 2021, Vol. 41
2021, Vol. 41文章信息
- 刘凤, 曾永年
- LIU Feng, ZENG Yongnian
- 2000—2015年青海高原植被碳源/汇时空格局及变化
- Analysis of the spatio-temporal variation of vegetation carbon source/sink in Qinghai Plateau from 2000—2015
- 生态学报. 2021, 41(14): 5792-5803
- Acta Ecologica Sinica. 2021, 41(14): 5792-5803
- http://dx.doi.org/10.5846/stxb201909302057
-
文章历史
- 收稿日期: 2019-09-30
- 网络出版日期: 2021-05-13

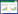




2. 中南大学地球科学与信息物理学院, 长沙 410083
2. School of Geosciences and Info-physics, Central South University, Changsha 410083, China
碳循环能够综合地反映生态系统对气候变化的响应及人类活动的影响[1-3], 因而成为全球气候变化研究的重要内容之一[4-6], 也是近年来全球碳计划、全球变化与陆地生态系统响应计划等一系列国际全球变化研究核心计划的焦点内容。陆地植被碳循环是碳循环的重要组成部分, 目前陆地植被碳循环的相关研究从对GPP(植被总生产力, Gross Primary Productivity)、NPP(植被净初级生产力, Net Primary Productivity)的研究, 进一步发展到对NEP(植被净生态系统生产力, Net Ecosystem Productivity)的估算、区域碳收支的确定、碳源/汇的估算以及定量的评估气候因素变化、人类活动对植被碳储量的影响[7-9]。NEP表示NPP与RH(土壤异养呼吸, Heterotrophic Respiration)之间的差值, 是植被生态系统碳的净吸收或净储存, 可以定量描述植被生态系统碳源/汇的能力。当NEP>0时, 表示生态系统发挥碳汇功能, 反之则为碳源[10-11]。因此, NEP作为表征植被活动的关键参量, 对其准确估算不仅有助于衡量植被生态系统健康程度, 更有助于定量分析区域植被生态系统的固碳状况与潜力。
青海高原平均海拔在3000 m以上, 是全球海拔最高的独特自然地理单元——青藏高原的重要组成部分, 是我国以及南亚、东南亚地区的“江河源”、“生态源”, 是区域乃至全球气候变化的“启动器”和“调节器”, 也是我国和全球生物多样性保护地和生态安全屏障[12]。因此, 青海高原植被碳源/汇时空格局及变化的研究, 对深刻认识和理解青海高原植被生态系统碳汇功能及对全球变化的响应与影响具有重要的科学意义, 对青海高原植被生态系统碳循环的合理调控和生态保护工程的实施具有重要的科学参考价值。目前, 已有较多的研究对青海高原相关地区植被覆盖度变化及植被NPP进行了分析[13-16], 对我国、青藏高原、三江源的陆地生态系统碳收支进行了分析[17-22]。然而, 目前对青海高原植被NEP及其碳源/汇时空格局研究的尺度尚不够精细, 研究的时间序列有限从而影响到全面、精细地认识青海高原植被碳源/汇的时空分布差异及其影响因素。因此, 采用适用于青海高原的NEP估算模型, 利用较高时空分辨率的MODIS数据, 定量估算并分析青海高原植被碳源/汇的时空格局及变化具有重要的理论与实践意义。
本研究利用高分辨率遥感数据、气象数据, 采用改进的CASA模型(基于过程的遥感模型, Carnegie Ames Stanford Approach Model)、土壤呼吸模型, 估算青海高原不同生态区植被NEP, 分析NEP的时空分布格局与变化, 探讨不同生态区碳源/汇分布差异、年际变化及NEP对气候因素变化的响应, 以期深入揭示青海高原碳源/汇时空格局及其区域差异性。
1 研究区及数据 1.1 研究区概况青海高原位居我国西部, 青藏高原东北部, 地处东部季风区、西北干旱区和青藏高原区三大地理区域的交汇处, 是长江、黄河、澜沧江的发源地[23]。地理位置介于东经89°25′—103°04′, 北纬31°40′—39°19′之间, 土地总面积72.23万km2, 地势西高东低, 南北高中间低, 平均海拔3000 m以上, 地形复杂, 地貌多样。气候属大陆性高原气候, 冬季漫长寒冷、夏季短暂凉爽[24], 年降水量为15—750 mm, 降水量时空分布不均, 从西北到东南渐增。年均温为-6—9℃, 气温的垂直变化明显。主要植被类型有常绿针叶林、落叶阔叶林、高寒灌丛、高寒草甸、高寒草原、高寒荒漠、温性草原等, 植被水平与垂直地带性分异显著。青海高原生态区包括5个一级生态分区和14个二级生态分区:Ⅰ—1阿尔金山荒漠生态区;Ⅰ—2昆仑山东段高寒荒漠草原生态区;Ⅱ—1柴达木盆地荒漠生态区;Ⅱ—2柴达木盆地东北部山地高寒荒漠草原生态区;Ⅲ—1西祁连山高寒荒漠草原生态区;Ⅲ—2东祁连山云杉林——高寒草甸生态区;Ⅲ—3青海湖湿地及上游高寒草甸生态区;Ⅲ—4湟水谷地农业生态区;Ⅳ北羌塘高寒荒漠草原生态区;Ⅴ—1长江源高寒草甸草原生态区;Ⅴ—2澜沧江源高寒草甸草原生态区;Ⅴ—3黄河源高寒草甸草原生态区;Ⅴ—4共和盆地高寒草原生态区;Ⅴ—5海东—甘南高寒草甸草原生态区(图 1)。

|
| 图 1 青海高原生态区划及DEM示意图 Fig. 1 Ecological regionalization and DEM schematic of Qinghai Plateau Ⅰ—1阿尔金山荒漠生态区;Ⅰ—2昆仑山东段高寒荒漠草原生态区;Ⅱ—1柴达木盆地荒漠生态区;Ⅱ—2柴达木盆地东北部山地高寒荒漠草原生态区;Ⅲ—1西祁连山高寒荒漠草原生态区;Ⅲ—2东祁连山云杉林——高寒草甸生态区;Ⅲ—3青海湖湿地及上游高寒草甸生态区;Ⅲ—4湟水谷地农业生态区;Ⅳ北羌塘高寒荒漠草原生态区;Ⅴ—1长江源高寒草甸草原生态区;Ⅴ—2澜沧江源高寒草甸草原生态区;Ⅴ—3黄河源高寒草甸草原生态区;Ⅴ—4共和盆地高寒草原生态区;Ⅴ—5海东—甘南高寒草甸草原生态区 |
数据包括遥感影像数据、气象数据、地形数据、植被覆盖数据及其他数据。其中, 遥感数据来源于NASA官网(http://edcimswww.cr.usgs.gov/pub/imswelcome/)提供的EOS/MODIS产品2000—2015年青海高原365期MOD13Q1—NDVI(归一化差值植被指数, Normalized Difference Vegetation Index)数据集, 数据空间分辨率为250 m×250 m, 时间分辨率为16 d。采用MRT软件与ArcGIS软件, 完成影像的拼接、裁剪、最大值合成等预处理, 并对NDVI数据作滤波处理, 剔除异常值, 获得2000—2015年研究区逐月NDVI序列数据。
气象数据来源于中国气象数据网(http://cdc.nmic.cn/)提供的2000—2015年青海高原及其周围59个气象站点的月总降水量、月平均气温观测数据, 以及8个太阳辐射站点的月太阳总辐射量数据。为提高模型中气象数据的精度, 利用覆盖研究区的DEM数据及经纬度数据, 在SPSS和ArcGIS软件中, 采用多元线性回归法[25]完成气象数据的空间插值, 该方法引入多个影响因子(如经度、纬度、高程等)作为自变量, 结合已知气象站点观测值, 采用线性回归的方法, 最终获得与NDVI数据一致空间分辨率、一致投影信息的气象要素栅格数据集。
地形数据源自地理空间数据云(http://www.gscloud.cn/)下载的30 m分辨率DEM数据;植被覆盖数据来源于地球系统科学数据共享网(http://www.geodata.cn/)中, 中国地区2005年1:25万土地覆盖遥感调查与监测数据, 该数据集的分类精度为80%—90%, 随后为了分析不同植被类型NPP, 依据分类标准将青海高原主要植被类型合并为草地、森林、农田, 3个一级土地覆盖类型。青海高原行政区划数据源自国家基础地理信息中心(http://ngcc.sbsm.gov.cn/)。生态区边界数据采用中国陆地生态系统数据库[26](http://www.ecosystem.csdb.cn/)中青海高原生态区划数据, 并根据不同生态区的生态系统类型、地理特征等自然条件, 分为5个一级生态分区和14个二级生态分区。
2 模型与方法 2.1 NEP估算模型植被NEP是衡量区域上植被碳源、碳汇的重要指标, 在不考虑其他自然和人为因素的影响下, NEP等于植被NPP与土壤微生物呼吸消耗碳量的差值[27], 计算公式如下:
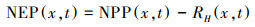
|
(1) |
式中, NEP(x, t)为像元x在第t月的植被净生态系统生产力, 单位为gC/m2, NPP(x, t)为像元x在第t月的植被净初级生产力(gC/m2), RH(x, t)表示像元x在第t月的土壤微生物呼吸量(gC/m2)。当NEP>0, 说明植被固定的碳多于土壤呼吸排放碳, 植被固碳体现为碳汇作用, 反之, 则为碳源作用。
采用朱文泉等[28-29]改进的CASA模型实现对青海高原2000—2015年植被NPP的估算, 模型中通过植被吸收的光合有效辐射和光能利用率, 两个驱动因子计算NPP, 公式如下:
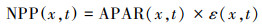
|
(2) |
式中, APAR(x, t)表示像元x在t月吸收的光合有效辐射(MJ/m2);ε(x, t)表示像元x在t月的实际光能利用率(gC/MJ), 公式中相关参数的计算参见文献[28-31]。
土壤异养呼吸消耗RH的估算, 采用裴永志等[32]建立的模型, 即:
|
(3) |
式中, T(x, t)表示像元x在第t月的平均气温(℃);R(x, t)表示像元x在第t月的月平均降水量(mm)。
2.2 时空变化分析方法 2.2.1 NEP年际变化率采用一元线性回归法分析植被NEP的年际波动规律与动态变化[33], 计算研究区逐像元多年回归趋势线斜率, 用于反映植被NEP年际变化率。计算公式如下:
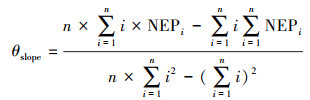
|
(4) |
式中, θslope表示趋势斜率, n表示估算年数, NEP表示第i年的植被NEP, 利用NEP值与时间序列的相关关系表示NEP年际变化的显著性, θslope>0表示增加, 反之减少。
2.2.2 变异系数(稳定性分析)2000—2015年青海高原植被NEP的稳定性用基于像元的变异系数[34](式5)衡量,
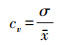
|
(5) |
式中, cv为变异系数, σ为标准差, x为平均值。变异系数反映数据的离散程度, c值越小, 表示数据分布越集中, 数据随时间波动小, 具有较高的稳定性;反之, 则表示数据随时间序列波动大, 稳定性低。
2.2.3 NEP与气候因素相关性采用基于像元的空间分析法, 分析植被NEP与气候因素(降水量、气温)的相关性, 确定气候因素与NEP的相关系数(式(6)), 以及偏相关系数(式(7))。
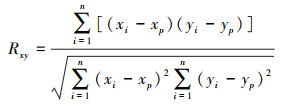
|
(6) |
式中, Rxy表示x、y变量间的相关关系;xi为第i年的NEP;yi表示第i年变量y的值;xp为多年NEP的平均值;yp为变量y多年平均值;n为样本数。
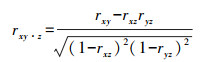
|
(7) |
式中, rxy, ryz, rxz分别表示变量x与y, y与z, 及x与z的相关系数, rxy·z指变量z固定后, 变量x与y之间的偏相关系数。
采用T检验法[35]对各相关系数、偏相关系数进行显著性检验, 显著性水平取α=0.05。依据显著性水平将各系数划分为显著正相关(R>0, P < 0.05)、不显著正相关(R>0, P≥0.05)、显著负相关(R < 0, P < 0.05)和不显著负相关(R < 0, P≥0.05), 并将研究区按不同显著性水平划分为4种不同的区域。
2.2.4 精度评价在分析研究区相关文献资料的基础上, 统计野外站点植被NPP实测值、地上净初级生产量、野外样地生物量实测数据[36-40]。为利用地上生物量实测数据获得植被NPP值, 首先, 确定地上生物量与地下生物量的比率关系(即根冠比, R/S), 本文选用的R/S为5.8[40], 根据根冠比计算出相应植被的地下生物量, 并计算得到总生物量, 再根据生物量与NPP的换算比率将生物量转换成碳单位, 本文采用0.45[41]的比率将生物量转换成碳单位, 得到统计NPP实测数据;其次, 对于实测数据中有气象站点位置信息的, 提取对应位置、时间的RH计算结果, 对于仅有所在县级单位的实测数据, 提取对应年份、月份, 同一县级单元RH计算结果平均值。最后, 利用NPP、RH值计算获得NEP实测数据, 并采用与RH同样的提取方式, 提取本文模拟植被NEP值, 利用SPSS软件分析模拟植被NEP与实测植被NEP的相关性, 并检验其显著性。同时, 整理统计其他研究模拟的NEP结果, 提取与本研究对应时间、相同植被类型的NEP值进行对比分析与评价分析。
3 结果与分析 3.1 NEP估算精度评价根据2000—2015年青海高原野外气象站点植被NPP、植被净初级生产量、植被地上生物量观测数据换算获得的植被NPP实测值, 结合土壤呼吸消耗RH计算结果, 获得植被NEP实测数据, 再与本文估算植被NEP结果进行相关性分析、显著性检验, 结果表明(图 2), 本文模拟植被NEP结果与统计实测数据的相关系数为0.87(P < 0.01, R2=0.759)(n=27), 模拟值与实测值呈显著相关, 说明模型估算植被NEP结果的可靠性。

|
| 图 2 植被净生态系统生产力(NEP) 模拟值与实测值对比 Fig. 2 Comparison of Net Ecosystem Productivity (NEP) simulated value and measured value |
2000—2015年, 青海高原植被NEP空间分布格局呈东高西低、南高北低, 由西北向东南逐步增加趋势, 青海高原不同生态区植被NEP空间分布及其特征, 如图 3、图 4所示。

|
| 图 3 植被NEP平均值空间分布 Fig. 3 Spatial distribution of vegetation NEP mean value |

|
| 图 4 植被NEP—16年总值空间分布 Fig. 4 Spatial distribution of 16 years′ total value of NEP |
青海高原植被年NEP多年平均值为128.40 gC m-2a-1, 年平均固碳量约为51.69 TgC/a。其中, 碳汇区(NEP>0)面积约为39.56万km2, 约占植被总面积的73.11%, 年固碳量平均值为188.43 gC m-2a-1, 年平均碳汇量约为59.1 TgC/a;碳源区(NEP < 0)面积约为14.55万km2, 约占植被总面积的26.89%, 年碳排放量平均值为-61.70 gC m-2a-1, 年平均碳排放量为-7.39 TgC/a, 年平均净碳汇总量约为51.7 TgC/a(图 3)。
2000—2015年, 青海高原植被累积NEP平均值为1101.92 gC/m2。其中, 碳汇区平均累积NEP为3014.92 gC/m2, 总固碳量约为0.946 PgC;碳源区平均累积NEP约为-987.13 gC/m2, 总碳排放量为-0.119 PgC(图 4)。
青海高原各生态区植被NEP及碳汇/源分布的空间格局特征如下:
Ⅰ帕米尔—昆仑山—阿尔金山生态区
该生态区, 地处青海高原西北部, 植被分布稀疏, 区内降水量稀少, 气温偏高, 土地沙漠化、荒漠化趋势明显, 不利于植被NEP累积, 多数区域土壤呼吸消耗碳大于植被NPP累积量。因此, 该生态区整体上属于青海高原植被NEP低值区, 16年累积固碳总量约为8.631 TgC, 属弱碳汇区。
Ⅰ—1阿尔金山荒漠生态区, 该区地处柴达木盆地西北部, 荒漠化严重, 植被生态系统呈现为碳源。植被年NEP平均值为-5.31 gC m-2a-1, 年固碳量约为-0.02 TgC;16年植被累积NEP平均值为-49.51 gC m-2, 累积碳排放总量约为-0.329 TgC。
Ⅰ—2昆仑山东段高寒荒漠草原生态区, 该区地处柴达木盆地南缘的高寒荒漠草原区, 整体以碳汇效应为主, 但固碳能力较弱。植被年NEP平均值仅8.73 gC m-2a-1, 年固碳量为0.56 TgC;16年植被累积NEP平均值为139.65 gCm2, 累积固碳总量约为8.96 TgC。
Ⅱ柴达木盆地生态区
该生态区植被覆盖稀少, 土壤沙化、盐渍化严重, 气候干燥少雨, 属青海高原植被NEP的低值区, 全区16年累积固碳总量为-40.37 TgC, 属碳源区。
Ⅱ—1柴达木盆地荒漠生态区, 位于柴达木盆地中部, 天然牧草地呈现不同程度的退化, 土地沙漠化严重, 属碳源区。植被年NEP平均值为-20.57 gC m-2a-1, 年累积固碳量为-2.77 TgC;16年累积植被NEP平均值为-329.20 gCm2, 累积碳排放量约为-44.37 TgC。
Ⅱ—2柴达木盆地东北部山地高寒荒漠草原生态区, 地处柴达木盆地东北部, 植被年NEP平均值为6.95 gC m-2a-1, 年平均固碳量为0.25 TgC/a;16年累积植被NEP平均值为111.09 gCm2, 累积固碳总量约为4.003 TgC, 属弱碳汇区。
Ⅲ祁连山生态区
该生态区属青海高原草地、荒漠草原的重要分布区之一, 分布有丰富的森林、草地资源, 区内水热条件耦合作用良好[42], 年植被NEP平均值在36.52—232.17 gC m-2a-1之间, 16年累积固碳总量为0.198 PgC, 植被NEP较高, 以碳汇作用为主。
Ⅲ—1西祁连山高寒荒漠草原生态区, 位于祁连山地西北部, 该区草地和森林资源较为丰富, 草地有轻微退化现象, 植被年NEP平均值为36.52 gC m-2a-1, 年平均固碳量为0.576 TgC;16年累积植被NEP平均值为584.30 gCm2, 累积固碳总量约为9.275 TgC。
Ⅲ—2东祁连山云杉林——高寒草甸生态区, 地处祁连山地东部, 林草资源丰富, 植被NEP年平均值为185.31 gC m-2a-1, 年固碳量为3.85 TgC;16年累积植被NEP平均值为2964.25 gCm2, 累积固碳总量约为61.58 TgC, 该区为青海高原的碳汇区。
Ⅲ—3青海湖湿地及上游高寒草甸生态区, 草地、沼泽湿地、森林资源富足, 水热组合条件良好, 植被年NEP平均值为118.40 gC m-2a-1, 年平均固碳量为3.83 TgC, 16年累积植被NEP平均值为1893.5 gCm2, 累积固碳总量约为61.30 TgC。
Ⅲ—4湟水谷地农业生态区, 位于青海高原东部, 地处河湟谷地, 是青海高原重要的耕地分布区, 气候条件温暖湿润, 有利于植被生长, 植被年NEP平均值为232.17 gC m-2a-1, 年平均固碳量为4.13 TgC, 16年累积植被NEP平均值为3711.79 gCm2, 累积固碳总量约为66.08 TgC。
Ⅳ北羌塘高寒荒漠草原生态区
该生态区地处青南高原西北部, 地势偏高, 牧草稀疏, 降水少、气温低, 气候条件恶劣, 碳汇能力有限, 属于弱碳汇区。年植被NEP平均值为25.93 gC m-2a-1, 年平均固碳量为1.07 TgC, 16年累积植被NEP平均值为414.71 gCm2, 累积固碳总量约为17.117 TgC。
Ⅴ江河源生态区
该生态区年植被NEP平均值在32.21—167.61 gC m-2a-1, 多年累积固碳总量为0.669 PgC, 是青海高原主要的碳汇区。区内不同生态子区之间固碳量差异与区域植被分布差异及水热耦合作用相异有关。
Ⅴ—1长江源高寒草甸草原生态区, 地处三江源西部与南部, 该区降水量偏少, 气温偏低, 植被NEP由西北向东南渐增, 碳汇作用逐步增强。植被年NEP平均值为99.72 gC m-2a-1, 年平均固碳量约为16.26 TgC, 16年累积植被NEP平均值为1593.79 gCm2, 累积固碳总量约为260.16 TgC。
Ⅴ—2澜沧江源高寒草甸草原生态区, 植被以高寒草甸草原为主, 区内水热组合条件良好, 草地资源丰富, 植被NEP较高, 植被年NEP平均值为159.34 gC m-2a-1, 年总固碳量为6.07 TgC;16年累积植被NEP平均值为2544.34 gCm2, 累积固碳总量约为97.12 TgC。
Ⅴ—3黄河源高寒草甸草原生态区, 位于三江源东部, 该区植被覆盖类型以草地为主[42-43], 低、中等覆盖度草地占绝对优势, 降水充足、气温适宜, 水热耦合作用良好, 植被年NEP平均值为167.61 gC m-2a-1, 年平均固碳量为16.39 TgC, 16年累积植被NEP平均值为2679.78 gCm2, 累积固碳总量约为262.24 TgC, 该区NEP较高, 植被固碳能力强。
Ⅴ—4共和盆地高寒草原生态区, 植被NEP自北向南逐步增加, 年NEP平均值为32.21 gC m-2a-1, 年平均固碳量为0.54 TgC, 16年累积植被NEP平均值为515.16 gC/m2, 累积固碳总量约为8.64 TgC。
Ⅴ—5海东—甘南高寒草甸草原生态区, 降水量充沛, 气温适宜, 植被年NEP平均值为158.71 gC m-2a-1, 年平均固碳量为2.54 TgC, 16年累积植被NEP平均值为2537.51 gC/m2, 累积固碳总量约为40.64 TgC。
3.3 青海高原植被碳源/汇时空变化特点 3.3.1 植被碳源/汇年际变化特点2000—2015年, 青海高原年植被NEP分布在-3.2—64.42 TgC, 植被碳循环总体以碳汇为主。2000—2015年植被NEP总值年际变化率为1.06 TgC/a, 植被固碳总值逐年增加, 呈平稳上升趋势(图 5)。

|
| 图 5 2000—2015年青海高原年NEP总值年际变化 Fig. 5 Inter-annual variation of the annual NEP value of Qinghai Plateau from 2000 to 2015 |
依据植被年NEP总值变化趋势将2000—2015年划分为以下几个阶段:2000年, 研究区植被覆盖度低, 气候干燥少雨, 植被总体固碳量较少, 土壤呼吸排放碳量大于植被固定碳量, 植被起碳源作用, 总固碳量为-3.2 TgC;2001年, 青海高原降水量增加、气温升高, 气候条件的改善使植被固碳量增加, 去除土壤呼吸消耗碳, 植被发挥碳汇作用;2002—2015年, 植被固碳出现四个波峰, 固碳总值在波动中较前期明显增加, 土壤呼吸排放碳量小于植被固定碳量, 植被固碳表现为碳汇。其中, 2002—2008年, 年NEP总值基本在多年平均值附近波动, 呈平稳增加态势, 并在2002、2006年达到阶段性最大值;2007、2008年NEP总值较2006年稍有下降, 但总体固碳水平仍体现为碳汇;2009—2012年, 年NEP总值较前期明显提升, 随着气候的逐步暖湿化, 植被的生长条件改善、生长期延长, 植被光合作用总体固碳量增加, 植被总碳汇量增加;2013年, 由于研究区降水量偏少, 气温偏高, 气候干燥少雨, 植被NPP总体偏少, 去除土壤呼吸消耗之后, NEP总值较2012年明显减少;2014年, 虽植被NPP总值较2013年有所增加, 但土壤呼吸消耗也相继增加, 植被NEP总值较2013年略有下降;2015年的气象条件与2013年较为相似, 表现为干燥少雨, 虽降水量较往年偏少但气温适宜植被生长, 且受2014年气候条件对植被的滞后影响, 其植被NPP总值较往年显著增加, 扣除土壤呼吸消耗, 植被NEP总值较前期上升, 植被固碳体现为碳汇。
总体上, 2000—2015年青海高原碳汇功能逐步增强, 年净固碳量在42.86 TgC与64.42 TgC之间波动。
3.3.2 植被碳汇空间变化特点2000—2015年, 青海高原年NEP呈现逐步增加的趋势(图 6), 平均趋势系数为1.52。年NEP增加的区域占植被总面积的25.72%, 其中, 年NEP明显增加的区域占植被总面积的8.22%, 集中分布于Ⅲ—4湟水谷地农业生态区的中部、南部, Ⅴ—3黄河源高寒草甸草原生态区的东北部, Ⅴ—4共和盆地高寒草原生态区南部, 以及Ⅴ—5海东—甘南高寒草甸草原生态区的中部, 这些区域处于青海高原河湟谷地及周围地区, 温度适宜、水源丰富, 植被NEP明显增加, 且降水量、气温年际变化幅度较小, 土壤呼吸消耗碳量偏少, 植被碳汇能力逐年增加。同时, 自2000年以来, 一系列的生态保护工程实施, 包括退耕还林还草、已垦草原还草、草地病虫害、鼠害治理、水土保持与治理等[43], 改善了这些区域植被生态环境的状况, 有利于植被固碳能力, 增强了植被碳汇功能;年NEP轻微增加区域占植被总面积的17.5%, 主要分布于Ⅰ—2昆仑山东段高寒荒漠草原生态区的西北部与东南部, 该区植被覆盖度增加, 植被固碳能力增强, 总固碳量大于土壤呼吸消耗, 植被碳汇功能增强。

|
| 图 6 青海高原植被NEP空间变化趋势 Fig. 6 Spatial variation trend of vegetation NEP in Qinghai Plateau |
2000—2015年, 青海高原年NEP减少的区域较少, 仅占植被总面积的1.06%, 其中, 明显减少的区域为0.29%, 轻微减少的区域为0.77%, 零星分布于Ⅲ—2东祁连山生态区东南部、Ⅲ—3青海湖湿地生态区东北部、Ⅲ—4湟水谷地生态区中部, 以及Ⅴ—1、Ⅴ—2、Ⅴ—3三江源流域生态区的东南部, 这些区域年降水量、年均温逐步增加, 气候条件的逐步暖湿化, 使土壤呼吸消耗碳增加, 多年植被NEP有减小趋势, 导致区域碳汇功能下降。
2000—2015年, 青海高原其余植被分布区植被年NEP基本稳定, 约占青海高原植被总面积的39%。总体上, 研究区年NEP空间变化中, 以基本稳定、轻微好转、明显好转为主, 所占植被总面积百分比分别为39%、17.5%、8.22%。
3.3.3 青海高原植被碳汇变化趋势分析2000—2015年, 青海高原植被NEP变异系数空间分布呈中部高边缘低趋势, 青海高原不同生态区植被NEP变异系数的空间分布存在明显的差异, 变异系数以中等、较高、高波动居多(图 7)。

|
| 图 7 青海高原植被NEP变异系数 Fig. 7 Coefficient of variation of vegetation NEP in Qinghai Plateau |
高波动地区主要分布在柴达木盆地东部、祁连山地北部、环青海湖湖区周边、河湟谷地流域、长江源区的中部与东南部、黄河源区西北部, 说明这些区域植被NEP易受到自然或人为因素的干扰, 稳定性较低。结合高波动地区植被NEP空间变化趋势(图 6), 这些区域植被NEP呈增加态势, 说明这部分地区在生态保护工程的实施、气候暖湿化的条件下, 植被碳汇功能得到增强;
中等波动区集中分布于青海高原东南部, 即祁连山地东部、环青海湖流域、湟水谷地北部、海东—甘南生态区南部、三江源地区东南部;较低波动区分布于柴达木盆地中部、青海湖南部、东部地区、长江源西北部、黄河源东南部亦有零星分布, 说明这部分区域植被NEP较为稳定, 受外界干扰较少。
植被NEP低、较低、中等、较高、高波动区占青海高原植被总面积的比例分别为1.42%、6.26%、20.25%、5.06%和9.11%, 说明研究区大部分植被NEP动态变化以较低、中等波动为主, 稳定性较高。
3.4 青海高原NEP与气候因素的相关性青海高原年气候因素空间分布差异明显, 降水量从西北向东南递增, 气温的垂直变化明显。2000—2015年, 青海高原年平均降水量为1—774 mm, 年均温-14—10 ℃。
植被NEP与降水量的平均相关系数为0.075, 总体上植被NEP与降水量呈正相关(图 8)。研究区植被NEP与降水量呈显著正相关的区域占总植被面积的13.26%。主要分布于祁连山地东南部(Ⅲ—1、Ⅲ—2生态区)、青海湖周边湿地区域(Ⅲ—3区)、湟水谷地农业区(Ⅲ—4区), 以及长江源生态区的西北部(Ⅴ—1区), 这些区域植被NEP随降水量的增加而升高;植被NEP与降水量显著负相关的区域占总植被面积比为3.75%, 分布于柴达木盆地中部及三江源地区东南部, 该地区随降水量增加, 土壤呼吸消耗碳增加, 植被NEP减少。

|
| 图 8 NEP与降水量、气温相关性 Fig. 8 Correlation between NEP and precipitation, temperature |
在气温不变的情况下, NEP与降水量的偏相关分析表明(图 8), 显著正相关区域占总植被面积为7.28%。主要分布于祁连山地东南部(Ⅲ—2区)、澜沧江生态区的西北部(Ⅴ—2区)及黄河源生态区(Ⅴ—3区), 说明这些区域降水量、气温对植被生长的影响以耦合作用为主, 两者共同作用于植被生长。
植被NEP与气温的平均相关系数为-0.20(图 8), 以显著负相关为主, 负相关区域占总植被面积的27.90%, 占据着青海高原植被分布区的大部分面积, 说明气温的升高阻碍NEP积累, 这可能是由于气温的升高使得植被蒸腾作用加强, 导致植被生长所需水热不同步, 使植被积累的NPP总量较少, 而气温的升高又致使土壤呼吸作用增加, 消耗大量有机碳, 故植被NEP减少;植被NEP与气温显著正相关区域占总植被面积的0.16%, 零星的分布在青海高原的东南部, 这进一步充分说明气温对研究区植被NEP以阻碍作用为主。
在控制降水量对NEP的影响的条件下, 气温与NEP显著负相关的区域占总植被面积比例下降为16.37%, 显著正相关区域的面积比稍有增加(图 8)。空间分布上, 主要分布于三江源生态区的东南部(Ⅴ—1、Ⅴ—3区), 表明该地区气候因素对植被生长的作用以耦合作用为主, 降水量、气温共同调节植被NEP的变化与积累。
植被NEP与气候因素的相关、偏相关分析表明, 降水量对NEP积累以促进作用为主, 气温对其以抑制作用为主, 气温对研究区植被NEP的影响范围明显高于降水量, 且降水、气温对NEP的耦合作用较为显著。
3.5 讨论(1) 青海高原植被NEP与降水量、气温与之间的相关分析表明, 降水量对植被NEP起促进作用的区域占植被分布区的13.26%, 对植被生长起到良好促进作用的区域仅占总植被分布区的4.98%, 其中, 0.04%的分布区, 气温对植被NEP亦表现为促进作用;其余4.94%的分布区仅降水量对植被NEP表现为促进作用, 气温对植被NEP的影响不明显。另外, 占植被分布区8.28%的区域, 由于受气温对NEP抑制作用的影响, 使降水量对NEP的积极作用削弱;
(2) 气温对NEP起阻碍作用的区域占植被分布区面积的27.90%, 明显高于降水量对NEP影响的区域, 其中, 8.28%的分布区由于降水量对NEP的积极作用, 使气温对植被固碳的阻碍作用有所缓解, 促进了植被固碳;气温对植被NEP起较强抑制作用的区域占植被分布区的19.62%, 其中3.01%的区域, 降水量对植被NEP也表现为抑制作用, 气象因素阻碍植被NEP积累;剩余16.61%的区域中, 仅气温对植被NEP表现为抑制作用, 降水量对植被NEP的影响不显著。这说明气温是控制青海高原植被NEP积累的主要气候因素, 部分区域降水量、气温对植被NEP的耦合作用较为明显。
(3) 本文通过估算植被NEP在一定程度上揭示了青海高原植被碳源/汇空间分布的特征与规律。因植被碳循环、土壤呼吸均是较为复杂的生态学过程, 通过模型估算均存在一定的不确定性。受NPP估算和RH估算不确定性的影响, NEP估算结果仍存在一定程度的不确定性。
(4) 本文仅考虑青海高原不同生态区的植被NEP、碳汇时空变化, 定量分析了气象因素对NEP的影响, 但植被NEP是自然和人类活动共同作用的结果, 除气温和降水量外, 大气CO2浓度、地表蒸散、地形因子、土壤、放牧、人工围栏等对植被NEP空间分布和植被碳收支均具有重要影响。因此, 进一步研究中将定量的分析人类活动、气象因素、地形因素对植被NEP动态波动的影响, 以及不同植被类型的固碳能力与年际变化, 以便更为深刻地认识青海高原地区植被生态系统碳循环的增汇减源贡献机制。
4 结论本研究在土壤呼吸模型和改进的CASA模型基础上, 估算青海高原植被NEP, 分析了2000—2015年青海高原植被NEP、区域碳源/汇的时空格局、年际动态变化与稳定性, 并就气候因素变化对植被NEP的变化进行了多角度定量分析, 主要结论如下:
(1) 2000—2015年, 青海高原植被年NEP空间分布呈东高西低、南高北低, 由西北向东南逐步增加趋势, 不同生态区之间差异显著, 碳汇区集中于祁连山生态区、三江源生态区。
(2) 2000—2015年, 青海高原植被年碳汇功能逐步增强, 植被NEP变异系数的空间分布以较低、中等波动为主, 稳定性较高。
(3) 2000—2015年, 青海高原年NEP空间变化中, 以基本稳定、轻微好转、明显好转为主。年际波动以中等波动为主, 稳定性较高, 自然与人为因素对植被NEP的改善作用明显。
(4) 青海高原降水量对植被NEP以促进作用为主, 气温以抑制作用为主, 气温对NEP的影响范围强于降水。去除两者之间的相互影响, 降水量对NEP起促进作用的面积比为4.94%, 气温对NEP起抑制作用的面积比为16.61%。
| [1] |
於琍, 朴世龙. IPCC第五次评估报告对碳循环及其他生物地球化学循环的最新认识. 气候变化研究进展, 2014, 10(1): 33-36. DOI:10.3969/j.issn.1673-1719.2014.01.007 |
| [2] |
Wang Q F, Zheng H, Zhu X J, Yu G R. Primary estimation of Chinese terrestrial carbon sequestration during 2001-2010. Science Bulletin, 2015, 60(6): 577-590. DOI:10.1007/s11434-015-0736-9 |
| [3] |
杨绪红, 金晓斌, 项晓敏, 范业婷, 刘晶, 单薇, 周寅康. 近300年中国耕地开垦导致的碳排放估算. 中国科学: 地球科学, 2019, 49(3): 554-568. |
| [4] |
Mekonnen Z A, Grant R F, Schwalm C. Sensitivity of modeled NEP to climate forcing and soil at site and regional scales: implications for upscaling ecosystem models. Ecological Modelling, 2016, 320: 241-257. DOI:10.1016/j.ecolmodel.2015.10.004 |
| [5] |
宋金明, 曲宝晓, 李学刚, 袁华茂, 李宁, 段丽琴. 黄东海的碳源汇: 大气交换、水体溶存与沉积物埋藏. 中国科学: 地球科学, 2018, 48(11): 1444-1455. |
| [6] |
朴世龙, 张新平, 陈安平, 刘强, 连旭, 王旭辉, 彭书时, 吴秀臣. 极端气候事件对陆地生态系统碳循环的影响. 中国科学: 地球科学, 2019, 49(9): 1321-1334. |
| [7] |
吴艳艳, 吴志峰, 余世孝. 定量评价人类活动对净初级生产力的影响. 应用生态学报, 2017, 28(8): 2535-2544. |
| [8] |
王伯炜, 牟长城, 王彪. 长白山原始针叶林沼泽湿地生态系统碳储量. 生态学报, 2019, 39(9): 3344-3354. |
| [9] |
徐丽, 于贵瑞, 何念鹏. 1980s-2010s中国陆地生态系统土壤碳储量的变化. 地理学报, 2018, 73(11): 2150-2167. DOI:10.11821/dlxb201811008 |
| [10] |
戴尔阜, 黄宇, 吴卓, 赵东升. 内蒙古草地生态系统碳源/汇时空格局及其与气候因子的关系. 地理学报, 2016, 71(1): 21-34. |
| [11] |
Zhou W, Huang L, Yang H, Ju W M, Yue T X. Interannual variation in grassland net ecosystem productivity and its coupling relation to climatic factors in China. Environmental Geochemistry and Health, 2019, 41(3): 1583-1597. DOI:10.1007/s10653-018-0236-3 |
| [12] |
牛犇, 张宪洲, 何永涛, 石培礼, 付刚, 杜明远, 张扬建, 宗宁, 张晶, 武建双. 青藏高原高寒沼泽化草甸总初级生产力的遥感估算: 多模型比较(英文). 资源与生态学报, 2017, 8(1): 57-66. |
| [13] |
张镱锂, 祁威, 周才平, 丁明军, 刘林山, 高俊刚, 摆万奇, 王兆锋, 郑度. 青藏高原高寒草地净初级生产力(NPP)时空分异(英文). 地理学报, 2014, 24(2): 269-287. |
| [14] |
孙庆龄, 李宝林, 李飞, 张志军, 丁玲玲, 张涛, 许丽丽. 三江源植被净初级生产力估算研究进展. 地理学报, 2016, 71(9): 1596-1612. |
| [15] |
邵全琴, 曹巍, 樊江文, 黄麟, 徐新良. 三江源生态保护和建设一期工程生态成效评估(英文). 地理学报, 2017, 27(2): 183-204. |
| [16] |
Deng X Z, Gibson J, Wang P. Quantitative measurements of the interaction between net primary productivity and livestock production in Qinghai Province based on data fusion technique. Journal of Cleaner Production, 2017, 142: 758-766. DOI:10.1016/j.jclepro.2016.05.057 |
| [17] |
方精云, 郭兆迪, 朴世龙, 陈安平. 1981-2000年中国陆地植被碳汇的估算. 中国科学D辑: 地球科学, 2007, 37(6): 804-812. |
| [18] |
Chen H, Zhu Q, Peng C H, Wu N, Wang Y F, Fang X Q, Gao Y H, Zhu D, Yang G, Tian J Q, Kang X M, Piao S L, Ouyang H, Xiang W H, Luo Z B, Jiang H, Song X Z, Zhang Y, Yu G R, Zhao X Q, Gong P, Yao T D, Wu J H. The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau. Global Change Biology, 2013, 19(10): 2940-2955. DOI:10.1111/gcb.12277 |
| [19] |
张继平, 刘春兰, 郝海广, 孙莉, 乔青, 王辉, 宁杨翠. 基于MODIS GPP/NPP数据的三江源地区草地生态系统碳储量及碳汇量时空变化研究. 生态环境学报, 2015, 24(1): 8-13. |
| [20] |
王斌, 李洁, 姜微微, 赵亮, 古松. 草地退化对三江源区高寒草甸生态系统CO2通量的影响及其原因. 中国环境科学, 2012, 32(10): 1764-1771. DOI:10.3969/j.issn.1000-6923.2012.10.006 |
| [21] |
任小丽, 何洪林, 张黎, 葛蓉, 冯艾琳, 于贵瑞, 张林波. 2001-2010年三江源区草地净生态系统生产力估算. 环境科学研究, 2017, 30(1): 51-58. |
| [22] |
周夏飞, 於方, 曹国志, 杨威杉, 周颖. 2001-2015年青藏高原草地碳源/汇时空变化及其与气候因子的关系. 水土保持研究, 2019, 26(1): 76-81. |
| [23] |
赵健赟, 彭军还. 基于MODIS NDVI的青海高原植被覆盖时空变化特征分析. 干旱区资源与环境, 2016, 30(4): 67-73. |
| [24] |
国家环境保护总局, 国家测绘局. 中国西部地区生态环境现状遥感调查图集. 北京: 科学出版社, 2002.
|
| [25] |
陈晓玲, 曾永年. 亚热带山地丘陵区植被NPP时空变化及其与气候因子的关系——以湖南省为例. 地理学报, 2016, 71(1): 35-48. |
| [26] |
刘春雨, 董晓峰, 刘英英, 潘竟虎, 车彦军. 甘肃省净初级生产力时空变化特征. 中国人口·资源与环境, 2014, 24(1): 163-170. DOI:10.3969/j.issn.1002-2104.2014.01.023 |
| [27] |
巩杰, 张影, 钱彩云. 甘肃白龙江流域净生态系统生产力时空变化. 生态学报, 2017, 37(15): 5121-5128. |
| [28] |
朱文泉. 中国陆地生态系统植被净初级生产力遥感估算及其与气候变化关系的研究[D]. 北京: 北京师范大学, 2005.
|
| [29] |
朱文泉, 陈云浩, 潘耀忠, 李京. 基于GIS和RS的中国植被光利用率估算. 武汉大学学报: 信息科学版, 2004, 29(8): 694-698, 714-714. |
| [30] |
朱文泉, 潘耀忠, 何浩, 于德永, 扈海波. 中国典型植被最大光利用率模拟. 科学通报, 2006, 51(6): 700-706. DOI:10.3321/j.issn:0023-074X.2006.06.014 |
| [31] |
周广胜, 张新时. 自然植被净第一性生产力模型初探. 植物生态学报, 1995, 19(3): 193-200. DOI:10.3321/j.issn:1005-264X.1995.03.005 |
| [32] |
裴志永, 周才平, 欧阳华, 杨文斌. 青藏高原高寒草原区域碳估测. 地理研究, 2010, 29(1): 102-110. DOI:10.3969/j.issn.1004-9479.2010.01.015 |
| [33] |
王强, 张廷斌, 易桂花, 陈田田, 别小娟, 何奕萱. 横断山区2004-2014年植被NPP时空变化及其驱动因子. 生态学报, 2017, 37(9): 3084-3095. |
| [34] |
潘竟虎, 黄克军, 李真. 2001-2010年疏勒河流域植被净初级生产力时空变化及其与气候因子的关系. 生态学报, 2017, 37(6): 1888-1899. |
| [35] |
吴珊珊, 姚治君, 姜丽光, 王蕊, 刘兆飞. 基于MODIS的长江源植被NPP时空变化特征及其水文效应. 自然资源学报, 2016, 31(1): 39-51. |
| [36] |
范月君. 围栏与放牧对三江源区高山嵩草草甸植物形态、群落特征及碳平衡的影响[D]. 兰州: 甘肃农业大学, 2013.
|
| [37] |
牛志春, 倪绍祥. 青海湖环湖地区草地植被生物量遥感监测模型. 地理学报, 2003, 58(5): 695-702. DOI:10.3321/j.issn:0375-5444.2003.05.007 |
| [38] |
Luo T X, Pan Y D, Ouyang H, Shi P L, Luo J, Yu Z L, Lu Q. Leaf area index and net primary productivity along subtropical to alpine gradients in the Tibetan Plateau. Global Ecology and Biogeography, 2004, 13(4): 345-358. DOI:10.1111/j.1466-822X.2004.00094.x |
| [39] |
王莉雯, 卫亚星. 青海省植被净初级生产力的模拟研究. 第四纪研究, 2011, 31(1): 180-188. DOI:10.3969/j.issn.1001-7410.2011.01.22 |
| [40] |
孙庆龄, 李宝林, 周成虎, 李飞, 张志军, 丁玲玲, 张涛, 许丽丽. 三江源植被净初级生产力估算研究进展(英文). 地理学报, 2017, 27(2): 161-182. |
| [41] |
张颖, 章超斌, 王钊齐, 杨锐, 张艳珍, 李建龙, 安如. 气候变化与人为活动对三江源草地生产力影响的定量研究. 草业学报, 2017, 26(5): 1-14. |
| [42] |
刘凤, 曾永年. 近16年青海高原植被NPP时空格局变化及气候与人为因素的影响. 生态学报, 2019, 39(5): 1528-1540. |
| [43] |
邵全琴, 樊江文, 刘纪远, 黄麟, 曹巍, 刘璐璐. 基于目标的三江源生态保护和建设一期工程生态成效评估及政策建议. 中国科学院院刊, 2017, 32(1): 35-44. |