 2021, Vol. 41
2021, Vol. 41文章信息
- 刘燕山, 谷先坤, 唐晟凯, 李大命, 曾庆飞, 陆建明, 毛志刚, 殷稼雯, 陈辉辉, 沈冬冬, 何浩然, 张彤晴
- LIU Yanshan, GU Xiankun, TANG Shengkai, LI Daming, ZENG Qingfei, LU Jianming, MAO Zhigang, YIN Jiawen, CHEN Huihui, SHEN Dongdong, HE Haoran, ZHANG Tongqing
- 东太湖鱼类群落结构特征及其与环境因子的关系
- Fish community structure and its relationship with main environmental variables in the East Taihu Lake
- 生态学报. 2021, 41(2): 769-780
- Acta Ecologica Sinica. 2021, 41(2): 769-780
- http://dx.doi.org/10.5846/stxb202004130872
-
文章历史
- 收稿日期: 2020-04-13
- 修订日期: 2020-06-30





2. 江苏省内陆水域渔业资源重点实验室, 南京 210017;
3. 中国科学院南京地理与湖泊研究所, 南京 210008;
4. 江苏省太湖渔业管理委员会办公室, 苏州 215104
2. Key Laboratory of Fisheries Resources in Inland Water, Nanjing 210017, China;
3. Nanjing Institute of Georgraphy and Limonology, Chinese Academy of Science, Nanjing 210008, China;
4. Taihu Fishery Management Committee Office of Jiangsu Province, Suzhou 215104, China
转变渔业发展方式, 由经济渔业向生态渔业发展, 是当前大水面渔业正确的发展方向。东太湖位于太湖东南隅, 是太湖的主要出水通道, 有蓄积、排泄洪水、供水等重要功能, 面积约13125 hm2, 是长江中下游典型的草型浅水湖泊, 其年平均水位1.78 m, 平均水温17.1 ℃, 水体容积约1.2×108 m3, 2018年出水量达31.1×108 m3[1-2]。高密度围网养殖是大水面进行经济渔业发展的有效技术之一, 东太湖湖区围网渔业养殖始于1984年, 随着养殖技术的发展, 规模不断扩大, 至2007年养殖面积达到东太湖水体面积的85.30%[3]。大规模的围网养殖不仅带动了经济发展, 同时也带来了水域生态环境恶化、生物资源破坏等一系列问题[4]。2007年5月的“太湖蓝藻事件”后, 东太湖围网面积被缩减至3000 hm2 [5], 但仍未从根本上改善湖泊生态环境。为贯彻落实国务院和江苏省人民政府对太湖水污染防治的重大决策, 按照中央环保督查整改要求, 2018年4月, 政府决定全面拆除东太湖围网, 至2019年4月初, 围网基本全部拆除。因此, 为了向生态渔业发展, 急需了解围网拆除后东太湖的渔业资源状态, 以及渔业资源与环境因子的关系。
生态系统受损后, 其生物多样性将会受到严重干扰[6-7], 鱼类可以作为监测的指示生物, 通过鱼类群落来评价生态系统的生物完整性[8-11]。国内有关鱼类群落与环境因子的关系研究多集中在近海与河口[12-17], 内陆江河较少[18-19], 而内陆浅水型湖泊更是鲜有报道。2019年4月、7月和10月对围网拆除后第一年内的东太湖鱼类群落和主要环境因子进行了监测, 了解东太湖的鱼类群落结构特征, 分析其与主要环境因子的关系, 以便为今后东太湖渔业资源的恢复、保护和管理等提供理论基础和科学依据。
1 材料与方法 1.1 调查站位设置与调查方法2019年4月、7月和10月, 在东太湖的近太浦河口、曾经的主要养殖区、出湖口设置如图 1所示的4个站位进行渔获物和水样的采集。在曾经的养殖区(S1和S2)有一定量留存的水草分布, 出湖口为敞水区(S3)几乎没有水草分布, 近太浦河口(S4)零星分布, 监测站位的平均水深1.53 m, 溶解氧7.02 mg/L, pH为7.83, 透明度42 cm。每个站位均设置一条多目刺网(以下简称刺网)和三条地笼(一个网笼单元)。刺网规格:全长300 m, 分5种网目, 其中有2 cm、3 cm、4 cm、7 cm、10.5 cm网目分别60 m。地笼网规格:中间部位为60个30 cm × 35cm的钢筋框, 两端各有4个直径为26.5 cm的钢筋圈, 钢筋框间距为30 cm, 钢筋圈间距20 cm, 进鱼口纵深22 cm, 网目规格9 mm, 全长20 m。刺网和地笼均放置12 h(约为19:00至次日7:00)。对所采集的鱼类进行鉴定分类、称量和计数。鱼类的学名、分类地位及生态类型等以《太湖鱼类志》[20]、《江苏鱼类志》[21]为主要依据。

|
| 图 1 东太湖鱼类调查站位图 Fig. 1 Sampling stations for the survey of the fishes in the East Taihu Lake 图中S1, S2, S3, S4代表调查站位 |
采集渔获物的同时, 利用哈希HQ40D多参数水质检测仪监测水温(Tem)、溶解氧(DO)、pH等, SM-5(5A)手持式测深仪测量水深(Dep), 萨克斯盘测水体的透明度(SD), 测量总磷(TP)、总氮(TN)、氨氮(NH4-N)、硝态氮(NO3-N)、磷酸盐(PO4-P)、化学需氧量(COD)、和叶绿素a(Chla)等环境因子, 样品的采集、保存及分析参照《内陆水域渔业自然资源调查手册》[22]进行。
1.2 数据整理与分析将每条刺网和每个网笼单元的鱼类重量作为一个样本来进行数据分析, 分析前对原始数据进行标准化处理, 采用Shannon-Wiener多样性指数(H)[23]、Margalef种类丰富度指数(d)[24]、Pielou均匀度指数(J)[25]以及Pinkas相对重要性指数(IRI)[26]对鱼类数据进行分析。各指数的计算公式:(1)多样性指数: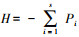
用Primer 5.2软件, 以各月份不同监测站位鱼类的生物量为原始矩阵, 进行四次方根转换以后, 利用等级聚类(Cluster)[28]和非度量多维尺度排序(NMDS)[29]分析各季节不同断面鱼类的组成特征。利用ABC(abundance-biomass comparison)曲线和W统计(W-statistic)描述群落稳定性以及生物量/丰度的优势地位[30-31]。采用单因子相似性分析(ANOSIM)研究不同月份组之间的群落种类组成的差异显著性[29], SIMPER相似性百分比分析鱼类群落的不同月份组内相似性和组间差异性的贡献率, 造成各月份组内群落结构相似的典型种和造成组间群落结构差异的分歧种, 以及各鱼种对组内相似性和组间相异性的贡献率[32]。
采用CANOCO 4.5软件对各月份的鱼类重量组成和环境因子进行梯度分析, 首先将各样方的渔获量组成数据取对数后进行去趋势对应分析(DCA), 根据分析结果中各排序轴长度决定排序的模型(如排序轴长度大于4, 选择单峰模型, 反之, 则选择线性模型), 由于本研究排序轴长度均小于3, 选择线性模型的主成分分析(PCA)和冗余分析(RDA)两种排序方法[13]。在RDA分析过程中, 运用蒙特卡罗检验分析评价环境因子对鱼类群落变化影响的显著程度。
使用Excel 2010、SPSS 19.0软件对相关数据进行统计处理, 不同监测月份和站位间的差异利用单因素方差分析, 显著性水平为α=0.05。使用PRIMER 5.2、CANOCO 4.5和Adobe Illustrator CS6软件绘图, 站位布设图在ArcGIS 10.2软件中完成。
2 结果和分析 2.1 鱼类群落种类组成及优势种调查期间, 共监测到东太湖鱼类4目10科31属39种, 鲤形目鱼类27种, 占69.23%, 鲈形目8种, 占20.51%, 鲇形目3种, 占7.69%, 鲱形目1种, 占2.56%;鲤科鱼类有26种, 占据66.67%, 优势明显, 其它各科种类数均不超过3种。鱼类的生态类型:按洄游方式, 定居性鱼类分别占总数量和总质量的52.64%和48.79%, 河湖洄游性鱼类分别占31.16%和28.17%, 江海洄游性鱼类分别占16.2%和23.04%;按栖息水层, 中上层鱼类分别占总数量和总质量的59.19%和62.89%, 中下层鱼类分别占28.79%和27.04%, 底层鱼类分别占12.02%和10.07%;按食性, 肉食性鱼类分别占总数量和总质量的32.87%和38.30%, 杂食性分别占32.87%和28.38%, 植食性分别占19.24%和18.82%, 浮游生物食性分别占13.40%和11.49%, 碎屑食性分别占1.62%和3.01%。鱼类种类及生态类型详见表 1。
| 图中代号 Symbol in the figures |
种类 Species |
生态类型 Ecological types |
| Coi | 鲱形目Clupeiformes | |
| 鳀科Engraulidae | ||
| 刀鲚Coilia nasus | RS, U, C | |
| 鲤形目Cypriniformes | ||
| 鲤科Cyprinidae | ||
| Cte | 草鱼Ctenopharyngodon idellus | RL, L, H |
| Ari | 鳙Aristichthys nobilis | RL, U, P |
| Cyp | 鲤Cyprinus carpio | SE, D, O |
| Car | 鲫Carassius auratus | SE, L, O |
| Hel |  鱼Hemiculter leucisculus 鱼Hemiculter leucisculus |
SE, U, O |
| Heb | 贝氏 Hemiculter bleekeri |
SE, U, O |
| Cua | 翘嘴鲌Culter alburnus | SE, U, C |
| Cum | 蒙古鲌Culter mongolicus | SE, U, C |
| Cud | 达氏鲌Culter dabryi | SE, U, C |
| Cue | 红鳍原鲌Cultrichthys erythropterus | SE, U, C |
| Pap | 鳊Parabramis pekinens | RL, L, H |
| Tox | 似鱎Toxabramis swinhonis | SE, U, O |
| Meg | 团头鲂Megalobrama amblycephala | RL, L, O |
| Pss | 似鳊Pseudobrama simoni | SE, L, H |
| Xea | 银鲴Xenocypris argentea | SE, D, H |
| Xem | 细鳞鲴Xenocypris microlepis | SE, L, H |
| Acm | 大鳍鱊Acheilognathus macropterus | SE, U, De |
| Acc | 兴凯鱊Acheilognathus chankaensis | SE, U, De |
| Rhs | 中华鰟鲏Rhodeus sinensis | SE, U, De |
| Rho | 高体鳑鮍Rhodeus ocellatus | SE, U, De |
| Pse | 麦穗鱼Pseudorasbora parva | SE, D, O |
| Abb | 棒花鱼Abbottina rivularis | SE, D, O |
| Hem | 花 Hemibarbus maculatus Hemibarbus maculatus |
SE, L, C |
| Sau | 蛇鮈Saurogobio dabryi | SE, L, O |
| Sar | 华鳈Sarcocheilichthys sinensis | SE, L, C |
| Par | 似刺鳊鮈Paracanthobrama guichenoti | SE, D, C |
| 鳅科Cobitidae | ||
| Mis | 泥鳅Misgurnus anguillicaudatus | SE, D, O |
| 鲇形目Siluriformes | ||
| 鲿科Bagridae | ||
| Pef | 黄颡鱼Pelteobagrus fulvidraco | SE, D, C |
| Pee | 长须黄颡鱼Pelteobagrus eupogon | SE, D, C |
| Pen | 光泽黄颡鱼Pelteobagrus nitidus | SE, D, C |
| 鲈形目Perciformes | ||
| 虾虎鱼科Gobiidae | ||
| Rhi | 子陵吻虾虎鱼Rhinogobius giurinus | RS, D, C |
| Tae | 须鳗虾虎鱼Taenioides cirratus | RS, D, C |
| 塘鳢鱼科Eleotridae | ||
| Odo | 河川沙塘鳢Odontobutis potamophila | SE, D, C |
| Mic | 小黄黝鱼Micropercops swinhonis | SE, D, O |
| 斗鱼科Belontiidae | ||
| Mac | 圆尾斗鱼Macropodus chinensis | SE, L, C |
| 月鳢科Channidae | ||
| Cha | 乌鳢Channa argus | SE, D, C |
| 刺鳅科Mastacembelidae | ||
| Mas | 中华刺鳅Mastacembelus sinensis | SE, D, C |
| 鮨科Serranidae | ||
| Sin | 鳜Siniperca chuatsi | SE, L, C |
| RS:江海洄游性River-sea migratory;RL:河湖涸游性River-lake migratory;SE:定居性Sedentary fish;U:中上层Upper;L:中下层Lower;D:底层Demersal;P:浮游生物食性Plankivore;De:碎屑食性Detritivore;C:肉食性Carnivore;H:植食性Herbivore;O:杂食性Omnivore | ||
表 2给出了各频次调查中IRI≥100的鱼类种类分布, 主要种鱼类有16种。整个监测期间, 东太湖鱼类优势种为鲫(Carassius auratus)、大鳍鱊(Acheilognathus macropterus)和(Hemiculter leucisculus)。4月、7月和10月, 优势种分别为鲫和, 大鳍鱊、鲫、刀鲚(Coilia nasus)、麦穗鱼(Pseudorasbora parva)和红鳍原鲌(Cultrichthys erythropterus), 以及刀鲚和红鳍原鲌。根据渔获量组成, 鳙(Aristichthys nobilis)的生物量占比最多(23.02%), 其次为鲫(12.25%)、(11.49%)、刀鲚(7.65%)、红鳍原鲌(7.07%), 其它种类共占38.51%;按数量百分比, 刀鲚最多(16.10%), 其次为(13.73%)、大鳍鱊(13.40%)、鲫(9.93%)、麦穗鱼(8.88%)、红鳍原鲌(5.89%), 其余共占32.07%。
| 种类Species | 2019-04 | 2019-07 | 2019-10 | 总和Sum |
| 刀鲚Coilia nasus | 79.14 | 1481.92 | 4385.89 | 989.65 |
| 鳙Aristichthys nobilis | 949.87 | 778.27 | 739.71 | 783.76 |
| 鲤Cyprinus carpio | 55.18 | 39.26 | 198.62 | 68.21 |
| 鲫Carassius auratus | 1705.97 | 1878.83 | 443.32 | 1570.76 |
| 鱼Hemiculter leucisculus |
2747.62 | 727.88 | 136.74 | 1261.19 |
| 贝氏 Hemiculter bleekeri |
855.55 | - | - | 154.28 |
| 红鳍原鲌Cultrichthys erythropterus | 570.24 | 1171.74 | 2100.91 | 972.28 |
| 似鳊Pseudobrama simoni | 483.58 | 13.99 | 162.72 | 208.25 |
| 银鲴Xenocypris argentea | 149.39 | - | - | 27.49 |
| 细鳞鲴Xenocypris microlepis | 145.56 | - | - | 26.93 |
| 大鳍鱊Acheilognathus macropterus | 662.97 | 2901.55 | 874.06 | 1267.05 |
| 兴凯鱊Acheilognathus chankaensis | 629.01 | 149.10 | 65.40 | 277.67 |
| 麦穗鱼Pseudorasbora parva | 217.27 | 1296.72 | 29.94 | 470.67 |
| 花Hemibarbus maculatus |
226.12 | 37.01 | 34.18 | 106.74 |
| 黄颡鱼Pelteobagrus fulvidraco | 574.94 | 475.24 | - | 339.65 |
| 长须黄颡鱼Pelteobagrus eupogon | 893.51 | 9.35 | 38.29 | 280.06 |
各监测站位在4月、7月、10月的鱼类种类数、渔获量、丰富度指数、均匀度指数和多样性指数结果见表 3。单因素方差分析表明, 监测时间内不同站位间鱼类的种类数、渔获量、丰富度指数、均匀度指数和多样性指数差异均不显著(P>0.05);除均匀度指数外, 3个监测月份间鱼类的种类数、渔获量、丰富度指数和多样性指数均差异显著(P<0.05), 为4月>7月>10月。
| 站位 Station |
种类数 Species number |
重量/g Wet weight |
丰富度 Species richness |
均匀度 Pielou's evenness |
多样性指数 Shannon-Wiener index |
||||||||||
| 2019-04 | 2019-07 | 2019-10 | 2019-04 | 2019-07 | 2019-10 | 2019-04 | 2019-07 | 2019-10 | 2019-04 | 2019-07 | 2019-10 | 2019-04 | 2019-07 | 2019-10 | |
| S1 | 22 | 12 | 5 | 14511.9 | 3564.2 | 563.6 | 3.641 | 2.147 | 1.294 | 0.848 | 0.775 | 0.750 | 3.784 | 2.777 | 1.741 |
| S2 | 17 | 10 | 7 | 9409.2 | 3432.9 | 3158.7 | 2.959 | 1.617 | 1.403 | 0.724 | 0.679 | 0.698 | 2.959 | 2.254 | 1.960 |
| S3 | 20 | 15 | 11 | 15267.9 | 7575.5 | 4212.9 | 3.363 | 2.919 | 2.078 | 0.667 | 0.590 | 0.635 | 2.883 | 2.306 | 2.197 |
| S4 | 17 | 13 | 9 | 6822.2 | 5002.2 | 3478.4 | 2.904 | 2.219 | 2.127 | 0.832 | 0.546 | 0.833 | 3.403 | 2.021 | 2.641 |
| 均值 Mean | 19 | 12.5 | 8 | 11502.8 | 4893.7 | 2853.4 | 3.217 | 2.226 | 1.726 | 0.768 | 0.647 | 0.729 | 3.257 | 2.340 | 2.135 |
对鱼类群落进行等级聚类分析(Cluster)和非度量多维尺度排序(NMDS)分析(图 2)。Cluster分析结果表明, 除10月份的1号点和3号点外, 各样方间的相似度在46.75%以上。相似性最高的两个样方为4月份的1号点和3号点, 相似度达到76.19%。NMDS分析结果与聚类分析相似度在50%以上的结果一致。Stress系数用来检验NMDS结果的可信度[29]:当Stress<0.1时结果具有较好的解释意义, 0.1≤Stress<0.2时结果具有一定的解释意义, Stress≥0.2时结果不具解释意义。本次检验的Stress值为0.15, 具有一定的解释意义。

|
| 图 2 东太湖鱼类群落结构聚类和NMDS排序分析 Fig. 2 The dendrogram and the NMDS for the similarity matrix of fish community of the East Taihu Lake S1-04:4月S1站位S1 in April;S2-04:4月S2站位S2 in April;S3-04:4月S3站位S3 in April;S4-04:4月S4站位S4 in April;S1-07:7月S1站位S1 in July;S2-07:7月S2站位S2 in July;S3-07:7月S3站位S3 in July;S4-07:7月S4站位S4 in July;S1-10:10月S1站位S1 in October;S2-10:10月S2站位S2 in October;S3-10:10月S3站位S3 in October;S4-10:10月S4站位S4 in October |
单因子相似性分析(ANOSIM)表明, 监测月份间的鱼类群落物种组成, 4月和10月间差异显著(P=0.029), 其它月份间差异不显著(P>0.05)。相似性百分比(SIMPER)表明, 4月各站位间鱼类的平均相似性为65.41%, 特征种为兴凯鱊(Acheilognathus chankaensis)、长须黄颡鱼(Pelteobagrus eupogon)、贝氏(Hemiculter bleekeri)、、红鳍原鲌、黄颡鱼(Pelteobagrus fulvidraco)、鲫和麦穗鱼, 累计贡献64.64%;7月各站位间鱼类的平均相似性为58.35%, 特征种为、大鳍鱊、红鳍原鲌、黄颡鱼、鲫、麦穗鱼和兴凯鱊, 累计贡献89.28%, 10月各站位间鱼类的平均相似性为41.59%, 特征种为刀鲚、大鳍鱊、鲫、红鳍原鲌、似鳊(Pseudobrama simoni)、长须黄颡鱼和麦穗鱼, 累计贡献91.54%。4月和10月的平均相异性为59.41%, 分歧种为贝氏、黄颡鱼、兴凯鱊、、细鳞鲴(Xenocypris microlepis)、银鲴(Xenocypris microlepis)和河川沙塘鳢(Odontobutis potamophila), 累计贡献35.87%。
ABC法可以通过比较生物量和丰度曲线来分析不同干扰情况下鱼类群落的反应[30-31]:在未受干扰的状态下, 生物量优势度曲线在上方;中等干扰状态时, 两条曲线将相交;当生物量的优势度曲线在下方, 表明群落处于严重干扰。鱼类ABC曲线显示(图 3), 东太湖鱼类生物量和丰度曲线有不同程度的交叉重叠, W值为-0.006, 故鱼类群落处于中度干扰状态。

|
| 图 3 东太湖鱼类丰度/生物量曲线 Fig. 3 Abundance-biomass comparison curves of the fish in the East Taihu Lake |
DCA排序结果如表 4所示, 第一轴最长为2.39, 解释的物种变化率为21.3%, 第一轴与环境因子的相关性较高(96.5%), 四个排序轴的长度均小于3, 因此东太湖鱼类群落参数对环境因子的响应呈线性关系, 适用线性模型[33]。
| 排序轴编号Axes | 1 | 2 | 3 | 4 |
| 特征值Eigenvalues | 0.285 | 0.127 | 0.049 | 0.012 |
| 梯度长度Lengths of gradient | 2.394 | 1.566 | 1.655 | 1.312 |
| 物种和环境因子排序轴相关系数Species-environment correlations | 0.965 | 0.997 | 0.764 | 0.999 |
| 变量累计百分比Cumulative percentage variance/% | ||||
| 物种数据累计百分比Cumulative percentage variance of species data/% | 21.3 | 30.7 | 34.4 | 35.3 |
| 物种和环境关系累计百分比 Cumulative percentage variance of species-environment relation/% |
20.8 | 34.3 | 0.0 | 0.0 |
对群落进行PCA分析(表 5), 前两轴为主成分轴, 解释了53.3%的物种变化, 利用10个环境因子的变化能解释94.1%的鱼类群落总变化。根据PCA分析结果, 做物种—环境因子的排序图 4, 可以看出总磷、溶解氧与第一轴正相关性较强, 溶解氧、pH与第二排序轴负相关性较强。
| 排序轴编号Axes | 1 | 2 | 3 | 4 |
| 特征值Eigenvalues | 0.320 | 0.213 | 0.108 | 0.075 |
| 物种和环境因子排序轴相关系数Species-environment correlations | 0.996 | 0.949 | 0.987 | 0.956 |
| 变量累计百分比Cumulative percentage variance/% | ||||
| 物种数据累计百分比Cumulative percentage variance of species data/% | 32.0 | 53.3 | 64.1 | 71.6 |
| 物种和环境关系累计百分比 Cumulative percentage variance of species-environment relation/% |
33.8 | 54.1 | 65.3 | 72.6 |

|
| 图 4 物种与环境因子的PCA双序图 Fig. 4 PCA graph of species and environment factors Tem(水温), DO(溶解氧), Dep(水深), SD(透明度), TP(总磷), TN(总氮), 磷酸盐(PO4-P)、COD(化学需氧量), Chla(叶绿素a) |
针对PCA分析结果, 利用RDA分析来研究总磷和其它环境因子与主要鱼类物种的关系(图 5), 分析表明总磷对主要种类的解释量为21.0%, 是显著影响因子(蒙特卡罗检验P=0.036), 红鳍原鲌、鳙、花(Hemibarbus maculatus)等与总磷相关性较大。将总磷作为协变量, 而将其它环境因子作为环境变量引入RDA分析(图 6), 结果表明, 除总磷以外的环境因子对主要鱼类群落的解释率达到74.2%, pH与第一轴负相关性高, 对长须黄颡鱼、黄颡鱼、兴凯鱊、贝氏、鲫鱼、刀鲚的影响较大;溶解氧与第二轴正相关性较高, 对似鳊、鲤(Cyprinus carpio)、红鳍原鲌、大鳍鱊的影响较大。蒙特卡罗检验表明, 除总磷外其它环境因子对主要鱼类群落的解释不显著(P=0.24)。

|
| 图 5 基于RDA分析的总磷对主要鱼类分布影响图 Fig. 5 TP-main species biplot based on RDA analysis |

|
| 图 6 除总磷外的其它环境因子与鱼类群落关系图 Fig. 6 Other environment factors-main species biplot based on RDA analysis with TP as covariable |
根据记载, 历史上太湖共有鱼类107种[20], 朱松泉等[34]在2002—2006年的调查中共监测到60种, 毛志刚等[35]于2009—2010年捕获到鱼类50种。然而, 针对东太湖鱼类的调查, 未见报道。本文采集到东太湖鱼类39种, 其中鲤科鱼类种类数占比66.67%, 均为太湖常见鱼类物种。鱼类种类数远低于20纪90年代的60多种[36], 仅为太湖历史鱼类总种类的36.44%, 且低于2013—2014年太湖流域河流的46种[35];鲤科种类占比与太湖鱼类相当(64%)[35], 高于太湖流域河流的56.52%[37]。定居性鱼类为东太湖的主要鱼类, 而受建造闸坝等的影响, 洄游性鱼类非常少, 未发现尖头鲌(Culter oxycephaloides)、鱤(Elopichthys bambusa)等河湖洄游性鱼类, 以及日本鳗鲡(Anguilla japonica)等江海洄游性鱼类, 而仅有的江海洄游性鱼类刀鲚、子陵吻虾虎鱼(Rhinogobius giurinus)和须鳗虾虎鱼(Taenioides cirratus)也适应了这里的生态环境并能够繁殖, 演变成了次生的定居性鱼类[20, 27, 34-35]。从食性上看, 主要为杂食性和肉食性鱼类, 可能是因为杂食性鱼类食物种类多样, 生态位较宽, 适应环境能力较强[38];而肉食性鱼类中主要是刀鲚、红鳍原鲌、黄颡鱼等摄食小鱼、小虾、小型软体动物、水生昆虫等的种类[20-21], 这也与东太湖的资源现状相适应。
鱼类小型化已成为内陆江河、水库、湖泊等的一种常态[27, 35, 37-41], 按照初次性成熟小于2龄、最大体长小于24 cm的鱼类划为小型鱼类的标准[39], 东太湖主要种鱼类中除鳙、鲤外, 均为小型鱼类, 表明东太湖鱼类呈现小型化。这可能是由于湖区长期大范围的高密度围网养殖等人类活动挤压了鱼类在自然水域的生存空间, 以及受水质污染、湖泊沼泽化、蓝藻水华等水环境问题的影响[1, 42]。
4月、7月和10月东太湖的平均多样性指数分别为3.257, 2.340和2.135, 在一般指数范围(1.5—3.5)之内[43], 说明鱼类群落多样性处于一般水平。4月鱼类的种类数、渔获量、丰富度指数和多样性指数均较大, 这是由于4月采样正值东太湖围网基本拆除之后, 鱼类生存空间增大, 活动增强, 被捕获几率增加, 故捕获种类和渔获量也较多, 至7月和10月逐渐趋于平稳。此外, 捕捞渔具的使用也会相应的影响监测鱼类的多样性指数[27, 35], 主动型渔具(如拖网、高踏网等)多捕捞游泳能力较弱的鱼类(如刀鲚), 而被动型渔具(鱼簖、地笼、刺网等)更容易受水流、天气等外界因素的影响, 而本研究所用渔具均属被动型网具。
相似性分析显示, 4月和10月的鱼类群落物种组成有显著差异, 主要分歧种为贝氏、黄颡鱼、兴凯鱊等, 而4月和7月, 7月和10月差异并不显著。聚类分析也显示, 除10月份的1号点和3号点外, 各样方间的物种组成相似度较高。表明, 围网拆除后东太湖的鱼类群落结构正在随着时间的推移发生变化。这可能是由季节差异造成的, 也与生态系统的自我恢复有很大关系。
3.2 鱼类群落结构与环境因子关系鱼类群落的空间格局除受其本身生活习性影响外, 还受环境因子空间异质性的影响[18], 主要环境影响因子根据水域环境特点不同而有差异。水温、水深、盐度等是影响近海与河口及其邻近水域鱼类群落结构的主要环境因子[12-17], 河宽、温度、海拔、pH、溶解氧、距河口距离、据水坝距离等则是影响河流鱼类群落结构的主要环境因子[18-19, 44], 水深和透明度与通河湖泊鱼类群落结构变化密切相关[45]。程林在研究长江流域湖泊鱼类群落与主要环境因子关系及其预测模型中得出, 总磷、总氮、叶绿素a、透明度和水深是影响长江流域湖泊鱼类群落的主要环境因素[46]。本文通过RDA分析及蒙特卡罗检验, 得出总磷是目前影响东太湖鱼类群落结构变化的显著因子, 与红鳍原鲌、鳙、花等相关性较强。这是因为这些鱼类的耐受能力较强, 而耐受鱼类的增多同样也是水域环境恶化的标志[46-47]。此外, 由于东太湖受水环境污染的影响较大, 而氮、磷负荷是影响水环境的重要因素, 又有研究表明东太湖水质污染主要表现为富营养化, 限制因子为总磷, 当水体营养处于超饱和状态时, 氮负荷的影响力下降, 而磷的负荷影响力上升, 故磷污染贡献远高于氮[42, 48-49]。
去除总磷, 分析其它环境因子对鱼类群落的影响, 表明pH和溶解氧对鱼类群落的第一轴和第二轴影响最大。监测期间东太湖水体的pH值在7.06—8.20之间, 呈中性至弱碱性, 刀鲚与东太湖pH的变化呈正相关, 而长须黄颡鱼、黄颡鱼、兴凯鱊等呈负相关。有研究表明水体的pH值变化会对鱼类的摄食、生长等造成一定的影响, 也会影响鱼类的感官、代谢、呼吸等生理过程[50-51]。溶解氧与东太湖似鳊、鲤、红鳍原鲌等相关性较强。受季节、围网拆除等因素影响, 湖区4月水草植被较少, 7月较繁茂, 10月一般, 然而7月水温较高(30.8℃)、水中含有大量蓝藻, 而4月(21.9℃)和10月(20.5℃)水温较适宜, 再加上围网养殖残留在沉积物中的有机物降解消耗等, 导致水中溶解氧的含量在7月较低(5.2 mg/L)、4月最高(8.6 mg/L)。因此, 溶解氧也是季节、水温、底质沉积物等因素综合影响的表现。
4 结论与建议本文对围网拆除后东太湖的鱼类群落结构及其与主要环境因子的关系进行了研究, 共监测到鱼类4目10科31属39种, 按不同生态类型主要为定居性鱼类、中上层鱼类以及杂食性和肉食性鱼类, 优势种为鲫、大鳍鱊、等, 鱼类个体小型化明显, 鱼类多样性水平一般, 鱼类群落结构处于中度干扰状态, 目前总磷、pH、溶解氧等是影响东太湖鱼类群落的主要环境因子。建议通过恢复水生植被、合理放流净水物种(如河蚬、鲢、鳙等)、控制陆源污染物等方式调节水域环境质量, 以加快东太湖生态系统恢复。
| [1] |
吴庆龙. 东太湖养殖渔业可持续发展的思考. 湖泊科学, 2001, 13(4): 337-343. DOI:10.3321/j.issn:1003-5427.2001.04.007 |
| [2] |
殷鹏, 张建华, 华萍. 改善东太湖水环境的水工程措施研究. 江苏水利, 2019(7): 1-4, 8-8. |
| [3] |
何俊, 谷孝鸿, 刘国锋. 东太湖网围养蟹效应及养殖模式优化. 湖泊科学, 2009, 21(4): 523-529. DOI:10.3321/j.issn:1003-5427.2009.04.011 |
| [4] |
范亚民, 李海宇, 何华春, 周盛兵. 近30年来太湖围湖利用及东太湖网围养殖动态变化研究. 长江流域资源与环境, 2012, 21(S2): 121-126. |
| [5] |
生楠.东太湖围网养蟹经济效益研究[D].上海: 上海海洋大学, 2018.
|
| [6] |
Allan J, Johnson L. Catchment-scale analysis of aquatic ecosystems. Freshwater Biology, 1997, 37(1): 107-111. DOI:10.1046/j.1365-2427.1997.00155.x |
| [7] |
Grevilliot F, Krebs L, Muller S. Comparative importance and interference of hydrological conditions and soil nutrient gradients in floristic biodiversity in flood meadows. Biodiversity & Conservation, 1998, 7(11): 1495-1520. |
| [8] |
Karr J R. Assessment of biotic integrity using fish communities. Fisheries, 1981, 6(6): 21-27. DOI:10.1577/1548-8446(1981)006<0021:AOBIUF>2.0.CO;2 |
| [9] |
Karr J R, Fausch K D, Angermeier P L, Yant I J, Schlosser I J. Assessing Biological Integrity in Running Waters:A Method and Its Rationale. Champaign:Illinois Natural History Survey, 1986, 1-28. |
| [10] |
Joy M K, Death R G. Predictive modelling of freshwater fish as a biomonitoring tool in New Zealand. Freshwater Biology, 2002, 47(11): 2261-2275. DOI:10.1046/j.1365-2427.2002.00954.x |
| [11] |
Barbour M T, Gerritsen J, Snyder B D, Stribling J B. Rapid Bioassessment Protocols For Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates, and Fish. 2nd ed. Washington, DC: U.S. Environmental Protection Agency, Office of Water, 1999.
|
| [12] |
汪振华, 章守宇, 王凯. 三横山鱼礁生境鱼类和大型无脊椎动物群落特征. 生态学报, 2010, 30(8): 2026-2035. |
| [13] |
吴忠鑫, 张磊, 张秀梅, 张沛东, 李文涛. 荣成俚岛人工鱼礁区游泳动物群落特征及其与主要环境因子的关系. 生态学报, 2012, 32(21): 6737-6746. |
| [14] |
王小林, 徐宾铎, 纪毓鹏, 任一平. 海州湾及邻近海域冬季鱼类群落结构及其与环境因子的关系. 应用生态学报, 2013, 24(6): 1707-1714. |
| [15] |
许永久, 俞存根, 张平, 邓小艳, 张喆, 沈海琪. 杭州湾-舟山近海春季游泳动物群落结构及与环境因子的关系. 水产学报, 2019, 43(3): 605-617. |
| [16] |
翟璐, 徐宾铎, 纪毓鹏, 任一平. 黄河口及其邻近水域夏季鱼类群落空间格局及其与环境因子的关系. 应用生态学报, 2015, 26(9): 2852-2858. |
| [17] |
何思璇, 何斌源. 防城河口湾鱼类群落结构及其与环境因子关系研究. 热带海洋学报, 2019, 38(5): 86-97. |
| [18] |
李捷, 李新辉, 贾晓平, 谭细畅, 王超, 李跃飞, 邵晓风. 连江鱼类群落多样性及其与环境因子的关系. 生态学报, 2012, 32(18): 5795-5805. |
| [19] |
张迎秋, 黄稻田, 李新辉, 刘乾甫, 李捷, 李跃飞, 杨计平, 朱书礼. 西江鱼类群落结构和环境影响分析. 南方水产科学, 2020, 16(1): 42-52. |
| [20] |
倪勇, 朱成德. 太湖鱼类志. 上海: 上海科学技术出版社, 2005.
|
| [21] |
倪勇, 伍汉霖. 江苏鱼类志. 北京: 中国农业出版社, 2006.
|
| [22] |
张觉民, 何志辉. 内陆水域渔业自然资源调查手册. 北京: 农业出版社, 1991.
|
| [23] |
Ludwig J A, Reynolds J F. Statistical Ecology:A Primer on Methods and Computing. New York: Wiley, 1988.
|
| [24] |
Margalef R. Perspectives in Ecological Theory. Chicago: University of Chicago Press, 1968: 1-111.
|
| [25] |
Pielou E C. Species-diversity and pattern-diversity in the study of ecological succession. Journal of Theoretical Biology, 1966, 10(2): 370-383. DOI:10.1016/0022-5193(66)90133-0 |
| [26] |
Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, Bluefin tuna, and bonito in California waters. Fish Bulletin, 1971, 152: 1-105. |
| [27] |
刘燕山, 唐晟凯, 李大命, 谷先坤, 朱滨清, 毛国庆, 张彤晴. 淮河江苏段鱼类群落结构特征. 中国水产科学, 2020, 27(2): 224-235. |
| [28] |
Bray J R, Curtis J T. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 1957, 27(4): 325-349. DOI:10.2307/1942268 |
| [29] |
Clarke K R, Gorley R N, Somerfield P J, Warwick R M. Change in Marine Communities:An Approach to Statistical Analysis and Interpretation. 3rd ed. Plymouth: PRIMER-E, 2014.
|
| [30] |
Warwick R M. A new method for detecting pollution effects on marine macrobenthic communities. Marine Biology, 1986, 92(4): 557-562. DOI:10.1007/BF00392515 |
| [31] |
Yemane D, Field J G, Leslie R W. Exploring the effects of fishing on fish assemblages using abundance biomass comparison (ABC) curves. ICES Journal of Marine Science, 2005, 62(3): 374-379. DOI:10.1016/j.icesjms.2005.01.009 |
| [32] |
Clarke K R. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 1993, 18(1): 117-143. DOI:10.1111/j.1442-9993.1993.tb00438.x |
| [33] |
Lepš J, Šmilauer P. Multivariate Analysis of Ecological Data Using CANOCO. Cambridge: Cambridge University Press, 2003.
|
| [34] |
朱松泉, 刘正文, 谷孝鸿. 太湖鱼类区系变化和渔获物分析. 湖泊科学, 2007, 19(6): 664-669. DOI:10.3321/j.issn:1003-5427.2007.06.007 |
| [35] |
毛志刚, 谷孝鸿, 曾庆飞, 周露洪, 王小林, 吴林坤, 曹萍, 孙明波. 太湖鱼类群落结构及多样性. 生态学杂志, 2011, 30(12): 2836-2842. |
| [36] |
吴庆龙, 陈开宁, 周万平. 东太湖生物资源利用与生态环境保护. 自然资源, 1996(3): 43-47. |
| [37] |
李其芳, 严云志, 储玲, 朱仁, 高俊峰, 高永年. 太湖流域河流鱼类群落的时空分布. 湖泊科学, 2016, 28(6): 1371-1380. |
| [38] |
王银平, 匡箴, 蔺丹清, 李佩杰, 杨彦平, 刘凯. 长江安庆新洲水域鱼类群落结构及多样性. 生态学报, 2020, 40(7): 2417-2426. |
| [39] |
张晓可, 于道平, 王慧丽, 万安, 蒋泽球, 陈寿文. 长江安庆段江豚主要栖息地鱼类群落结构. 生态学报, 2016, 36(7): 1832-1839. |
| [40] |
向鹏, 刘良国, 王冬, 曾平文, 邓玲玲, 杨春英, 杨品红. 湖南沅水五强溪水库鱼类资源现状及其历史变化. 湖泊科学, 2016, 28(2): 379-386. |
| [41] |
陈校辉, 边文冀, 赵钦, 严维辉, 彭刚. 长江江苏段鱼类种类组成和优势种研究. 长江流域资源与环境, 2007, 16(5): 571-577. DOI:10.3969/j.issn.1004-8227.2007.05.005 |
| [42] |
谷孝鸿, 王晓蓉, 胡维平. 东太湖渔业发展对水环境的影响及其生态对策. 上海环境科学, 2003, 22(10): 702-704. |
| [43] |
Magurran A E. Ecological Diversity and Its Measurement. New Jersey: Princeton University Press, 1988: 7-45.
|
| [44] |
Kadye W T, Magadza C H D, Moyo N A G, Kativu S. Stream fish assemblages in relation to environmental factors on a montane plateau (Nyika Plateau, Malawi). Environmental Biology of Fishes, 2008, 83(4): 417-428. DOI:10.1007/s10641-008-9364-4 |
| [45] |
Tejerina-Garro F L, Fortin R, Rodríguez M A. Fish community structure in relation to environmental variation in floodplain lakes of the Araguaia River, Amazon Basin. Environmental Biology of Fishes, 1998, 51(4): 399-410. DOI:10.1023/A:1007401714671 |
| [46] |
程琳.长江流域湖泊鱼类群落与主要环境因子关系及其预测模型[D].北京: 中国科学院大学, 2011.
|
| [47] |
Schleiger S L. Use of an index of biotic integrity to detect effects of land uses on stream fish communities in west-central Georgia. Transactions of the American Fisheries Society, 2000, 129(5): 1118-1133. DOI:10.1577/1548-8659(2000)129<1118:UOAIOB>2.0.CO;2 |
| [48] |
展永兴, 吴心艺. 东太湖水体中嗅味物质分布及网围养殖对其影响. 江苏水利, 2018(9): 1-5. |
| [49] |
赖力, 黄贤金. 东太湖网围养殖生态环境影响的磷响应模型分析及生态环境成本测算//太湖高级论坛交流文集. 上海:中国水利学会, 2004, 316-323. |
| [50] |
李明德. 鱼类生态学. 天津:天津科技翻译出版公司, 1990, 3l-32. |
| [51] |
周洪琪, 彼得·约翰逊. pH对鱼类化学感觉的影响. 水产学报, 1988, 12(2): 169-172. |