixed forest |
600—1300 | 104.0 | 297.0 | 193.0 | |
| 针阔混交林 Coniferous and broadleaved mixed forest |
1500—2400 | 120.9 | 286.6 | 165.7 | 针阔混交林 Coniferous and broadleaved mixed forest |
1300—2200 | 114.9 | 290.6 | 175.7 | |
| 针叶林 Coniferous forest |
2400—3100 | 133.5 | 277.0 | 143.5 | 针叶林 Coniferous forest |
2200—3000 | 131.9 | 278.2 | 146.3 | |
| 高山灌丛草甸 Alpine bush and meadow |
>3100 | 135.7 | 288.6 | 152.9 | 高山灌丛草甸 Alpine bush and meadow |
>3000 | 139.2 | 285.7 | 146.5 | |
| GSS: 物候始期 Growing season start; GSE: 物候末期 Growing season end; GSL: 物候生长期 Growing season length | ||||||||||
三者的变化在≤600m植被带上表现出差异性, 可能与北坡低海拔区平缓, 人口密度大, 人类活动对北坡早春气温回升的影响大于南坡有关;而在高山灌丛草甸带的不同, 一方面可能是因为高海拔区增温幅度更显著, 另一方面也可能与受气候变暖的影响林线树种太白红杉上限已伸入灌丛草甸区有关。
三者在高山灌丛草甸带发生时间及时长南坡与北坡发生转换的原因, 可能归于山顶南北两侧气候的差异, 南坡向阳, 土壤干燥, 水分蒸发快, 而北坡背阴, 阳光照射时间短, 水分蒸发慢, 土壤相对湿润, 故在高山草甸区北坡的气候条件更利于植物生长, 更利于物候始期提前与末期推迟。
5 结论与讨论 5.1 讨论为寻求遥感物候与实测物候之间的关系, 绘制同期(2000—2015年)遥感物候与实测物候、气候表征的物候的GSS与GSE变化趋势图(见图 8)。由图 8可知, 虽然三种意义表征的物候期变化均表现出GSS提前及GSE推迟, 但变化趋势并不完全一致, 尤其是秋季物候。GSS遥感物候与实测物候的相关性为0.540(P<0.05), 其与气候表征的物候的GSS相关性为0.519(P<0.05), 均达到了显著性水平;而GSE遥感物候与其他两类物候的相关性均为达到显著性水平。实际上, 实测物候与遥感物候之间本身就因观测尺度与观测内容的不同而存在明显的内涵差异, 二者所监测的物候期并不是明确的对应关系[17], 而气候表征的物候虽然能够实现较大空间尺度的生长期变化研究, 但存在仅考虑了气温因素的自身局限性。因此, 如何找到一种合理科学的方法, 建立起实测物候与遥感物候之间转换的纽带, 实现二者之间监测尺度和监测内容的匹配, 从而实现实测物候与遥感物候的结合, 将地面个体水平上观测的植物物候期扩展到遥感监测的植被群落甚至生态系统尺度上, 实现物候研究尺度从点到面的区域扩展、时间序列的延长、分辨率的精度更高, 将有望获得更准确的研究结果。这仍然是物候研究的重要内容。

|
| 图 8 2000—2015年秦岭遥感物候与实测物候、气候表征的物候GSS与GSE的变化趋势 Fig. 8 Changing trends in the GSS and GSE of phenology by remote sensing monitoring and ground observation, phenology represented by climate in the Qinling Mountains from 2000 to 2015 |
研究结果显示, 秦岭南北坡的同一类型植被带上物候变化存在明显差异, 这不仅与南北气候差异有关, 而且与植被带内的树种差异有关。由于秦岭南北气候的差异, 南北坡植被垂直带谱呈现明显的分异现象, 主要表现在各植被垂直带的分布高度及基带特征[44]。北坡基带为落叶阔叶林, 主要分布于海拔600—1500m的地区, 以栓皮栎林为代表;南坡基带为常绿落叶阔叶混交林, 主要分布于海拔600—1300m的地区, 除栓皮栎、麻栎外, 常有常绿树种小青冈和岩栎等混生其中[44]。此外, 北坡有较明显的辽东栎林亚带, 而南坡则没有, 辽东栎林亚带出现与否被认为是秦岭南北坡植被垂直带差异明显的标志之一[44]。
此外, 研究亦发现气候变暖背景下秦岭山地植被物候变化的显著区域主要集中在人为影响较少的高海拔区, 这一结论亦从植被物候变化视角证实了学者们的研究发现, 全球变化背景下山地对气候变化具有高度的敏感性, 其中高山带对气候变化的响应更敏感, 高海拔地区对全球变暖的响应较低海拔区更强烈[50-56]。再者, 高海拔区亦是秦岭天然林连片分布区, 气候变暖导致秦岭高山林线波动上升, 植被带上限向更高海拔区上升, 各植被带内植物适生区扩大。与此同时, 秦岭高山区特有物种的太白红杉为适应生境变化正向高山灌丛、裸岩分布处迁移, 林线附近的幼龄林随处可见;而巴山冷杉却出现大面积退化甚至死亡现象, 尤其是在其上限区域更为严重。那么, 山地高海拔区变暖更为显著的原因是什么?有关机理仍有待进一步深入探究。
5.2 结论(1) 近18年来, 秦岭山地植被物候变化表现出明显的地形和气候的地域分异规律。全区GSS主要发生于70—130DOY, 平均为107.0DOY, 北坡较南坡约早6.2d;GSE主要发生于270—310DOY, 平均为292.3DOY, 南坡较北坡约晚5.8d。全区GSL主要集中在150—210d, 平均为185.3d, 南北坡GSL平均长度基本相同, 但时长范围相差较大, 北坡约114.6—249.7d, 南坡约91.4—231.0d, 北坡较南坡约长18.7—23.2d。
(2) 近18年来, 秦岭山地植被物候变化趋势存在明显的空间差异, 尤以高海拔区的变化最为显著, 物候始期对气候变化的响应程度显著大于末期, 且南北坡的变化不仅存在区域差异且存在季节差异。GSS全区以提前趋势为主, 83.29%的区域提前, 平均速率为-3.7d/10a, 提前主要集中在0—5d/10a(44.46%)及5—10d/10a(28.60%), 南坡的提前趋势较北坡显著;GSE全区变化趋势不明显, 50.17%的区域推迟, 49.83%的区域提前, 北坡的推迟趋势较南坡显著;GSL全区以延长趋势为主, 65.34%的区域延长, 平均速率为3.7d/10a, 延长在0—5d/10a(19.28%)、5—10d/10a(20.71%)及10—15d/10a(14.12%)均有分布, 北坡的延长趋势较南坡显著。
(3) GSS、GSS及GSS随海拔升高呈规律性变化, 表现出显著的海拔敏感性, 南北坡三者随海拔上升的变化幅度呈现明显的差异。随着海拔升高, GSS逐渐推迟, GSE逐渐提前, GSL逐渐缩短。在海拔≤600m盆地平原区, 三者随海拔变化的波动幅度较大, 规律不明显, 在海拔≥2700m高海拔区, 三者随海拔升高变化的波动幅度相对更为剧烈。海拔每上升100m, 北坡GSS推迟1.76d, GSE提前0.25d, GSL缩短2.01d;南坡GSS推迟1.50d, GSE提前0.44d, GSL缩短1.94d。
(4) GSS、GSE及GSL在南北坡植被垂直带上的变化存在明显的差异, 尤以≤600m植被带上及高山灌丛草甸带上的差异最为明显, 且三者在高山灌丛草甸带发生时间及时长南坡与北坡发生转换。GSS落叶阔叶林带与高山灌丛草甸带北坡较南坡分别平均早4.2d、3.5d, 而针阔混交林带与针叶林带北坡较南坡分别平均晚6.0d、1.6d。GSE落叶阔叶林带、针阔混交林带、针叶林带南坡较北坡依次分别平均晚4.7d、4d、1.2d, 而高山灌丛草甸带北坡较南坡平均晚2.9d;GSL落叶阔叶林带、针阔混交林带、针叶林带南坡较北坡依次分别平均长0.4d、10.0d、2.8d, 而高山灌丛草甸带北坡较南坡平均长6.4d。
| [1] |
King D A. Climate change science: adapt, mitigate, or ignore?. Science, 2004, 303(5655): 176-177. DOI:10.1126/science.1094329 |
| [2] |
Xia J Y, Chen J Q, Piao S L, Ciais P, Luo Y Q, Wan S Q. Terrestrial carbon cycle affected by non-uniform climate warming. Nature Geoscience, 2014, 7(3): 173-180. DOI:10.1038/ngeo2093 |
| [3] |
王根绪, 邓伟, 杨燕, 程根伟. 山地生态学的研究进展、重点领域与趋势. 山地学报, 2011, 29(2): 129-140. DOI:10.3969/j.issn.1008-2786.2011.02.001 |
| [4] |
Zhang B P, Yao Y H. Implications of mass elevation effect for the altitudinal patterns of global ecology. Journal of Geographical Sciences, 2016, 26(7): 871-877. DOI:10.1007/s11442-016-1303-2 |
| [5] |
Zhu Z C, Piao S L, Myneni R B, Huang M T, Zeng Z Z, Canadell J G, Ciais P, Sitch S, Friedlingstein P, Arneth A, Cao C X, Cheng L, Kato E, Koven C, Li Y, Lian X, Liu Y W, Liu R G, Mao J F, Pan Y Z, Peng S S, Peñuelas J, Poulter B, Pugh T A M, Stocker B D, Viovy N, Wang X H, Wang Y P, Xiao Z Q, Yang H, Zaehle S, Zeng N. Greening of the Earth and its drivers. Nature Climate Change, 2016, 6(8): 791-795. DOI:10.1038/nclimate3004 |
| [6] |
Barr A, Black T A, McCaughey H. Climatic and phenological controls of the carbon and energy balances of three contrasting boreal forest ecosystems in western Canada//Noormets A, ed. Phenology of Ecosystem Processes. New York: Springer, 2009: 3-34.
|
| [7] |
Root T L, Price J T, Hall K R, Schneider S H, Rosenzweig C, Pounds J A. Fingerprints of global warming on wild animals and plants. Nature, 2003, 421(6918): 57-60. DOI:10.1038/nature01333 |
| [8] |
孔冬冬, 张强, 黄文琳, 顾西辉. 1982-2013年青藏高原植被物候变化及气象因素影响. 地理学报, 2017, 72(1): 39-52. |
| [9] |
朱连奇, 许立民. 全球变化对陆地生态系统的影响研究. 地域研究与开发, 2011, 30(2): 161-164. DOI:10.3969/j.issn.1003-2363.2011.02.034 |
| [10] |
穆少杰, 李建龙, 陈奕兆, 刚成诚, 周伟, 居为民. 2001-2010年内蒙古植被覆盖度时空变化特征. 地理学报, 2012, 67(9): 1255-1268. |
| [11] |
张晓东, 朱文博, 张静静, 朱连奇, 赵芳, 崔耀平. 伏牛山地森林植被物候及其对气候变化的响应. 地理学报, 2018, 73(1): 41-53. |
| [12] |
方修琦, 余卫红. 物候对全球变暖响应的研究综述. 地球科学进展, 2002, 17(5): 714-719. DOI:10.3321/j.issn:1001-8166.2002.05.013 |
| [13] |
莫非, 赵鸿, 王建永, 强生才, 周宏, 王绍明, 熊友才. 全球变化下植物物候研究的关键问题. 生态学报, 2011, 31(9): 2593-2601. |
| [14] |
白红英. 秦巴山区森林植被对环境变化的响应. 北京: 科学出版社, 2014.
|
| [15] |
Orlandi F, Ruga L, Romano B, Fornaciari M. Olive flowering as an indicator of local climatic changes. Theoretical and Applied Climatology, 2005, 81(3/4): 169-176. DOI:10.1007/s00704-004-0120-1 |
| [16] |
Donnelly A, Jones M B, Sweeney J. A review of indicators of climate change for use in Ireland. International Journal of Biometeorology, 2004, 49(1): 1-12. |
| [17] |
范德芹, 赵学胜, 朱文泉, 郑周涛. 植物物候遥感监测精度影响因素研究综述. 地理科学进展, 2016, 35(3): 304-319. |
| [18] |
Walther A, Linderholm H W. A comparison of growing season indices for the Greater Baltic Area. International Journal of Biometeorology, 2006, 51(2): 107-118. DOI:10.1007/s00484-006-0048-5 |
| [19] |
Linderholm H W, Walther A, Chen D L. Twentieth-century trends in the thermal growing season in the Greater Baltic Area. Climatic Change, 2008, 87(3/4): 405-419. DOI:10.1007/s10584-007-9327-3 |
| [20] |
郭灵辉, 吴绍洪, 赵东升, 冷国勇, 张清雨. 近50年内蒙古地区生长季变化趋势. 地理科学, 2013, 33(4): 505-512. |
| [21] |
郑景云, 葛全胜, 郝志新. 气候增暖对我国近40年植物物候变化的影响. 科学通报, 2002, 47(20): 1582-1587. DOI:10.3321/j.issn:0023-074X.2002.20.013 |
| [22] |
张文霞, 刘普幸, 冯青荣, 汪天广, 王天强. 1960-2015年中国绿洲胡杨生长季对全球变暖的时空响应及原因. 地理学报, 2017, 72(7): 1151-1162. |
| [23] |
Moulin S, Kergoat L, Viovy N, Dedieu G. Global-scale assessment of vegetation phenology using NOAA/AVHRR satellite measurements. Journal of Climate, 1997, 10(6): 1154-1170. DOI:10.1175/1520-0442(1997)010<1154:GSAOVP>2.0.CO;2 |
| [24] |
Karlsen S R, Høgda K A, Wielgolaski F E, Tolvanen A, Tømmervik H, Poikolainen J, Kubin E. Growing-season trends in Fennoscandia 1982-2006, determined from satellite and phenology data. Climate Research, 2009, 39: 275-286. DOI:10.3354/cr00828 |
| [25] |
Neil K L, Landrum L, Wu J G. Effects of urbanization on flowering phenology in the metropolitan Phoenix region of USA: Findings from herbarium records. Journal of Arid Environments, 2010, 74(4): 440-444. DOI:10.1016/j.jaridenv.2009.10.010 |
| [26] |
Jeong J H, Ho C H, Linderholm H W, Jeong S J, Chen D, Choi Y S. Impact of urban warming on earlier spring flowering in Korea. International Journal of Climatology, 2011, 31(10): 1488-1497. DOI:10.1002/joc.2178 |
| [27] |
丁明军, 张镱锂, 孙晓敏, 刘林山, 王兆锋. 近10年青藏高原高寒草地物候时空变化特征分析. 科学通报, 2012, 57(33): 3185-3194. |
| [28] |
Myneni R B, Keeling C D, Tucker C J, Asrar G, Nemani R R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature, 1997, 386(6626): 698-702. DOI:10.1038/386698a0 |
| [29] |
Runtunuwu E, Kondoh A, Wikantika K, et al. NDVI-derived length of the growth period estimations for different vegetation types in Monsoon Asia. IECI Chapter Japan Series, 2001, 3(1): 106-109. |
| [30] |
王宏, 李晓兵, 莺歌, 王丹丹, 龙慧灵. 基于NOAA NDVI的植被生长季模拟方法研究. 地理科学进展, 2006, 25(6): 21-32. DOI:10.3969/j.issn.1007-6301.2006.06.003 |
| [31] |
Piao S L, Fang J Y, Zhou L M, Ciais P, Zhu B. Variations in satellite-derived phenology in China's temperate vegetation. Global Change Biology, 2006, 12(4): 672-685. DOI:10.1111/j.1365-2486.2006.01123.x |
| [32] |
于信芳, 庄大方. 基于MODIS NDVI数据的东北森林物候期监测. 资源科学, 2006, 28(4): 111-117. DOI:10.3321/j.issn:1007-7588.2006.04.023 |
| [33] |
夏浩铭, 李爱农, 赵伟, 边金虎, 雷光斌. 2001-2010年秦岭森林物候时空变化遥感监测. 地理科学进展, 2015, 34(10): 1297-1305. |
| [34] |
Rocha A V, Shaver G R. Advantages of a two band EVI calculated from solar and photosynthetically active radiation fluxes. Agricultural and Forest Meteorology, 2009, 149(9): 1560-1563. DOI:10.1016/j.agrformet.2009.03.016 |
| [35] |
Huete A, Didan K, Miura T, Rodriguez E P, Gao X, Ferreira L G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing of Environment, 2002, 83(1/2): 195-213. |
| [36] |
马勇刚, 张弛, 陈曦. 利用遥感数据优化物候模型时样本选择的新方法. 植物生态学报, 2015, 39(3): 264-274. |
| [37] |
Zhao H, Yang Z W, Di L P, Pei Z Y. Evaluation of temporal resolution effect in remote sensing based crop phenology detection studies//Li D L, Chen Y Y, eds. Computer and Computing Technologies in Agriculture V. Berlin: Springer, 2012: 135-150.
|
| [38] |
Zhang X Y, Friedl M A, Schaaf C B. Sensitivity of vegetation phenology detection to the temporal resolution of satellite data. International Journal of Remote Sensing, 2009, 30(8): 2061-2074. DOI:10.1080/01431160802549237 |
| [39] |
白红英, 马新萍, 高翔, 侯钦磊. 基于DEM的秦岭山地1月气温及0℃等温线变化. 地理学报, 2012, 67(11): 1443-1450. DOI:10.11821/xb201211001 |
| [40] |
刘洪滨, 邵雪梅. 采用秦岭冷杉年轮宽度重建陕西镇安1755年以来的初春温度. 气象学报, 2000, 58(2): 223-233. DOI:10.3321/j.issn:0577-6619.2000.02.010 |
| [41] |
高翔, 白红英, 张善红, 贺映娜. 1959-2009年秦岭山地气候变化趋势研究. 水土保持通报, 2012, 32(1): 207-211. |
| [42] |
贺映娜. 秦岭植被物候期及遥感生长季的变化研究[D]. 西安: 西北大学, 2012.
|
| [43] |
马新萍, 白红英, 贺映娜, 秦进. 基于NDVI的秦岭山地植被遥感物候及其与气温的响应关系——以陕西境内为例. 地理科学, 2015, 35(12): 1616-1621. |
| [44] |
岳明. 秦岭植物垂直带谱完整复杂. 森林与人类, 2015(2): 76-81. |
| [45] |
宋怡, 马明国. 基于GIMMS AVHRR NDVI数据的中国寒旱区植被动态及其与气候因子的关系. 遥感学报, 2008, 12(3): 499-505. |
| [46] |
邓晨晖, 白红英, 翟丹平, 高山, 黄晓月, 孟清, 贺映娜. 气候变化背景下1964-2015年秦岭植物物候变化. 生态学报, 2017, 37(23): 7882-7893. |
| [47] |
李淑娟, 刘雅莉. 西安主要季色叶植物观赏特征及物候图谱研究初报. 西北林学院学报, 2013, 28(2): 42-47. DOI:10.3969/j.issn.1001-7461.2013.02.08 |
| [48] |
陈效逑, 韩建伟. 我国东部温带植物群落的季相及其时空变化特征. 植物生态学报, 2008, 32(2): 336-346. DOI:10.3773/j.issn.1005-264x.2008.02.010 |
| [49] |
刘晓东, 晏利斌, 程志刚, 李新周. 中低纬度高原山地气候变暖对海拔高度的依赖性. 高原山地气象研究, 2008, 28(1): 19-23. DOI:10.3969/j.issn.1674-2184.2008.01.003 |
| [50] |
Huber U M, Bugmann H K M, Reasoner M A. Global Change and Mountain Regions: An Overview of Current Knowledge. Berlin: Springer, 2005.
|
| [51] |
UNGA. Resolution A/RES/58/216 on sustainable development in mountain regions, adopted by the general assembly. New York: United Nations Headquarters, 2004.
|
| [52] |
姚檀栋, 刘晓东, 王宁练. 青藏高原地区的气候变化幅度问题. 科学通报, 2000, 45(1): 98-106. DOI:10.3321/j.issn:0023-074X.2000.01.021 |
| [53] |
丁明军, 李兰晖, 张镱锂, 刘林山, 王兆锋. 1971-2012年青藏高原及周边地区气温变化特征及其海拔敏感性分析. 资源科学, 2014, 36(7): 1509-1518. |
| [54] |
Fyfe J C, Flato G M. Enhanced climate change and its detection over the Rocky Mountains. Journal of Climate, 1999, 12(1): 230-243. DOI:10.1175/1520-0442(1999)012<0230:ECCAID>2.0.CO;2 |
| [55] |
Rangwala I, Miller J R. Climate change in mountains: a review of elevation-dependent warming and its possible causes. Climatic Change, 2012, 114(3/4): 527-547. |
| [56] |
Qin J, Yang K, Liang S L, Guo X F. The altitudinal dependence of recent rapid warming over the Tibetan Plateau. Climatic Change, 2009, 97(1/2): 321-327. |