 2021, Vol. 41
2021, Vol. 41文章信息
- 林惠瑛, 元晓春, 周嘉聪, 曾泉鑫, 孙俊, 程蕾, 林开淼, 徐建国, 程栋梁, 陈岳民
- LIN Huiying, YUAN Xiaochun, ZHOU Jiacong, ZENG Quanxin, SUN Jun, CHENG Lei, LIN Kaimiao, XU Jianguo, CHENG Dongliang, CHEN Yuemin
- 海拔梯度变化对武夷山黄山松林土壤磷组分和有效性的影响
- Effects of different elevational gradients on soil phosphorus fractions and availab ility in Pinus taiwanensis forest on Wuyi Mountain
- 生态学报. 2021, 41(14): 5611-5621
- Acta Ecologica Sinica. 2021, 41(14): 5611-5621
- http://dx.doi.org/10.5846/stxb202008072054
-
文章历史
- 收稿日期: 2020-08-07
- 网络出版日期: 2021-05-13





2. 湿润亚热带山地生态国家重点实验室培训基地, 福州 350007;
3. 福建戴云山国家级自然保护区管理局, 泉州 362500
2. Cultivation Base of State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou 350007, China;
3. Daiyun Mountain National Nature Reserve Administration Bureau, Quanzhou 362500, China
磷(P)是植物生长最重要的养分元素之一, 基于全球范围的研究表明, 在自然陆地生态系统中地上植物生产普遍受到P的限制[1]。其中, P限制现象除了在热带地区广泛存在之外, 还常常发生在亚热带地区[1-2]。土壤中的P以不同的形态存在, 根据其溶解度和生物有效性的不同, 可划分为速效P、易分解态P、中等易分解态P和难利用P等组分[3]。P限制程度和P有效性的变化因气候带和地区的差异而不同, 受温度[4]、水分[5]、土壤pH[6]和微生物活性[7]等因素的影响。因此, 了解亚热带森林土壤P组分状况是预测环境变化(如气候变化)如何影响P循环和P有效性的重要前提[8]。海拔梯度作为一种自然地理变化, 会导致温度、水分、土壤特性、微生物活性等因素的改变[9], 是探究土壤P组分和P有效性如何响应环境变化的重要平台。
目前, 关于不同海拔的土壤P组分以及P有效性的变化规律尚无统一定论。在中国福建戴云山的研究中, 随海拔降低, 土壤各P组分和P有效性增加[10];而在意大利中部山毛榉林则得出相反的结果[11]。此外, 海拔梯度的变化可能改变植被类型, 通过植物的地下过程间接改变土壤P组分。Vincent等[12]研究发现, 北极苔原地区土壤P组分的海拔变化一方面是由于不同植被类型对磷的吸附能力不同, 另一方面因为不同植被导致土壤性质改变, 从而影响P组分的变化。在中国贡嘎山的研究中也发现, 不同海拔植被类型的差异导致土壤pH的改变, 从而影响土壤P组分的海拔分布模式[13]。由此可见, 海拔梯度上植被类型的变化可能会造成土壤P组分对海拔的响应更为复杂。因此, 控制植被类型变量, 探究土壤P组分和P有效性对海拔梯度的响应, 有利于更好理解山地森林生态系统中土壤养分循环的过程。
武夷山国家级自然保护区拥有全球同纬度现存面积最广、保存最完整的亚热带森林生态系统[14], 是亚热带东南大陆海拔梯度最大的区域, 也是当前研究的热点区域。黄山松(Pinus taiwanensis)又称台湾松, 是我国亚热带山区的主要建群种, 也是武夷山绿化和用材的重要优良树种, 且主要分布于武夷山海拔1100 m以上, 具有良好的海拔梯度分布特征[15], 是探究土壤P组分和P有效性对海拔梯度响应的理想平台。因此, 本研究以不同海拔黄山松林为研究对象, 探究土壤P组分和P有效性的变化, 以期为预测土壤P循环对环境变化的响应提供理论依据, 也为探究亚热带山地森林生态系统中土壤养分循环以及武夷山森林保护区可持续发展提供科学依据。
1 研究地区与研究方法 1.1 研究区概况试验区位于江西省武夷山国家级自然保护区(27°48′11″—28°00′35″ N, 117°39′30″—117°55′47″ E), 海拔350—2161 m, 属于亚热带季风气候。年均气温14.2 ℃, 年均降水量2583 mm, 年均湿度84%, 年均蒸发量778 mm, 年均无霜期231 d[14, 16]。
1.2 试验设计2014年10月, 根据江西武夷山国家级自然保护区内黄山松的实际生长及分布情况, 依据随机性和独立性的样地选取原则, 在海拔1200—2000 m之间, 每200 m为1个间隔设置了5个海拔梯度(其中包括2000、1800、1600、1400、1200 m), 并且在每个海拔梯度设置了3个20 m×20 m的样方, 样方之间设有10 m以上的隔离带, 共计15个样方[16]。之后, 对样地内胸径≥5 cm的乔木进行胸径和树高的测量, 样地基本情况详见表 1。试验区土壤主要以山地黄壤为主。
| 2000 m | 1800 m | 1600 m | 1400 m | 1200 m | |
| 郁闭度Canopy closure | 0.50±0.03c | 0.60±0.05b | 0.90±0.33a | 0.86±0.02a | 0.87±0.03a |
| 密度Density/(株/hm2) | 525.00±160.73c | 417.29±130.98c | 1533.36±370.90b | 1991.12±256.73a | 717.25±65.08c |
| 平均胸径/cm Average diameter at breast high |
13.17±0.66b | 14.84±0.69b | 14.53±0.40b | 14.25±0.36b | 23.99±0.79a |
| 平均树高Average height/m | 4.76±0.14e | 6.22±0.20d | 10.49±0.16c | 13.29±0.23b | 16.19±0.46a |
2019年7月在每个样方以“S”型5点取样法采样, 先去除表面凋落物, 然后采集表层土壤(0—10 cm)混合均匀, 放入保温箱带回实验室, 去除石砾、碎屑以及植物根系后测定土壤磷酸酶活性, 一部分土壤过2 mm筛后测定土壤微生物生物量, 另取一部分自然风干后测定土壤性质。
1.3 测定项目与方法土壤环境因素和理化性质的测定:土壤温度(ST)采用温度传感器(Model SK-250WP, Sato, 日本)测定。用玻璃电极pH计(STARTER 300, Ohaus, 美国)测定土壤pH, 土水比为1∶2.5。土壤含水率(SM)用烘干法测定。土壤总有机碳(SOC)和总氮(TN)用碳氮元素分析仪(Elementar Vario EL III, Elementar, 德国)测定。矿质氮用2 mol/L KCl浸提, 用连续流动分析仪(Skalar san++, Skalar, 荷兰)测定滤液中的铵态氮(NH4+-N)和硝态氮(NO3--N)含量[17]。可溶性有机碳(DOC)和可溶性有机氮(DON)用去离子水浸提(土水比1∶4), 用总有机碳分析仪(TOC-VCPH/CPN, Shimadzu, 日本)测定DOC含量, 用连续流动分析仪测定DON含量[18]。
微生物生物量(SMB)的测定:微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸-硫酸钾浸提法[19], MBP采用氯仿熏蒸-NaHCO3浸提法[20-21], 用总有机碳分析仪测定提取液中的总有机碳含量, 用连续流动分析仪测定总氮和磷酸盐含量, 计算公式为:
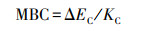
|
(1) |
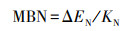
|
(2) |

|
(3) |
式中, ΔEC为熏蒸与未熏蒸土壤总C含量的差值;KC为MBC的浸提系数(0.45);ΔEN为熏蒸与未熏蒸土壤总N含量的差值;KN为MBN的浸提系数(0.54);ΔEP为熏蒸与未熏蒸土壤总P含量的差值;KP为MBP的浸提系数(0.40);Kb为添加KH2PO4后的P回收率[22]。
酸性磷酸酶活性的测定:参照Saiya-Cork[23]和Tian等[24]的方法测定酸性磷酸单酯酶(ACP)和磷酸双酯酶(PD)。分别取1 g新鲜土壤, 分别用125 mL 50 mmol/L的醋酸盐缓冲液(pH=5)和100 mL 50 m mol/L的THAM-硫酸缓冲液(pH=8)提取, 用磁力搅拌器搅拌5 min使其均质化, 用移液器取200 mL移于96孔微孔板。用伞形酮(MUB)作为标准液标示水解酶活性。ACP和PD的底物分别为4-甲基伞形酮磷酸酯(MUP)和bis-4-甲基伞形酮磷酸酯(bis-MUP)。微平板置于暗环境下20℃恒温培养4 h后, 用多功能酶标仪(SpectraMax M5, Molecular Devices, 美国)测定。
土壤P组分的测定:采用Hedley[25]连续浸提法, 用连续流动分析仪测定。参照Fan等[26]和Hou等[3]的方法, 依据各P组分的溶解度和生物有效性不同, 划分为(1)速效P包括树脂膜提取P(Resin-Pi), 是可被植物直接吸收利用的无机磷;(2)易分解P包括NaHCO3提取P(NaHCO3-Pi和NaHCO3-Po);(3)中等易分解P包括NaOH提取P(NaOH-Pi和NaOH-Po);(4)难利用P包括NaOH浸提并经超声处理提取的P(NaOHu-Pi和NaOHu-Po)、HCl提取P(HCl-Pi)以及浓硫酸、高氯酸消解的残余P(Residual-P)。土壤总P(TP)为所有P组分之和。
1.4 数据处理采用SPSS 21.0软件对数据进行统计分析。利用单因素方差分析检验海拔梯度对土壤基本理化性质、微生物生物量、酸性磷酸酶活性以及P组分的影响, 使用最小显著性差异法(LSD法, α=0.05)来比较不同海拔间的差异。用Pearson法对土壤基本理化性质与酸性磷酸酶活性和微生物生物量进行相关性分析。以土壤基本理化性质、微生物生物量和酸性磷酸酶活性为环境因子, 以各P组分含量为物种因子, 利用Canaco 5.0软件进行冗余分析。利用Origin 9.1软件制图。
2 结果与分析 2.1 土壤环境因素和理化性质除了DON外, 不同海拔黄山松林土壤的基本理化性质存在显著差异(表 2)。ST随海拔降低而升高, 且在各海拔之间具有显著差异。随海拔降低, 土壤pH、SM总体呈降低趋势, 且在海拔1200 m和1400 m处较低。SOC、TN、NH4+-N、NO3-N、DOC、DON含量随海拔升高呈现先增加后降低趋势, 其中SOC、TN在海拔2000 m和1800 m高于海拔1400 m和1200 m, 而DOC、DON则相反。
| 2000 m | 1800 m | 1600 m | 1400 m | 1200 m | P | |
| ST/℃ | 17.48±0.42d | 18.59±0.08d | 19.79±0.16c | 20.86±0.04b | 22.88±0.21a | <0.001 |
| pH | 4.34±0.17a | 4.41±0.01a | 4.06±0.10bc | 4.09±0.01b | 3.88±0.16c | 0.001 |
| SM/% | 94.95±0.01a | 58.72±0.09c | 72.15±0.09b | 60.36±0.20c | 52.42±0.04c | <0.001 |
| SOC/(g/kg) | 58.93±9.38ab | 42.97±6.40c | 71.48±5.17a | 41.61±1.30c | 48.69±9.82c | 0.002 |
| TN/(g/kg) | 4.94±0.74a | 3.68±0.58b | 4.91±0.80a | 2.73±0.07b | 3.52±0.71b | 0.007 |
| NH4+-N/(mg/kg) | 21.95±1.84b | 11.21±1.46c | 33.96±0.62a | 20.19±4.91b | 14.05±3.41c | <0.001 |
| NO3--N/(mg/kg) | 3.51±1.62ab | 2.92±0.79ab | 4.35±0.29a | 0.93±0.22b | 1.21±0.44b | 0.002 |
| DOC/(mg/kg) | 131.14±13.37c | 115.02±27.87c | 239.85±9.83a | 178.58±13.94b | 136.29±21.78b | <0.001 |
| DON/(mg/kg) | 9.34±2.56c | 8.38±1.44c | 15.91±2.90a | 13.34±4.34a | 10.70±2.68b | 0.058 |
| 表中数据为平均值±标准差, 同一行不同小写字母表示不同海拔间差异显著(P < 0.05); ST: 土壤温度Soil temperature; pH: 酸碱度; SM: 土壤含水率Soil moisture; SOC: 总有机碳Soil organic carbon; TN: 总氮Total nitrogen; NH4+-N: 铵态氮; NO3--N: 硝态氮; DOC: 可溶性有机碳Dissolved organic carton; DON: 可溶性有机氮Dissolved organic nitrogen | ||||||
随着海拔降低, MBC、MBN、MBP含量总体呈降低的趋势, 其中MBP在海拔2000、1800 m和1600 m显著高于海拔1400 m和1200 m。MBC/MBN在海拔1400 m最低, MBC/MBP在海拔1200 m最高, MBN/MBP在海拔2000、1800 m和1600 m显著低于海拔1400 m和1200 m(图 1)。

|
| 图 1 不同海拔黄山松林土壤微生物生物量及其化学计量比的变化 Fig. 1 Changes of microbial biomass and its stoichiometric ratio of soil in Pinus taiwanensis forest at different elevations 图中数据为平均值±标准差, 同一行不同小写字母表示不同海拔间差异显著(P < 0.05); MBC: 微生物生物量碳Microbial biomass carbon; MBN: 微生物生物量氮Microbial biomass nitrogen; MBP: 微生物生物量磷; Microbial biomass phosphorus. SMB: 微生物生物量Microbial biomass |
PD、ACP随海拔降低呈先增加后降低趋势, 在海拔1400 m最高, 海拔1600、1200 m次之。同样地, PD/MBC、ACP/MBC随海拔降低也呈现先增后降趋势, 且在海拔1400 m最高, 海拔1200 m次之(图 2)。

|
| 图 2 不同海拔黄山松林土壤酸性磷酸酶活性及每单位微生物量生物量碳酶活性的变化 Fig. 2 Changes of soil acid phosphatase activities and mass-specific enzyme activities at different elevations 图中数据为平均值±标准差, 同一行不同小写字母表示不同海拔间差异显著(P < 0.05); ACP: 酸性磷酸单酯酶acid phosphomonoesterase; PD: 酸性磷酸双酯酶acid phosphodiesterase; MBC: 微生物生物量碳Microbial biomass carbon |
PD、ACP均与ST、DOC和DON呈显著正相关;MBC、MBN、MBP与ST呈负相关, 而与SOC、TN呈正相关;MBC、MBN与SM呈显著正相关(表 3)。
| 指标Index | PD | ACP | MBC | MBN | MBP |
| ST | 0.562* | 0.539* | -0.511 | -0.735** | -0.743** |
| pH | -0.521* | -0.419 | 0.264 | 0.510 | 0.648** |
| SM | -0.125 | -0.156 | 0.534* | 0.888** | 0.405 |
| SOC | -0.081 | 0.006 | 0.691** | 0.398 | 0.59** |
| TN | -0.423 | -0.353 | 0.813** | 0.654** | 0.693** |
| NH4+-N | 0.367 | 0.480 | 0.417 | 0.232 | 0.260 |
| NO3--N | -0.431 | -0.288 | 0.630* | 0.481 | 0.802** |
| DOC | 0.545* | 0.678** | 0.146 | -0.170 | 0.042 |
| DON | 0.572* | 0.608* | 0.044 | -0.083 | -0.161 |
| ACP: 酸性磷酸单酯酶acid phosphomonoesterase; PD: 酸性磷酸双酯酶acid phosphodiesterase; MBP: 微生物生物量磷Microbial biomass phosphorus; MBN: 微生物生物量氮Microbial biomass nitrogen; MBC: 微生物生物量碳Microbial biomass carbon; ST: 土壤温度Soil temperature; pH: 酸碱度; SM: 土壤含水率Soil moisture; SOC: 总有机碳Soil organic carbon; TN: 总氮Total nitrogen; NH4+-N: 铵态氮; NO3--N: 硝态氮; DOC: 可溶性有机碳Dissolved organic carton; DON: 可溶性有机氮Dissolved organic nitrogen. * P < 0.05; ** P < 0.01. -表示pearson相关系数为负值 | |||||
不同海拔土壤P组分存在显著差异(表 4)。速效P含量随海拔降低而增加, 且其含量在海拔2000、1800 m和1600 m显著低于海拔1400 m和1200 m。相反, 易分解P、中等易分解P、难利用P和总P含量随海拔降低而减少, 且在海拔2000、1800 m和1600 m显著高于海拔1400 m和1200 m。
| 2000 m | 1800 m | 1600 m | 1400 m | 1200 m | P | |
| 速效P Soluble phosphorus | 4.60±0.46b | 4.00±1.05b | 4.50±0.60b | 7.20±1.08a | 7.00±1.35a | 0.005 |
| 易分解P Labile phosphorus | 34.20±2.70a | 25.80±4.96b | 29.25±2.25a | 14.10±3.41c | 16.20±3.25c | <0.001 |
| 中等易分解P Moderately labile phosphorus |
129.30±17.76a | 83.70±17.57b | 99.45±13.05b | 45.90±8.59)c | 53.40±8.51c | <0.001 |
| 难利用P Occluded phosphorus |
348.17±7.27a | 184.63±0.55b | 172.00±19.67b | 121.50±11.01d | 147.80±5.80c | <0.001 |
| 总P Total phosphorus | 516.27±27.75a | 298.13±23.31b | 305.23±1.96b | 188.70±23.17c | 224.40±16.83c | <0.001 |
| 表中数据为平均值±标准差, 同一行不同小写字母表示不同海拔间差异显著(P < 0.05) | ||||||
以土壤P组分作为响应变量, 土壤基本性质、酸性磷酸酶活性和微生物生物量作为环境解释因子, 对不同海拔土壤P组分进行冗余分析(图 3)。分析显示, 不同海拔样点按照一定的规律进行聚类, 其中较高海拔(2000、1800 m和1600 m)和较低海拔(1400 m和1200 m)分别处于第1轴的正负两侧, 说明海拔梯度下环境解释因子的变化影响了P组分的变化。环境因子解释了土壤P组分变异的92.19%, 第1轴和第2轴分别解释了变量的86.51%和5.68%。其中, MBP解释了土壤P组分变化的60.5%, 且与易分解P、中等易分解P呈显著正相关, 与速效P呈显著负相关。MBN解释了P组分变化的24.1%, 且与难利用P、TP呈显著正相关。

|
| 图 3 不同海拔黄山松林土壤磷组分变化的冗余分析 Fig. 3 Redundancy analysis on changes of soil phosphorus fractions in Pinus taiwanensis forest at different elevations ST: 土壤温度Soil temperature; pH: 酸碱度; SOC: 总有机碳Soil organic carbon; TN: 总氮Total nitrogen; ACP: 酸性磷酸单酯酶acid phosphomonoesterase; PD: 酸性磷酸双酯酶acid phosphodiesterase; MBP: 微生物生物量磷Microbial biomass phosphorus; MBN: 微生物生物量氮Microbial biomass nitrogen; TP: 总磷Total phosphorus |
土壤SOC、TN在维持陆地生态系统土壤P有效性中具有重要作用[27], 其分解和矿化受到温度、水分等因素的调控[28]。本研究中, 土壤SOC、TN含量随海拔降低而呈减少趋势, 这与De等[11]和Chang等[28]的研究结果一致, 主要是由于温度升高和水分减少, 引起土壤SOC、TN矿化速率增加, 导致SOC、TN含量减少。然而, 在本研究中, 随海拔降低, 黄山松林郁闭度、林分密度、平均胸径和树高具有增加的趋势, 这表明其凋落物归还数量也会增加。尽管如此, 郑媛等[29]的研究发现本研究区低海拔的凋落物分解速率更高, 这意味着低海拔处具有更高的C周转速率, 即C输入与输出增加, 这可能是受到温度的调控。DOC、DON主要来源于土壤有机质、凋落物和细根残体的分解等[30], 本研究低海拔处相对较高的DOC、DON含量也进一步表明低海拔由于较高的温度加速其C周转。有研究表明, 亚热带森林越低的土壤pH使有机质的溶解性越高, 从而有利于增加DOC、DON含量[31]。相比较高海拔地区, 本研究中较低海拔地区温度较高和土壤pH较低, 有利于土壤有机质、凋落物等的分解和矿化, 从而提高DOC、DON含量。
3.2 海拔梯度变化对土壤微生物生物量和酸性磷酸酶活性的影响微生物生物量(SMB)是土壤养分循环的重要组成部分, 参与养分元素C、N、P的矿化和固定[9]。He等[32]关于全球meta分析的研究表明, 热带和亚热带地区土壤SMB随海拔降低而减少, 且SMB与SOC呈正相关, 并指出SOC是预测全球SMB海拔梯度模式的重要指标。本研究中, 随海拔降低, MBC、MBN、MBP含量呈减少趋势, 且MBC、MBN、MBP与SOC和TN呈显著正相关, 表明较低海拔土壤SOC、TN含量较少是导致其减少的重要原因。微生物生物量化学计量比是评估土壤生态系统营养限制的重要指标[33]。本研究中, MBP在较低海拔显著低于较高海拔, 而MBC/MBP和MBN/MBP在较低海拔显著高于较高海拔, 这表明较低海拔土壤微生物中的P素相对缺乏, 微生物对P的需求较高[34]。因此, 较低海拔土壤微生物通过提高ACP、PD活性以满足其对P素的需求(图 2)。
磷酸酶在调控和维持土壤P的周转中起着关键作用[35]。由于本地区土壤类型以酸性土壤为主, pH普遍较低, 因此, 占主导地位的磷酸酶主要包括酸性磷酸单酯酶(ACP)和酸性磷酸双酯酶(PD)[35]。底物有效性和生物体对磷的需求是影响磷酸酶生成的关键因素[35-36]。本研究中ACP、PD与DOC、DON呈显著正相关, 较低海拔处DOC、DON含量较高, 为微生物提供更多的有效养分, 有利于生物体合成更多ACP、PD。此外, 温度也是影响酶活性的重要因素[37], 本研究较低海拔处温度较高, 养分的周转速率较高, 生物对P的需求增加, 微生物通过提高APC、PD活性以获取P。有研究表明, 微生物通过优先分配能量和分泌胞外酶以获取最限制其生长的资源, 会导致SMB的降低[38]。酶活性/MBC的比值可以用于解释微生物对酶生产的能量分配[39]。本研究中, 较低海拔处ACP/MBC、PD/MBC较高, 这主要是由于该地区微生物对P的需求较高。因此, 微生物通过自身能量分配策略, 将更多的能量用于合成ACP、PD, 以获取有效P, 而降低对SMB的投入。
3.3 海拔梯度变化对土壤磷组分的影响及其调控因素Hou等[40]基于全球范围的研究表明, 气候可以通过影响主要土壤性质(如土壤pH、SOC含量等)、植物和微生物活性等因素进而影响土壤P组分和P有效性。本研究中, 土壤TP含量随海拔降低呈减少的趋势, 且在较低海拔显著低于较高海拔, 与Hou等[40]和Zhou等[13]的研究结果类似。土壤TP和P有效性通常随温度的增加而降低, 这主要由于温度的增加提高了土壤风化程度以及原生矿物磷、有机磷和有机碳的矿化[13, 40]。本研究中, 较低海拔处温度较高, 土壤pH较低, 土壤P更容易被风化损耗, 从而使TP含量减少[41]。另一方面, 较低海拔处SOC、TN的减少表明SOC的矿化可能增强, 从而不利于土壤TP含量和P有效性的提高[27]。此外, 在本研究中, 随海拔降低, 黄山松林郁闭度、林分密度、平均胸径和树高具有增加的趋势(表 1), 较高的植被生物量可能会导致凋落物量增加。然而, 这与土壤P含量的海拔变化趋势并不一致, 因此凋落物数量可能不是影响土壤中P含量的主要因素。本团队先前的研究[29]表明, 黄山松叶片对于P的重吸收随海拔降低而减少, 高海拔黄山松的重吸收能力高于低海拔, 因此凋落物P含量也并不是影响土壤中P含量的主要因素。综合以上结果表明, 植被凋落物并非是影响土壤P含量的主要因素。土壤微生物通过分泌酸性磷酸酶等物质促进P的矿化, 加速无机磷的分解和活化, 从而提高土壤P有效性[42]。速效P作为可以被植物直接吸收利用的无机磷[3], 本研究中速效P含量在较低海拔显著高于较高海拔, 这与热带森林[43]和寒带苔原地区[12]的研究结果相似, 而易分解P、中等易分解P、难利用P则相反。这是由于较低海拔微生物分泌较多的ACP、PD, 使更多较难分解的P组分矿化为无机磷[7], 从而增加速效P含量, 而易分解P、中等易分解P、难利用P含量减少。此外, 易分解P、中等易分解P含量在海拔较低处显著降低, 很可能还受到MBP减少的影响。由于MBP是土壤潜在有效P库的重要组成部分[44], 因此易分解P、中等易分解P表现出与MBP相同的趋势。
本研究结果表明, MBP和MBN是影响土壤P组分的主要因素。Bing等[45]研究表明, MBP在土壤P循环和植物P供应中发挥重要作用, 当土壤P素供应不足时, MBP的周转能够补充土壤溶液中的P, 并有利于植物对P素的吸收和利用。本研究中, MBP在较低海拔处显著降低, 与易分解P、中等易分解P呈显著正相关, 表明MBP作为土壤P的活性形式, 固持在微生物体内, 是土壤有效P潜在的供给源和储备库[46], 较低海拔处较高的温度促进了微生物对土壤P的转化, 提高速效P含量。然而, 从长期来看, 可能会导致P库的储备不足, 从而对较低海拔地区土壤P素养分的可持续供应产生不利影响。此外, 本研究中MBN含量的海拔变化呈现出与MBP相同的趋势, 且与TP、难分解P具有显著正相关关系, 表明MBN在P组分的转化中具有重要作用。由于微生物合成胞外酶时对N的需求较高, MBN含量会影响胞外酶的合成[47]。微生物通过降低MBN含量和增加N有效性, 能够促进土壤酸性磷酸酶的合成[48]。因此, 较低海拔土壤微生物对P的需求较高, MBN可能通过ACP、PD的合成间接影响土壤P组分的转化, 而MBP可直接参与P组分的转化。此外, MBC作为SMB的重要组成部分, 其在海拔梯度上也表现出与MBN、MBP相同的趋势。综上所述, 随海拔降低, 黄山松林土壤微生物通过提高ACP、PD活性和降低SMB含量的能量分配策略, 促进土壤P组分的转化, 从而提高土壤P有效性(图 4)。

|
| 图 4 海拔梯度变化对黄山松林土壤磷组分和有效性的影响的概念图(红色“↑”表示增加;蓝色“↓”表示降低) Fig. 4 Conceptual diagram of the effects of elevations on soil phosphorus fractions and availability in Pinus taiwanensis forest |
随海拔降低, 速效P含量显著增加, 而易分解P、中等易分解P、难利用P和TP含量显著减少。MBP和MBN是影响土壤P组分和有效性的主要因素。在低海拔地区, 微生物通过提高ACP、PD活性和降低SMB含量的能量分配策略, 促进更多较难分解P组分的转化, 从而提高速效P含量, 增加P有效性, 以满足微生物对P的需求。因此, 在低海拔地区, 微生物通过能量分配策略获取更多有效P, 可能有助于提高武夷山黄山松林土壤速效P的供给, 但从长远来看, 可能会增加P矿化速率和P损耗率, 导致P库的储备不足, 从而不利于土壤P素养分的可持续供应。此外, 微生物及其胞外酶的产生在P组分的转化中具有重要的作用, 而酸性磷酸酶的主要来源是真菌[49], 真菌群落对海拔梯度变化的响应将会影响土壤P组分转化的过程[50]。因此, 未来还需进一步探究真菌群落结构与功能之间的联系, 以了解微生物在调节山地生态系统土壤P组分和有效性的关键过程和机制。
| [1] |
Hou E Q, Luo Y Q, Kuang Y W, Chen C R, Lu X K, Jiang L F, Luo X Z, Wen D Z. Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems. Nature Communications, 2020, 11: 637. DOI:10.1038/s41467-020-14492-w |
| [2] |
Huang L M, Jia X X, Zhang G L, Shao M A. Soil organic phosphorus transformation during ecosystem development: a review. Plant and Soil, 2017, 417(1/2): 17-42. DOI:10.1007/s11104-017-3240-y |
| [3] |
Hou E Q, Chen C R, Kuang Y W, Zhang Y G, Heenan M, Wen D Z. A structural equation model analysis of phosphorus transformations in global unfertilized and uncultivated soils. Global Biogeochemical Cycles, 2016, 30(9): 1300-1309. DOI:10.1002/2016GB005371 |
| [4] |
Lie Z Y, Lin W, Huang W J, Fang X, Huang C M, Wu T, Chu G W, Liu S Z, Meng Z, Zhou G Y, Liu J X. Warming changes soil N and P supplies in model tropical forests. Biology and Fertility of Soils, 2019, 55(7): 751-763. DOI:10.1007/s00374-019-01382-7 |
| [5] |
Sardans J, Peñuelas J, Estiarte M. Warming and drought alter soil phosphatase activity and soil P availability in a Mediterranean shrubland. Plant and Soil, 2006, 289(1/2): 227-238. DOI:10.1007/s11104-006-9131-2 |
| [6] |
Hou E Q, Wen D Z, Kuang Y W, Cong J, Chen C R, He X J, Heenan M, Lu H, Zhang Y G. Soil pH predominantly controls the forms of organic phosphorus in topsoils under natural broadleaved forests along a 2500 km latitudinal gradient. Geoderma, 2018, 315: 65-74. DOI:10.1016/j.geoderma.2017.11.041 |
| [7] |
Fan Y X, Lin F, Yang L M, Zhong X J, Wang M H, Zhou J C, Chen Y M, Yang Y S. Decreased soil organic P fraction associated with ectomycorrhizal fungal activity to meet increased P demand under N application in a subtropical forest ecosystem. Biology and Fertility of Soils, 2018, 54(1): 149-161. DOI:10.1007/s00374-017-1251-8 |
| [8] |
Reed S C, Yang X J, Thornton P E. Incorporating phosphorus cycling into global modeling efforts: a worthwhile, tractable endeavor. New Phytologist, 2015, 208(2): 324-329. DOI:10.1111/nph.13521 |
| [9] |
He X J, Hou E Q, Liu Y, Wen D Z. Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China. Scientific Reports, 2016, 6: 24261. DOI:10.1038/srep24261 |
| [10] |
曾晓敏, 范跃新, 林开淼, 赵盼盼, 袁萍, 周嘉聪, 张秋芳, 程蕾, 徐建国, 陈岳民. 亚热带不同海拔黄山松林土壤磷组分及微生物特征. 生态学报, 2018, 38(18): 6570-6579. |
| [11] |
De F M, Cardelli V, Massaccesi L, Bol R, Willbold S, Cocco S, Corti G, Agnelli A. Effect of beech (Fagus sylvatica L.) rhizosphere on phosphorous availability in soils at different altitudes (Central Italy). Geoderma, 2016, 276: 53-63. DOI:10.1016/j.geoderma.2016.04.028 |
| [12] |
Vincent A G, Sundqvist M K, Wardle D A, Giesler R. Bioavailable soil phosphorus decreases with increasing elevation in a subarctic tundra landscape. PLoS One, 2014, 9(3): e92942. DOI:10.1371/journal.pone.0092942 |
| [13] |
Zhou J, Wu Y H, Bing H J, Yang Z J, Wang J P, Sun H Y, Sun S Q, Luo J. Variations in soil phosphorus biogeochemistry across six vegetation types along an altitudinal gradient in SW China. Catena, 2016, 142: 102-111. DOI:10.1016/j.catena.2016.03.004 |
| [14] |
聂阳意, 王海华, 李晓杰, 任寅榜, 金昌善, 徐自坤, 吕茂奎, 谢锦升. 武夷山低海拔和高海拔森林土壤有机碳的矿化特征. 应用生态学报, 2018, 29(3): 748-756. |
| [15] |
封磊, 洪伟, 吴承祯, 宋萍, 兰思仁. 武夷山黄山松群落物种多样性与种群空间格局的研究. 中国生态农业学报, 2004, 12(3): 16-18. |
| [16] |
周永姣, 程林, 王满堂, 朱国洁, 钟全林, 郭炳桥, 程栋梁. 武夷山不同海拔黄山松细根性状季节变化. 生态学报, 2019, 39(12): 4530-4539. |
| [17] |
Huang Z Q, Wan X H, He Z M, Yu Z P, Wang M H, Hu Z H, Yang Y S. Soil microbial biomass, community composition and soil nitrogen cycling in relation to tree species in subtropical China. Soil Biology and Biochemistry, 2013, 62: 68-75. DOI:10.1016/j.soilbio.2013.03.008 |
| [18] |
Jones D L, Willett V B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biology and Biochemistry, 2006, 38(5): 991-999. DOI:10.1016/j.soilbio.2005.08.012 |
| [19] |
Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703-707. DOI:10.1016/0038-0717(87)90052-6 |
| [20] |
Juma N G, Tabatabai M A. Comparison of kinetic and thermodynamic parameters of phosphomonoesterases of soils and of corn and soybean roots. Soil Biology and Biochemistry, 1988, 20(4): 533-539. DOI:10.1016/0038-0717(88)90069-7 |
| [21] |
曾晓敏, 范跃新, 林开淼, 袁萍, 赵盼盼, 陈怡然, 徐建国, 陈岳民. 亚热带不同植被类型土壤磷组分特征及其影响因素. 应用生态学报, 2018, 29(7): 2156-2162. |
| [22] |
Murphy J, Riley J P. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta, 1962, 27: 31-36. DOI:10.1016/S0003-2670(00)88444-5 |
| [23] |
Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315. DOI:10.1016/S0038-0717(02)00074-3 |
| [24] |
Tian J H, Wei K, Condron L M, Chen Z H, Xu Z W, Chen L J. Impact of land use and nutrient addition on phosphatase activities and their relationships with organic phosphorus turnover in semi-arid grassland soils. Biology and Fertility of Soils, 2016, 52(5): 675-683. DOI:10.1007/s00374-016-1110-z |
| [25] |
Hedley M J, Stewart J W B, Chauhan B S. Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Science Society of America Journal, 1982, 46(5): 970-976. DOI:10.2136/sssaj1982.03615995004600050017x |
| [26] |
Fan Y X, Zhong X J, Lin F, Liu C C, Yang L M, Wang M H, Chen G S, Chen Y, Yang Y S. Responses of soil phosphorus fractions after nitrogen addition in a subtropical forest ecosystem: insights from decreased Fe and Al oxides and increased plant roots. Geoderma, 2019, 337: 246-255. DOI:10.1016/j.geoderma.2018.09.028 |
| [27] |
Hou E Q, Chen C R, Wen D Z, Liu X. Relationships of phosphorus fractions to organic carbon content in surface soils in mature subtropical forests, Dinghushan, China. Soil Research, 2014, 52(1): 55-63. DOI:10.1071/SR13204 |
| [28] |
Chang E H, Chen T H, Tian G L, Chiu C Y. The effect of altitudinal gradient on soil microbial community activity and structure in moso bamboo plantations. Applied Soil Ecology, 2016, 98: 213-220. DOI:10.1016/j.apsoil.2015.10.018 |
| [29] |
郑媛. 武夷山黄山松凋落物分解的研究[D]. 福州: 福建师范大学, 2017.
|
| [30] |
赵盼盼, 周嘉聪, 林开淼, 林伟盛, 袁萍, 曾晓敏, 苏莹, 徐建国, 陈岳民, 杨玉盛. 不同海拔对福建戴云山黄山松林土壤微生物生物量和土壤酶活性的影响. 生态学报, 2019, 39(8): 2676-2686. |
| [31] |
肖好燕, 刘宝, 余再鹏, 万晓华, 桑昌鹏, 周富伟, 黄志群. 亚热带典型林分对表层和深层土壤可溶性有机碳、氮的影响. 应用生态学报, 2016, 27(4): 1031-1038. |
| [32] |
He X J, Hou E Q, Veen G F, Ellwood M D F, Dijkstra P, Sui X H, Zhang S, Wen D Z, Chu C J. Soil microbial biomass increases along elevational gradients in the tropics and subtropics but not elsewhere. Global Ecology and Biogeography, 2020, 29(2): 345-354. DOI:10.1111/geb.13017 |
| [33] |
曾泉鑫, 曾晓敏, 林开淼, 张秋芳, 程蕾, 周嘉聪, 林巧玉, 陈岳民, 徐建国. 亚热带毛竹林土壤磷组分和微生物对施氮的响应. 应用生态学报, 2020, 31(3): 753-760. |
| [34] |
Machmuller M B, Mohan J E, Minucci J M, Phillips C A, Wurzburger N. Season, but not experimental warming, affects the activity and temperature sensitivity of extracellular enzymes. Biogeochemistry, 2016, 131(3): 255-265. DOI:10.1007/s10533-016-0277-6 |
| [35] |
Hou E Q, Chen C R, Wen D Z, Liu X. Phosphatase activity in relation to key litter and soil properties in mature subtropical forests in China. Science of the Total Environment, 2015, 515-516: 83-91. DOI:10.1016/j.scitotenv.2015.02.044 |
| [36] |
Sinsabaugh R L, Shah J J F. Ecoenzymatic stoichiometry and ecological theory. Annual Review of Ecology, Evolution, and Systematics, 2012, 43(1): 313-343. DOI:10.1146/annurev-ecolsys-071112-124414 |
| [37] |
Meng C, Tian D S, Zeng H, Li Z L, Chen H Y H, Niu S L. Global meta-analysis on the responses of soil extracellular enzyme activities to warming. Science of the Total Environment, 2020, 705: 135992. DOI:10.1016/j.scitotenv.2019.135992 |
| [38] |
Čapek P, Starke R, Hofmockel K S, Bond-Lamberty B, Hess N. Apparent temperature sensitivity of soil respiration can result from temperature driven changes in microbial biomass. Soil Biology and Biochemistry, 2019, 135: 286-293. DOI:10.1016/j.soilbio.2019.05.016 |
| [39] |
Dong C C, Wang W, Liu H Y, Xu X T, Zeng H. Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: Evidence from soil extracellular enzyme stoichiometry. Ecological Indicators, 2019, 101: 453-464. DOI:10.1016/j.ecolind.2019.01.046 |
| [40] |
Hou E Q, Chen C R, Luo Y Q, Zhou G Y, Kuang Y W, Zhang Y G, Heenan M, Lu X K, Wen D Z. Effects of climate on soil phosphorus cycle and availability in natural terrestrial ecosystems. Global Change Biology, 2018, 24(8): 3344-3356. DOI:10.1111/gcb.14093 |
| [41] |
Dixon J L, Chadwick O A, Vitousek P M. Climate-driven thresholds for chemical weathering in postglacial soils of New Zealand. Journal of Geophysical Research: Earth Surface, 2016, 121(9): 1619-1634. DOI:10.1002/2016JF003864 |
| [42] |
Richardson A E, Simpson R J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiology, 2011, 156(3): 989-996. DOI:10.1104/pp.111.175448 |
| [43] |
Soethe N, Lehmann J, Engels C. Nutrient availability at different altitudes in a tropical montane forest in Ecuador. Journal of Tropical Ecology, 2008, 24(4): 397-406. DOI:10.1017/S026646740800504X |
| [44] |
贝昭贤, 张秋芳, 郑蔚, 杨柳明, 陈岳民, 杨玉盛. 模拟增温对中亚热带杉木人工林土壤磷有效性的影响. 生态学报, 2018, 38(3): 1106-1113. |
| [45] |
Bing H J, Wu Y H, Zhou J, Sun H Y, Luo J, Wang J P, Yu D. Stoichiometric variation of carbon, nitrogen, and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau. Journal of Soils and Sediments, 2016, 16(2): 405-416. DOI:10.1007/s11368-015-1200-9 |
| [46] |
Achat D L, Morel C, Bakker M R, Augusto L, Pellerin S, Gallet-Budynek A, Gonzalez M. Assessing turnover of microbial biomass phosphorus: combination of an isotopic dilution method with a mass balance model. Soil Biology and Biochemistry, 2010, 42: 2231-2240. DOI:10.1016/j.soilbio.2010.08.023 |
| [47] |
Malik A A, Puissant J, Goodall T, Allison S D, Griffiths R I. Soil microbial communities with greater investment in resource acquisition have lower growth yield. Soil Biology and Biochemistry, 2019, 132: 36-39. DOI:10.1016/j.soilbio.2019.01.025 |
| [48] |
Nottingham A T, Turner B L, Whitaker J, Ostle N J, McNamara N P, Bardgett R D, Salinas N, Meir P. Soil microbial nutrient constraints along a tropical forest elevation gradient: a belowground test of a biogeochemical paradigm. Biogeosciences,, 2015, 12(20): 6071-6083. DOI:10.5194/bg-12-6071-2015 |
| [49] |
Turner B L, Haygarth P M. Phosphatase activity in temperate pasture soils: potential regulation of labile organic phosphorus turnover by phosphodiesterase activity. Science of the Total Environment, 2005, 344(1/3): 27-36. |
| [50] |
Cui Y X, Bing H J, Fang L C, Wu Y H, Yu J L, Shen G T, Jiang M, Wang X, Zhang X C. Diversity patterns of the rhizosphere and bulk soil microbial communities along an altitudinal gradient in an alpine ecosystem of the eastern Tibetan Plateau. Geoderma, 2019, 338: 118-127. DOI:10.1016/j.geoderma.2018.11.047 |