 2021, Vol. 41
2021, Vol. 41文章信息
- 郭文婷, 王国华, 缑倩倩
- GUO Wenting, WANG Guohua, GOU Qianqian
- 钠盐胁迫对藜科一年生草本植物生长和生物量分配的影响
- Effects of sodium salt stress on the growth and biomass allocation of Chenopodiaceae annuals
- 生态学报. 2021, 41(16): 6633-6643
- Acta Ecologica Sinica. 2021, 41(16): 6633-6643
- http://dx.doi.org/10.5846/stxb202008142125
-
文章历史
- 收稿日期: 2020-08-14
- 网络出版日期: 2021-05-21

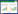




2. 中国科学院西北生态环境资源研究院, 沙漠与沙漠化重点实验室, 兰州 730000;
3. 中国科学院西北生态环境资源研究院临泽内陆河流域研究站, 兰州 730000
2. Key Laboratory of Desert and Desertification, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Science, Lanzhou 730000, China;
3. Linze Inland River Basin Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
土壤盐碱化是全球主要生态环境问题之一, 目前全世界可利用土地面积的60%存在土壤盐碱化, 且呈不断加剧趋势[1]。我国盐碱土分布广泛, 面积大且类型多样, 其中干旱、半干旱地区降水稀少、蒸发强烈, 可溶性盐分随水分运移到土壤表层累积形成大面积盐碱化区域, 深刻影响着干旱区农业发展和绿洲生态稳定[2], 威胁着植物的生长发育。荒漠绿洲过渡带作为干旱区绿洲生态系统的重要组成部分, 对维持绿洲生态稳定具有重要作用[3]。然而近年来受全球变暖、人口增加和人类活动加剧的影响, 荒漠绿洲边缘水分资源恶化, 天然植被衰退[4], 土地荒漠化和盐渍化加剧[5], 我国开始在绿洲边缘大量种植人工固沙植被以恢复绿洲边缘生态环境。随着人工林种植年限的增加, 土壤表层盐分(Na+、Cl-、HCO3-、SO42-)集聚严重[6-7], 灌木和多年生草本植物入侵稀少或难以扩张, 而一年生草本植物却可以大量入侵并成为草本层的优势植物类群。
一年生植物是干旱荒漠植物区系的重要组成部分, 具有重要的生态功能。干旱荒漠生态系统干旱少雨, 温度、湿度和降雨时空变异性大, 土壤基质空间异质性极强, 作为荒漠生态系统恒有层片, 一年生植物在自然选择和长期进化过程中形成了完善的生存策略[1], 并以特有的生物学特性使其成为许多植物生态学问题研究的最佳试验材料[8]。近年来许多学者研究了盐碱胁迫对一年生植物种子萌发[9]、光合作用[8;10]和生理变化[11-12]等的影响, 证明了盐碱胁迫对植物有很强的致害作用。在盐渍化土壤环境下, 盐碱胁迫是影响植物生长发育的关键非生物胁迫因子[13]。
藜科一年生草本植物在干旱荒漠区一年生草本植物层中占有绝对优势[14], 是荒漠、半荒漠和盐碱地上重要的建群种和共建种, 分布广泛, 适应性强, 在维持荒漠地区生态平衡等方面有着非常重要的作用[15]。目前关于藜科一年生草本植物的研究多见于群落结构[16]、生理生态特性[17]、种子萌发特性[18]和短期盐胁迫对种子萌发和幼苗生长的影响[1, 9]等。近年来, 随着干旱荒漠地区土地荒漠化和盐渍化的逐渐加剧, 国内外许多学者在藜科植物对盐碱胁迫的响应方面展开了大量研究, 进行了长期观测, 发现长期盐碱胁迫对藜科植物的形态和生理变化影响显著。例如, 雾冰藜(Bassia dasyphylla)在长期盐碱胁迫下表型可塑性发生相应变化, 株高、茎粗、主根根长、侧根数、生物量等指标总体呈减小趋势[19];钠猪毛菜(Salsola nitraria)和盐生草(Halogeton glomeratus)在长期盐胁迫下生长发育、生物量积累和分配受到显著影响, 根冠比呈下降趋势[20];碱蓬(Suaeda glauca)、盐角草(Salicornia europaea)和盐地碱蓬(Suaeda salsa)在长期盐胁迫下根系生长受到抑制[21];藜(Chenopodium album)在长期盐胁迫下生长受到明显抑制, 形态结构发生一定程度改变, 含水量和叶绿素含量显著降低[22]。已有研究多集中于长期盐碱胁迫下藜科植物的形态结构、生物量积累及分配变化, 而对不同浓度、不同盐类长期胁迫下, 藜科植物能否存活、繁殖状况及盐耐受性研究相对较少。
河西走廊地处我国西北干旱内陆地区, 绿洲镶嵌在广袤的荒漠之中, 由于深居内陆、干旱少雨、风沙活动频繁, 绿洲沙漠化严重, 在荒漠绿洲边缘建设人工固沙林成为当地重要的防风固沙措施之一。随着人工林种植年限的增加, 以Na+为主的盐分离子在不同土层均有明显增加, 且累积速率逐渐加快[6]。本研究以河西走廊荒漠绿洲边缘典型藜科一年生草本植物雾冰藜、刺沙蓬(Salsola ruthenica)和白茎盐生草(Halogeton arachnoideus)为研究对象, 研究3种植物在不同浓度盐分(NaCl和NaHCO3)胁迫下的生长、繁殖、生物量分配模式, 以及对盐分的敏感性和耐受性, 旨在了解长期盐碱胁迫下藜科一年生草本植物的生长特性和生物量分配模式, 以期为深入了解藜科一年生草本植物在盐碱生境的适应机制提供参考, 进而为开展盐碱地治理及人工林草本层植被恢复提供科学依据。
1 研究地区与研究方法 1.1 研究区概况研究区位于甘肃河西走廊典型荒漠绿洲边缘地带(39°21′ N, 100°07′ E, 海拔1367 m)。气候属于温带荒漠大陆性气候, 年降水量117.1 mm, 年均蒸发量2337.6 mm;年均温7.6 ℃, 最高气温约为39.1 ℃, 最低气温约为-27 ℃, 无霜期约为105 d;年平均风速3.2 m/s, 风沙活动主要集中在3—5月, 主风向为西北风。研究区土壤类型以风沙土和灰棕漠土为主。为防止风沙侵袭, 促进绿洲边缘沙生植被恢复, 维持绿洲农业生态系统稳定, 当地以梭梭(Haloxylon ammodendron)为优势种建立人工固沙林, 伴生种主要有天然生长的沙拐枣(Calligonum mongolicum)、泡泡刺(Nitraria sphaerocarpa)、红砂(Reaumuria songarica)等灌木半灌木树种, 人工林内雾冰藜、刺沙蓬、白茎盐生草等藜科一年生草本植物分布广泛且繁殖能力强, 成为人工固沙植被草本层的优势植物种, 在防止近地表风蚀发挥着不可忽略的作用。
1.2 试验设计 1.2.1 供试材料2018年10月在中国科学院临泽内陆河流域研究站附近盐碱地采集雾冰藜、刺沙蓬和白茎盐生草种子, 每种植物随机选择10株不同大小植株, 采集完好成熟的种子, 干燥后备用。
1.2.2 试验方法试验在中国科学院临泽内陆河流域研究站内进行, 依据苏永中等[6]和吕彪等[7]关于河西走廊荒漠绿洲过渡带土壤盐分组成的研究, 选取NaCl和NaHCO3进行盐胁迫试验, 参考刘正祥等[23]和郝汉等[24]的盐胁迫试验, 以及Flowers等[25]关于盐生植物的定义, 设置4个盐分梯度, 为对照(0), 轻度盐胁迫(50 mmol/L)、中度盐胁迫(100、150 mmol/L)和高度盐胁迫(200 mmol/L)。
盆栽培植试验参照李辛等[8]和王树凤等[26]的试验方法, 时间为2019年5月18日—10月15日, 花盆直径和深度均为30 cm, 花盆内盛装洗净的细河沙和蛭石, 3种植物每种处理3盆为3个重复。以含有相应浓度的NaCl和NaHCO3溶液为处理液, 每盆1000 mL分4次浇透花盆, 对照组分4次浇灌等量蒸馏水。所有花盆以相应浓度盐溶液浇透后, 开始进行播种。选取子粒饱满、大小基本一致且无病虫的3种藜科植物种子, 每种单独播种于花盆内, 每种植物播种27盆, 共81盆, 每盆播种30粒, 行距3 cm, 播深2 cm(图 1)。为防止系统误差, 所有花盆随机摆放并用标签纸进行标记。播种次日起以称重法补充蒸馏水, 为防止盆内土壤盐分随水分流失, 花盆下均放置托盘, 以将流出的溶液重新倒回盆内。幼苗长成后, 每盆保留3株有6—10片真叶, 株高约为5 cm, 长势良好, 生长状态一致的幼株继续胁迫试验。当3种植物均进入果熟末期(90%种子成熟)时结束试验, 统一收取植株。

|
| 图 1 盆栽种植(行距、株距)示意图 Fig. 1 The schematic diagram of pot planting (row spacing, plant spacing) |
植株存活率计算:分别计算每个处理下3个重复的植株存活率、取平均值。
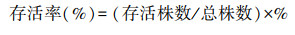
|
(1) |
形态参数:用精确到0.01 cm的直尺测量每株植物的株高和根长, 并记录叶片数。株高是测量根部以上的部分, 以根与茎的分节点为起点, 测量其到主茎顶部的距离。根长是测量植物的主根根长。
生物量及其分配比例计算:将每株植物分割为地上和地下部分, 地上部分又分为种子、茎和叶。将种子、根、茎和叶置于恒温80 ℃的烘箱中烘至质量恒定, 称量干重分别得到茎叶生物量、种子生物量和根系生物量, 其中根系生物量即为地下生物量(belowground biomass, BGB)。根据以下公式计算地上生物量(aboveground biomass, AGB)、总生物量(total biomass, TB)、根冠比(root shoot ratio, R/S)和繁殖分配。
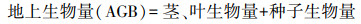
|
(2) |
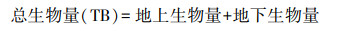
|
(3) |
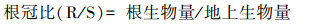
|
(4) |
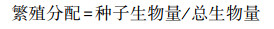
|
(5) |
盐敏感指数和盐耐受指数计算:

|
(6) |
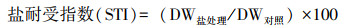
|
(7) |
式中, DW盐处理表示盐胁迫处理下植株干重, DW对照表示对照处理下植株干重[27-28]。
1.3 数据处理利用SPSS 21.0和Excel 2010软件对数据进行统计分析, 利用one-way ANOVA和Duncan法进行方差分析和显著性检验(α=0.05), 利用Origin 8.0软件作图。
2 结果与分析 2.1 钠盐胁迫下藜科一年生草本植物存活率及生长状况 2.1.1 植株存活率3种植物存活率均随NaCl和NaHCO3浓度的增加呈下降趋势(表 1):雾冰藜和刺沙蓬的存活率在50 mmol/L盐浓度下均能达到88.9%以上, 白茎盐生草则达到100%;盐浓度为200 mmol/L时, 雾冰藜和刺沙蓬无法存活或存活率极低, 而白茎盐生草在200 mmol/L NaCl胁迫下存活率仍能达到66.7%;此外, 雾冰藜和刺沙蓬存活率的下降幅度显著高于白茎盐生草。
| 盐胁迫 Salt stress |
物种 Species |
盐分浓度Salt concentration/(mmol/L) | ||||
| 0 | 50 | 100 | 150 | 200 | ||
| NaCl胁迫 | 雾冰藜Bassia dasyphylla | 100±0.0a | 88.9±11.1a | 77.8±11.1a | 33.3±19.2b | 0.0±0.0b |
| NaCl stress | 刺沙蓬Salsola ruthenica | 100±0.0a | 100.0±0.0a | 66.7±19.2ab | 33.3±19.2bc | 22.2±11.1c |
| 白茎盐生草Halogeton arachnoideus | 100±0.0a | 100.0±0.0a | 100.0±0.0a | 77.8±11.1a | 66.7±19.2a | |
| NaHCO3胁迫 | 雾冰藜Bassia dasyphylla | 100±0.0a | 88.9±11.1a | 66.7±19.2ab | 33.3±19.2bc | 0.0±0.0c |
| NaHCO3 stress | 刺沙蓬Salsola ruthenica | 100±0.0a | 88.9±11.1ab | 66.7±19.2ab | 55.6±11.1b | 0.0±0.0c |
| 白茎盐生草Halogeton arachnoideus | 100±0.0a | 100.0±0.0a | 77.8±22.2a | 66.7±19.2a | 0.0±0.0b | |
| 图中数据均为平均值±标准误, 不同字母表示不同浓度盐胁迫处理下差异显著(P < 0.05) | ||||||
不同浓度NaCl和NaHCO3胁迫下, 3种植物的株高生长差异显著(P < 0.05)(图 2):雾冰藜和刺沙蓬的平均株高随盐浓度的增加呈下降趋势, NaCl胁迫下最低值为最高值的60.2%和46.6%, NaHCO3胁迫下为51.2%和41.2%;白茎盐生草的平均株高呈先升后降趋势, 在100 mmol/L盐胁迫下最大。

|
| 图 2 不同NaCl和NaHCO3浓度下3种藜科植物的植株高度、叶片数量和主根长 Fig. 2 Plant height, leaf number and main root length of three Chenopodiaceae annuals under different concentrations of NaCl and NaHCO3 图中误差棒为平均值的标准误, N表示死亡 |
不同浓度NaCl和NaHCO3胁迫下, 3种植物的叶片数量差异显著(图 2):雾冰藜的平均叶片数量分别在0 mmol/L NaCl和50 mmol/L NaHCO3浓度下最大, 随盐浓度的增加呈下降趋势, NaCl胁迫下最低值为最高值的42.5%, NaHCO3胁迫下为23.0%;刺沙蓬的平均叶片数量随盐浓度的增加呈下降趋势, NaCl胁迫下最低值为最高值的26.6%, NaHCO3胁迫下为25.6%;白茎盐生草的平均叶片数量分别在200 mmol/L NaCl和100 mmol/L NaHCO3浓度下最大。
不同浓度NaCl和NaHCO3胁迫下, 雾冰藜和刺沙蓬根长差异显著(图 2), 平均根长随盐浓度的增加呈下降趋势, NaCl胁迫下最低值为最高值的56.7%和58.1%, NaHCO3胁迫下为61.6%和60.2%;白茎盐生草根长差异不显著。
2.2 钠盐胁迫下藜科一年生草本植物的生物量不同浓度NaCl胁迫下, 刺沙蓬和白茎盐生草的AGB、BGB和TB积累差异显著, 刺沙蓬的生物量积累随盐浓度的增加呈下降趋势, 最低值为最高值的5.4%、5.0%和5.3%, 白茎盐生草生物量积累随盐浓度的增加呈先升后降趋势, 150 mmol/L浓度下最大;雾冰藜的生物量积累差异不显著(表 2)。
| 盐胁迫 Salt stress |
物种 Species |
指标 Targets |
盐浓度Salt concentration/(mmol/L) | ||||
| 0 | 50 | 100 | 150 | 200 | |||
| NaCl胁迫 | 雾冰藜Bassia dasyphylla | AGB/g | 0.59±0.09a | 0.47±0.05a | 0.64±0.08a | 0.45±0.03a | N |
| NaCl stress | BGB/g | 0.05±0.01a | 0.04±0.01a | 0.05±0.01a | 0.04±0.00a | N | |
| TB/g | 0.64±0.10a | 0.51±0.05a | 0.69±0.10a | 0.49±0.03a | N | ||
| 刺沙蓬Salsola ruthenica | AGB/g | 1.11±0.12a | 0.33±0.14b | 0.33±0.09b | 0.26±0.02b | 0.06±0.00b | |
| BGB/g | 0.20±0.06a | 0.03±0.01b | 0.02±0.01b | 0.02±0.00b | 0.01±0.00b | ||
| TB/g | 1.31±0.17a | 0.36±0.15b | 0.35±0.09b | 0.28±0.02b | 0.07±0.01b | ||
| 白茎盐生草Halogeton arachnoideus | AGB/g | 0.14±0.02b | 0.11±0.01b | 0.39±0.03a | 0.38±0.05a | 0.37±0.10a | |
| BGB/g | 0.02±0.00b | 0.02±0.00b | 0.03±0.00ab | 0.05±0.01a | 0.04±0.01a | ||
| TB/g | 0.16±0.02b | 0.13±0.01b | 0.42±0.03a | 0.43±0.05a | 0.41±0.10a | ||
| NaHCO3胁迫 | 雾冰藜Bassia dasyphylla | AGB/g | 0.59±0.09b | 1.26±0.30a | 0.71±0.09b | 0.43±0.08b | N |
| NaHCO3 stress | BGB/g | 0.05±0.01ab | 0.14±0.13a | 0.05±0.02ab | 0.03±0.01b | N | |
| TB/g | 0.64±0.10b | 1.40±0.36a | 0.76±0.10b | 0.46±009b | N | ||
| 刺沙蓬Salsola ruthenica | AGB/g | 1.11±0.12a | 0.68±0.16b | 0.22±0.04c | 0.22±0.04c | N | |
| BGB/g | 0.20±0.05a | 0.06±0.01b | 0.02±0.00b | 0.02±0.00b | N | ||
| TB/g | 1.31±0.17a | 0.74±0.18b | 0.24±0.04c | 0.24±0.04c | N | ||
| 白茎盐生草Halogeton arachnoideus | AGB/g | 0.14±0.02b | 0.11±0.01b | 0.29±0.02a | 0.28±0.01a | N | |
| BGB/g | 0.02±0.00bc | 0.01±0.00c | 0.04±0.00a | 0.03±0.01ab | N | ||
| TB/g | 0.16±0.02b | 0.12±0.01b | 0.33±0.03a | 0.31±0.01a | N | ||
| AGB:地上生物量, aboveground biomass;BGB:地下生物量, belowground biomass;TB:总生物量, total biomass | |||||||
不同浓度NaHCO3胁迫下, 3种植物的AGB、BGB和TB积累差异显著:雾冰藜的生物量积累在50 mmol/L浓度下最大, 随盐浓度的增加呈下降趋势, 最低值为最高值的34.1%、21.4%和32.9%;刺沙蓬的生物量积累随盐浓度的增加呈下降趋势, 最低值为最高值的19.8%、10.0%和18.3%;白茎盐生草生物量积累随盐浓度的增加呈先升后降趋势, 100 mmol/L浓度下最大。
2.3 钠盐胁迫下藜科一年生草本植物的生物量分配 2.3.1 钠盐胁迫下藜科一年生草本植物不同器官的生物量分配不同浓度NaCl胁迫下, 3种植物的茎、叶生物量分配随盐浓度的增加呈上升趋势, 雾冰藜和刺沙蓬在150 mmol/L浓度下最大, 比对照组增加13.9%和15.6%, 白茎盐生草在200 mmol/L浓度下最大, 比对照组增加13.5%;根系生物量分配呈下降趋势;繁殖分配呈先升后降趋势, 50 mmol/L浓度下雾冰藜、刺沙蓬和白茎盐生草的繁殖分配比对照组分别增加8.0%、1.4%和8.0%(图 3)。

|
| 图 3 不同NaCl和NaHCO3浓度下3种藜科植物的生物量分配 Fig. 3 Biomass allocation of three Chenopodiaceae annuals under different concentrations of NaCl and NaHCO3 *表示同一物种在不同浓度盐胁迫处理下差异显著(P < 0.05) |
不同浓度NaHCO3胁迫下, 3种植物的茎、叶生物量分配随盐浓度的增加呈上升趋势, 均在150 mmol/L浓度下最大, 比对照组分别增加11.1%、12.1%和11.9%;根系生物量分配呈下降趋势;繁殖分配呈先升后降趋势, 雾冰藜和白茎盐生草在50 mmol/L浓度下比对照组增加41.1%和44.3%, 刺沙蓬在100 mmol/L浓度下比对照组增加66.3%(图 3)。
2.3.2 钠盐胁迫下藜科一年生草本植物的地上、地下生物量分配根冠比是植物地上、地下生物量分配策略的体现, 由图 4可以看出, 不同浓度NaCl胁迫下, 刺沙蓬和白茎盐生草的根冠比差异显著, 随盐浓度的增加呈下降趋势, 最低值为最高值的36.8%和78.9%;雾冰藜的根冠比差异不显著。不同浓度NaHCO3胁迫下, 白茎盐生草根冠比差异显著, 随盐浓度的增加呈下降趋势, 最低值为最高值的75.5%;雾冰藜和刺沙蓬根冠比差异不显著。

|
| 图 4 不同NaCl和NaHCO3浓度下3种藜科植物的根冠比 Fig. 4 Root shoot ratio of three Chenopodiaceae annuals under different concentrations of NaCl and NaHCO3 |
盐敏感指数在一定程度上能反映盐分对植物不同部位生长影响的大小, 某一部位盐敏感指数越小, 该部位对盐胁迫越敏感。由图 5可以看出, 不同浓度NaCl胁迫下, 雾冰藜和刺沙蓬不同部位的盐敏感指数随NaCl浓度增加总体呈下降趋势, 且根部盐敏感性始终高于地上部;白茎盐生草不同部位的盐敏感指数总体呈上升趋势, NaCl浓度为100—200 mmol/L时根部盐敏感性要高于地上部。不同浓度NaHCO3胁迫下, 雾冰藜不同部位的盐敏感指数均呈先升后降的趋势, NaHCO3浓度为100、150 mmol/L时根部盐敏感性要高于地上部;刺沙蓬不同部位的盐敏感指数均呈下降趋势, 而白茎盐生草不同部位的盐敏感指数均呈上升趋势, 且两者的根部盐敏感性始终高于地上部。从植物整体的盐耐受指数来看(表 3), 3种植物对NaCl和NaHCO3的耐受性不同, 白茎盐生草的盐耐受性要高于雾冰藜和刺沙蓬, 且3种植物的盐耐受指数随盐浓度增加的变化趋势与盐敏感指数的变化趋势相似, 表现为盐敏感性增强则盐耐受性降低, 而盐敏感性减弱则盐耐受性增强。

|
| 图 5 不同NaCl和NaHCO3浓度下3种藜科植物的盐敏感指数 Fig. 5 Salt sensitive index of three Chenopodiaceae annuals under different concentrations of NaCl and NaHCO3 |
| 物种 Species |
NaCl浓度 NaCl concentration/(mmol/L) |
NaHCO3浓度 NaHCO3 concentration/(mmol/L) |
|||||||||
| 0 | 50 | 100 | 150 | 200 | 0 | 50 | 100 | 150 | 200 | ||
| 雾冰藜 Bassia dasyphylla |
100 | 76.7 | 104.3 | 72.4 | N | 100 | 209.7 | 113.5 | 69.6 | N | |
| 刺沙蓬 Salsola ruthenica |
100 | 27.6 | 27.2 | 21.2 | 5.4 | 100 | 56.8 | 18.6 | 18.4 | N | |
| 白茎盐生草 Halogeton arachnoideus |
100 | 80.4 | 276.8 | 325.6 | 338.1 | 100 | 75.7 | 219.0 | 211.2 | N | |
本研究发现, 钠盐胁迫对3种植物的存活率有明显的抑制作用, 存活率均随盐胁迫的加剧逐渐下降。其中, 雾冰藜和刺沙蓬在NaCl和NaHCO3浓度达到200 mmol/L时基本无法存活, 而白茎盐生草在200 mmol/L NaCl胁迫仍能存活, 但在200 mmol/L NaHCO3胁迫下无法存活, 这表明藜科一年生草本植物对不同盐分胁迫的适应是有阈值的, 而这个阈值会因物种不同有所差异。Flowers等[25]认为能够在≥200 mmol/L NaCl胁迫下生长并完成生活史的植物, 即可定义为盐生植物, 而不能完成生活史的植物为非盐生植物。本研究中, 白茎盐生草在200 mmol/L NaCl胁迫下存活率仍能达到66.7%, 为盐生植物, 而雾冰藜和刺沙蓬为非盐生植物。
同时, 本研究还发现, 一定浓度的盐分胁迫对雾冰藜和白茎盐生草的生长和生物量积累有促进作用, 雾冰藜的生物量在50 mmol/L NaHCO3和100 mmol/L NaCl胁迫下与对照组相比有所增加, 白茎盐生草的株高、叶数和生物量在100、150 mmol/L盐胁迫下与对照组相比有所增加, 表明二者均能耐受一定程度的盐胁迫。有研究表明, 对于一些耐盐植物而言, 低盐胁迫可以促进其生长, 而高盐胁迫表现出抑制作用。例如, 藜科植物藜和灰绿藜(Chenopodium glaucum)生长均受中度盐分胁迫(100—150 mmol/L)的促进, 高度盐分胁迫(>200 mmol/L)下才受抑制[12]。干旱荒漠生态系统中, 一定浓度的盐离子渗入植物体内会降低其内部的渗透势, 促使植物具有渗透适应性, 增强植物的吸水保水能力[29], 不仅可以促进植物的生长发育, 还能在一定程度上增强植物的抗旱能力, 但盐分浓度过高则会产生渗透胁迫和离子毒害, 植物生长受到抑制甚至死亡[30]。但不同植物对盐分的敏感程度不同, 同一盐分浓度对敏感植物来说可能是高盐胁迫, 而对抗盐性较强的植物来说却是低盐胁迫[31]。
3.2 钠盐胁迫对藜科一年生草本植物生物量分配的影响本研究发现, 钠盐胁迫下3种植物的生物量分配模式发生变化。3种植物的根冠比随盐胁迫的加剧总体呈减小趋势, 其中根系生物量分配随盐浓度的增加呈下降趋势, 而地上部分的茎叶生物量分配和繁殖分配在不同盐分浓度下有所增加, 意味着植物在盐胁迫加剧的情况下会将更多的同化产物用于地上部分的生长和繁殖, 对根系的投入减少。生物量分配模式的改变是由不同部位的盐敏感性不同引起的[32], 随着盐浓度的增加, 3种植物根部的盐敏感性要高于地上部, 这可能是造成植物根系生物量分配随盐浓度增加而下降的重要原因。弋良鹏等对一些滨海盐生植物的研究发现, 碱蓬、盐角草等根部的盐敏感性随盐胁迫的加剧高于地上部, 高盐胁迫下植物的根冠比降低[21], 与本文的研究结果类似。盐胁迫下调节生物量分配模式是植物适应盐胁迫的方式之一[29], 根部是暴露于盐胁迫环境中的首要组织, 是最先受到盐胁迫伤害和对盐分最为敏感的部分[21], 盐胁迫下, 植物降低对根系生物量的分配, 能降低根系对盐分的吸收, 减少向地上部的盐分输送, 从而增强植物抵抗盐害的能力。
同时, 本研究还发现, 3种藜科植物在盐分胁迫下增加对地上部分的生物量分配, 但低盐胁迫下(50、100 mmol/L)主要增加繁殖分配, 提高繁殖输出, 而盐胁迫加剧时, 主要增加茎、叶生物量分配, 繁殖分配比例下降。Wertis等[33]通过对三角叶滨藜(Atriplex triangularis)的研究发现, 三角叶滨藜的繁殖分配随盐胁迫的加剧呈先升后降趋势, 与本文的研究结果类似。一年生草本植物作为机会主义者, 在应对胁迫环境时具有灵活的生长和繁殖适应策略[34], 轻度盐胁迫下, 一年生植物会增加对繁殖体的资源分配以达到最大的繁殖数量, 来保证繁殖成功和种族延续[35], 而胁迫加剧时, 植物生长受阻, 过多的繁殖分配不利于植物存活, 此时增加对营养体的资源分配能提高资源的获取能力[33;36], 以抵御不利的生存环境从而保证完成生活史, 这种资源分配的可塑性也是一年生草本植物适应盐碱胁迫的策略之一。
3.3 藜科一年生草本植物的耐盐性本研究发现, NaHCO3对3种藜科植物的胁迫作用强于NaCl。不同浓度NaHCO3胁迫下植株存活率较低, NaHCO3浓度达到200 mmol/L时3种植物均不能存活, 而NaCl浓度达到200 mmol/L时有两种植物存活, 同时NaHCO3胁迫下, 植株存活率随胁迫加剧的下降幅度高于NaCl胁迫。相关系数绝对值的大小可以反映盐胁迫因素对植物生长指标影响作用的强弱[19], 从表 4可以看出, 钠盐胁迫因素与3种藜科植物生长指标间的相关系数中, 70%达到显著或极显著水平, 61%表现为NaHCO3胁迫作用大于NaCl。NaHCO3胁迫下3种植物的生长状况较差, 与相同浓度NaCl胁迫相比, 雾冰藜的株高和叶数在150 mmol/LNaHCO3胁迫下降低13.9%和20.2%, 刺沙蓬的株高、叶数和总生物量在100 mmol/LNaHCO3胁迫下降低33.8%、33.8%和31.4%, 白茎盐生草的株高、叶数和总生物量在100—150 mmol/LNaHCO3胁迫下降低21.3%—32.6%。另外, 中度NaHCO3胁迫下(100、150 mmol/L)3种植物的整体盐耐受性较弱, 为相同浓度NaCl胁迫下的64.9%—96.1%。这与禾本科虎尾草(Chloris virgata)的研究结果一致[37]。两者虽都属于钠盐, 但NaCl是中性盐, NaHCO3是碱性盐, 碱性盐对植物的胁迫因素除了和中性盐共有的渗透胁迫和离子毒害外, 还有高PH值及明显降低矿质元素的可利用性等方面, 对植物的破坏强于中性盐[38], 这也从另一层面说明碱性盐对一年生草本植物的胁迫作用强于中性盐。
| 盐分 Salt |
物种 Species |
株高 Plant height/cm |
叶片数 Leaf number |
主根长 Main root length/cm |
地上生物量 Aboveground biomass/g |
地下生物量 Belowground biomass/g |
总生物量 Total biomass/g |
| NaCl | 雾冰藜Bassia dasyphylla | -0.789** | -0.693** | -0.765** | -0.174 | -0.263 | -0.192 |
| 刺沙蓬Salsola ruthenica | -0.554** | -0.584** | -0.841** | -0.756** | -0.632** | -0.746** | |
| 白茎盐生草Halogeton arachnoideus | 0.247 | 0.581** | 0.139 | 0.788** | 0.639** | 0.781** | |
| NaHCO3 | 雾冰藜Bassia dasyphylla | -0.777** | -0.490** | -0.716** | -0.247 | -0.244 | -0.248 |
| 刺沙蓬Salsola ruthenica | -0.687** | -0.741** | -0.833** | -0.820** | -0.682** | -0.814** | |
| 白茎盐生草Halogeton arachnoideus | 0.167 | 0.193 | -0.284 | 0.799** | 0.503* | 0.782** | |
| **表示P < 0.01, *表示P < 0.05 | |||||||
同时, 本文研究还发现, 白茎盐生草的耐盐能力强于雾冰藜和刺沙蓬。200 mmol/L NaCl胁迫下雾冰藜和刺沙蓬的存活率为白茎盐生草的0%和33.3%, 150 mmol/L NaHCO3胁迫下为白茎盐生草的49.9%和83.4%。此外, 白茎盐生草的盐敏感性弱于雾冰藜和刺沙蓬, 且整体的盐耐受性较强。这可能是因为白茎盐生草根系的盐敏感性远低于雾冰藜和刺沙蓬, 且受盐胁迫影响较小。根系与土壤直接接触, 对土壤环境更为敏感且易对土壤环境做出反应[39], 盐胁迫环境下, 不同物种根系的生长发育和生物量积累反映了植物抵御不利环境因素的能力[38]。本研究中, 雾冰藜和刺沙蓬的根长和根系生物量在钠盐胁迫下受到明显抑制, 随盐胁迫加剧逐渐下降, 而白茎盐生草受钠盐胁迫影响较小。由此可见, 藜科一年生草本植物耐盐性的强弱可能与根系的耐盐性和受盐胁迫的影响程度有关。
| [1] |
李辛, 赵文智. 雾冰藜(Bassia dasyphylla)种子萌发和幼苗生长对盐碱胁迫的响应. 中国沙漠, 2018, 38(2): 300-306. |
| [2] |
罗毅. 干旱区绿洲滴灌对土壤盐碱化的长期影响. 中国科学: 地球科学, 2014, 44(8): 1679-1688. |
| [3] |
王蕙, 赵文智, 常学向. 黑河中游荒漠绿洲过渡带土壤水分与植被空间变异. 生态学报, 2007, 27(5): 1731-1739. DOI:10.3321/j.issn:1000-0933.2007.05.008 |
| [4] |
杨新民, 李玲燕. 西北地区生态环境存在问题与生态修复对策. 水土保持研究, 2005, 12(5): 98-100, 106-106. DOI:10.3969/j.issn.1005-3409.2005.05.021 |
| [5] |
马松尧, 王刚, 杨生茂. 西北地区荒漠化防治与生态恢复若干问题的探讨. 水土保持通报, 2004, 24(5): 105-108. DOI:10.3969/j.issn.1000-288X.2004.05.025 |
| [6] |
苏永中, 刘婷娜. 流动沙地建植人工固沙梭梭林的土壤演变过程. 土壤学报, 2020, 57(1): 84-91. |
| [7] |
吕彪, 许耀照, 赵芸晨. 河西走廊内陆盐渍土生物修复与调控研究. 水土保持通报, 2008, 28(3): 198-200. |
| [8] |
李辛, 赵文智. 荒漠区植物雾冰藜光合特性对混合盐碱胁迫的响应. 生态学报, 2018, 38(4): 1183-1193. |
| [9] |
王景瑞, 王立, 徐先英, 闫好原, 胡生新, 高承兵, 魏林源. 干旱荒漠区4种一年生植物种子萌发期耐盐性. 草业科学, 2020, 37(2): 237-244. |
| [10] |
刘涛, 耿文春, 李丽, 刘一佳, 刘慧民. 混合盐碱胁迫对两种抗性不同的绣线菊光合特性的影响. 东北农业大学学报, 2009, 40(5): 32-36. DOI:10.3969/j.issn.1005-9369.2009.05.007 |
| [11] |
刘晓静, 张晓磊, 齐敏兴, 刘艳楠. 混合盐碱对紫花苜蓿种子萌发及幼苗期叶绿素荧光特性的影响. 草地学报, 2013, 21(3): 501-507. |
| [12] |
王璐, 蔡明, 兰海燕. 藜科植物藜与灰绿藜耐盐性的比较. 植物生理学报, 2015, 51(11): 1846-1854. |
| [13] |
宁建凤, 郑青松, 杨少海, 邹献中, 孙丽丽, 陈勇. 高盐胁迫对罗布麻生长及离子平衡的影响. 应用生态学报, 2010, 21(2): 325-330. |
| [14] |
党荣理, 潘晓玲. 西北干旱荒漠区种子植物科的区系分析. 西北植物学报, 2002, 22(1): 24-32. DOI:10.3321/j.issn:1000-4025.2002.01.004 |
| [15] |
赵哈林, 大黑俊哉, 李玉霖, 左小安, 黄刚, 周瑞莲. 人类放牧活动与气候变化对科尔沁沙质草地植物多样性的影响. 草业学报, 2008, 17(5): 1-8. DOI:10.3321/j.issn:1004-5759.2008.05.001 |
| [16] |
袁建立, 张景光, 王刚. 沙坡头人工固沙区草本层片组成及其动态研究. 中国沙漠, 2002, 22(6): 623-627. DOI:10.3321/j.issn:1000-694X.2002.06.018 |
| [17] |
张景光, 周海燕, 王新平, 李新荣, 王刚. 沙坡头地区一年生植物的生理生态特性研究. 中国沙漠, 2002, 22(4): 350-353. DOI:10.3321/j.issn:1000-694X.2002.04.008 |
| [18] |
刘志民, 李雪华, 李荣平, 蒋德明, 曹成有, 常学礼. 科尔沁沙地31种1年生植物萌发特性比较研究. 生态学报, 2004, 24(3): 648-653. DOI:10.3321/j.issn:1000-0933.2004.03.036 |
| [19] |
李辛. 一年生植物雾冰藜对盐碱胁迫的响应研究[D]. 兰州: 甘肃农业大学, 2018.
|
| [20] |
贺海波, 李彦. 干旱、盐胁迫条件下两种盐生植物生物量分配对策的研究. 干旱区研究, 2008, 25(2): 242-247. |
| [21] |
弋良朋, 王祖伟. 盐胁迫下3种滨海盐生植物的根系生长和分布. 生态学报, 2011, 31(5): 1195-1202. |
| [22] |
油天钰. 藜科植物藜在盐胁迫下的形态结构及其对盐的生理响应[D]. 乌鲁木齐: 新疆大学, 2010.
|
| [23] |
刘正祥, 张华新, 杨秀艳, 刘涛, 狄文彬. NaCl胁迫下沙枣幼苗生长和阳离子吸收、运输与分配特性. 生态学报, 2014, 34(2): 326-336. |
| [24] |
郝汉, 曹磊, 陈伟楠, 胡增辉, 冷平生. 盐胁迫对槲树(Quercus dentata)幼苗离子平衡及其生理生化特性的影响. 生态学报, 2020, 40(19): 6897-6904. |
| [25] |
Flowers T J, Colmer T D. Salinity tolerance in halophytes. New Phytologist, 2008, 179(4): 945-963. DOI:10.1111/j.1469-8137.2008.02531.x |
| [26] |
王树凤, 胡韵雪, 孙海菁, 施翔, 潘红伟, 陈益泰. 盐胁迫对2种栎树苗期生长和根系生长发育的影响. 生态学报, 2014, 34(4): 1021-1029. |
| [27] |
Rejili M, Vadel A M, Guetet A, Neffatti M. Effect of NaCl on the growth and the ionic balance K+/Na+ of two populations of Lotus creticus (L.) (Papilionaceae). South African Journal of Botany, 2007, 73(4): 623-631. DOI:10.1016/j.sajb.2007.06.006 |
| [28] |
Khayat P N, Jamaati-e-Somarin S, Zabihi-e-Mahmoodabad R, Yari A, Khayatnezhad M, Gholamin R. Screening of salt tolerance canola cultivars (Brassica napus L.) using physiological markers. Word Applied Sciences Journal, 2010, 10(7): 817-820. |
| [29] |
王佺珍, 刘倩, 高娅妮, 柳旭. 植物对盐碱胁迫的响应机制研究进展. 生态学报, 2017, 37(16): 5565-5577. |
| [30] |
廖岩, 彭友贵, 陈桂珠. 植物耐盐性机理研究进展. 生态学报, 2007, 27(5): 2077-2089. DOI:10.3321/j.issn:1000-0933.2007.05.049 |
| [31] |
王国华, 郭文婷, 缑倩倩. 钠盐胁迫对河西走廊荒漠绿洲过渡带典型一年生草本植物种子萌发的影响. 应用生态学报, 2020, 31(6): 1941-1947. |
| [32] |
严青青, 张巨松, 徐海江, 李星星, 王燕提. 盐碱胁迫对海岛棉幼苗生物量分配和根系形态的影响. 生态学报, 2019, 39(20): 7632-7640. |
| [33] |
Wertis B A, Ungar I A. Seed demography and seedling survival in a population of Atriplex triangularis Willd. The American Midland Naturalist, 1986, 116(1): 152-162. DOI:10.2307/2425947 |
| [34] |
张景光, 王新平, 李新荣, 张志山, 王刚, 王桑, 马风云. 荒漠植物生活史对策研究进展与展望. 中国沙漠, 2005, 25(3): 306-314. DOI:10.3321/j.issn:1000-694X.2005.03.002 |
| [35] |
张科, 田长彦, 李春俭. 一年生盐生植物耐盐机制研究进展. 植物生态学报, 2009, 33(6): 1220-1231. DOI:10.3773/j.issn.1005-264x.2009.06.023 |
| [36] |
Willson M F. Plant Reproductive Ecology. New York: John Wiley & Sons, 1983, 1-44. |
| [37] |
李长有. 盐碱地四种主要致害盐分对虎尾草胁迫作用的混合效应与机制[D]. 长春: 东北师范大学, 2009.
|
| [38] |
郎志红. 盐碱胁迫对植物种子萌发和幼苗生长的影响[D]. 兰州: 兰州交通大学, 2008.
|
| [39] |
Lynch J. Root architecture and plant productivity. Plant Physiology, 1995, 109(1): 7-13. DOI:10.1104/pp.109.1.7 |