 2021, Vol. 41
2021, Vol. 41文章信息
- 刘旻霞, 南笑宁, 张国娟, 李博文, 徐璐, 穆若兰, 李亮, 于瑞新
- LIU Minxia, NAN Xiaoning, ZHANG Guojuan, LI Bowen, XU Lu, MU Ruolan, LI Liang, YU Ruixin
- 高寒草甸不同坡向植物群落物种多样性与功能多样性的关系
- Relationship between species diversity and functional diversity of plant communities on different slopes in alpine meadow
- 生态学报. 2021, 41(13): 5398-5407
- Acta Ecologica Sinica. 2021, 41(13): 5398-5407
- http://dx.doi.org/10.5846/stxb202003250690
-
文章历史
- 收稿日期: 2020-03-25
- 网络出版日期: 2021-04-28





物种多样性与功能多样性是生态系统功能研究的重要内容[1]。物种多样性作为生物多样性的重要组成部分, 主要反映了生物资源的丰富程度[2], 物种多样性的增加会提高群落或生态系统的稳定性与生产力[3]。功能多样性是通过植物功能性状来反映植物在生态系统中的功能差异及其分布范围, 其不仅具有预测生态系统功能的变化能力, 也可以提供更多群落结构、功能及对资源利用状况的信息[4]。功能多样性是物种与其所在的环境共同作用的结果, 且功能多样性和物种多样性也是生态系统功能必不可少的预测因子[5], 因此, 在了解生物多样性(功能多样性和物种多样性)是如何影响生态系统功能, 应阐明其两者之间的关系[6]。由于环境条件的不同[7]、物种组成的差异[8]等, 目前对于物种多样性与功能多样性的关系(正相关、负相关、S形曲线、不相关)未达成一致。所以, 研究异质环境条件下生态系统物种多样性与功能多样性的关系, 既可以增加对物种多样性与功能多样性的了解, 也可以促进生物多样性保护理论的发展。
作为亚欧板块最大的草地区域, 青藏高原的天然草地面积约占全国草地面积的1/3, 其因寒冷、湿润的气候, 地理环境和生态构造较为独特。高寒草地是青藏高原生态系统的主体, 是高寒生态系统物种及遗传基因最丰富和最集中的地区之一, 在全球高寒生物多样性保护中具有十分重要的地位[9]。近年来, 在气候变化、人为干扰(放牧业过度发展、旅游业过度开发)等因素的影响之下, 该地区的草场退化严重、生物多样性急剧减少[10], 亟需加强对该生态系统生物多样性保护的研究。目前, 关于高寒草甸植物群落多样性大多只对单一的物种多样性或功能多样性进行研究[11-12], 而未将两者结合起来更加全面的考虑, 导致高寒草甸植物群落生态功能优化、多样性保护仅单纯立足于物种的数量状况或功能特征。而对于物种多样性与功能多样性多维的研究涉及较少。目前相关的研究主要是针对不同放牧方式下物种与功能多样性的关系[13]、高寒草甸植物群落多样性与初级生产力的关系[14]、植物多样性对不同干扰方式的响应[15]等的研究, 而对于坡向梯度上的植物群落物种多样性与功能多样性关系的研究较少。
基于这一研究背景, 本文选取青藏高原高寒草甸为研究对象, 开展不同坡向植物群落物种多样性与功能多样性的研究。我们进行如下假设:不同坡向梯度上, 植物群落物种多样性(Margalef、Simpson、Shannon及Pielou指数)与功能多样性(功能丰富度、功能均匀度及功能离散度)之间存在正线性相关关系。对此问题的科学解答, 有助于更好的理解植物群落的构建机制, 生物多样性保护与生态系统功能和过程间的关系等有重要意义, 同时为黄河上游生物多样性保护与生态功能恢复及优化提供理论依据。
1 材料与方法 1.1 自然概况本试验区位于甘肃南部玛曲县境内的“兰州大学高寒草甸与湿地生态系统定位研究站”-阿孜站(33°58′N, 101°53′E), 平均海拔3500 m, 属于高寒湿润型气候;年平均气温1.2 ℃, 1月平均气温-10 ℃, 7月平均气温11.7 ℃, 年无霜期大约90 d左右;6—9月份降水较为充沛, 年平均降水量620 mm。土壤属于高寒草甸土, 该地区植被类型丰富, 优势种为:长毛风毛菊(Saussurea hieracioides)、线叶龙胆(Gentiana farreri)、黄芪(Astragalus membranaceus)、圆叶堇菜(Viola striatella)、金露梅(Potentilla fruticosa)、鹅绒委陵菜(Potentilla anserina)、秦艽(Gentiana macrophylla)、黄帚橐吾(Ligularia virgaurea)等多年生草本植物。
1.2 取样方法于2018年7月中旬到8月上旬(植物生长旺盛期) 选择一座山体分异明显的山坡进行野外调查, 用GPS进行坡向定位, 沿顺时针方向依次选取5个不同坡向(即北、西北、西、西南和南坡)。在每个坡向的同一高度设置8个50 cm×50 cm小样方, 样方间距为1 m, 共40个样方。对样方内物种进行生物学调查, 同时测量各项环境指标(坡位、坡向、坡度、海拔)。选择前3天内未降雨的天气进行土壤采集, 选用直径为50 mm的土钻采用对角线法采集土壤, 土层深度为0—20 cm, 去除杂质后, 将其装入自封袋带回实验室, 尽快完成土壤鲜重的称量, 再将其放置在105℃的烘箱烘至恒重, 测其干重。剩余土壤样品自然风干后用于后续实验。采集每个样方内每种无病虫害的物种叶片, 标记后用于后期试验分析。
1.3 环境因子的计测土壤酸碱度采用电位法, 重复3次;土壤含水量采用烘干法测定, 重复3次;使用DT-131土温计测定土壤温度;使用HT8318照度仪测定不同坡向的光照强度;土壤有机碳采用重铬酸钾外加热法测定;土壤全氮采用微量凯氏法测定;全磷采用钼锑抗比色法测定[16]。
1.4 叶片功能性状的计测本研究选取了8个植物叶功能性状, 分别为:植物高度(Plant height, PH)、叶含水量(Leaf water content, LWC)、比叶面积(Specific leaf area, SLA)、叶干物质量(Leaf dry matter content, LDMC)、叶有机碳含量(Leaf carbon content, LCC)、叶全氮含量(Leaf nitrogen content, LNC)、叶全磷含量(Leaf phosphorus content, LPC)及叶全钾含量(Leaf kalium content, LKC) 等。每个样方内每种物种随机选择8—10株测量其株高(PH), 取其平均值。用CanoScan LiDE 110扫描仪扫描叶面积, 用烘箱(75℃)烘干后称叶重量。叶含水量=(鲜重-干重)/鲜重, 比叶面积=叶面积/叶干重, 叶干物质含量=叶干重/叶饱和鲜重。将采集剩余的植物叶片样本放入烘箱烘干至恒重, 用研钵磨成细粉, 进行养分含量的测定, 植物养分含量的测定方法同土壤, 植物叶片全钾的测定采用火焰光度计法[17]。
1.5 多样性指数的计算 1.5.1 物种多样性指数(1) Margalef指数
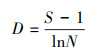
|
(1) |
(2) Simpson指数

|
(2) |
(3) Shannon-Wiener指数

|
(3) |
(4) Pielou指数

|
(4) |
式中, N为总个体数量, S为总物种数量, Ni为第i个种的个体数量, Pi为第i个物种的相对重要值。
1.5.2 功能多样性指数(1) 一元性状功能多样性
采用R语言“FD”包中的Rao′s二次方程指数(Rao's quadratic entropy, FDrao)进行计算[18]。

|
(5) |
式中, dij为物种i与物种j功能特征距离, pi为物种i的个体数占总个体数的比例。
(2) 多元性状功能多样性
多元性状的功能多样性用3个主要指数进行计算:
1) 功能丰富度
2005年Mason等提出用functional range指数来计算多元性状功能丰富度(functional richness, FRic) [19]。其计算公式为:

|
(6) |
式中, 物种在群落中所占的生态位用SFic表示;性状c在群落中的绝对特征值大小用Rc表示。
2) 功能均匀度
计算多元性状的功能均匀度(functional evenness, FEve) 指数用Villeger等提出的公式计算[20]:

|
(7) |

|
(8) |

|
(9) |
式中, EW表示均匀度, dist(i, j)表示物种i和j 的欧式距离, wi表示i物种的数目。
3) 功能离散度
利用Laliberte和Legendre提出的公式[18]来计算功能离散度指数(functional dispersion, FDis):

|
(10) |

|
(11) |
式中, zj表示群落中物种j到重心c的加权欧几里得距离。
1.6 数据分析与处理采用Excel 2010对所有数据进行计算, 用R 3.5.2软件进行功能多样性的拟合运算, 并用SPSS 18.0进行如下统计分析:1) 不同坡向的环境因子、物种多样性以及功能多样性进行单因素方差分析(One-Way ANOVA)并用LSD后置检验法检验;2) 功能多样性与环境因子的逐步回归分析与通径分析;3) 物种多样性与功能多样性之间的Pearson相关性分析。运用Origin 2018作图。
2 结果与分析 2.1 南北坡梯度上土壤理化因子变化不同坡向间的光照度(LI)与土温(ST)存在显著差异(P < 0.05), 总体趋势均为南坡大于北坡。南坡的土壤含水量(19.69%)明显低于其他坡向;土壤pH南坡明显高于北坡, 南坡偏碱性(7.85), 北坡偏酸性(6.45), 坡向间差异显著(P < 0.05);坡度(SG)由南坡向北坡逐渐递减, 坡向间差异显著(P < 0.05);北坡与西北坡的SOC、STN与STP高于其他坡向, 北坡分别为:SOC(48.42)、STN(3.67)、STP(0.48), 不同坡向具有显著性差异(P < 0.05), 且西坡均为最低值(表 1)。
| 坡向 Slope aspect |
光照度LI/(103lx) | 土温 ST/(℃) |
土壤含水量 SWC/% |
土壤pH | 坡度SG/° | 土壤有机碳 SOC/(g/kg) |
土壤全氮 STN/(g/kg) |
土壤全磷 STP/(g/kg) |
| 北坡(N) | 76.98±2.14d | 19.87±0.64c | 31.06±0.75a | 6.45±0.26c | 22.04±1.19d | 56.85±1.51a | 5.37±0.32a | 0.69±0.02a |
| 西北坡(NW) | 77.58±1.98c | 20.65±0.28c | 30.44±1.34a | 6.68±0.24c | 23.44±1.70d | 52.86±1.98b | 4.83±0.27b | 0.70±0.03a |
| 西坡(W) | 77.82±2.01c | 21.46±0.81b | 25.28±0.81b | 6.94±0.26b | 26.17±1.23c | 48.42±2.23d | 3.67±0.47c | 0.48±0.08c |
| 西南坡(SW) | 79.00±3.05b | 22.22±0.94ab | 21.26±1.86c | 7.25±0.27a | 28.02±1.07b | 50.76±1.26c | 4.73±0.24b | 0.56±0.04b |
| 南坡(S) | 81.22±2.19a | 23.54±1.21a | 19.69±1.57c | 7.85±0.19a | 30.41±1.06a | 50.89±1.44c | 4.47±0.18b | 0.52±0.02b |
| 不同的字母表示差异显著(a=0.05); LI: 光照度Illumination; ST: 土温Soil temperature; SWC: 土壤含水量Soil water content; SOC: 土壤有机碳Soil organic carbon; STP: 土壤有机碳Soil total phosphorus; SG: 坡度Slope gradient; pH: 土壤酸碱度Soil pH; STN: 土壤全氮Soil total nitrogen; S: 南坡South slope; N: 北坡North slope; SW: 西南坡Southwest slope; W: 西坡West slope; NW: 西北坡Northwest slope | ||||||||
从南坡到北坡的变化过程中, Margalef指数、Simpson指数、Shannon指数均呈逐渐增加的趋势。其中, Margalef指数、Simpson指数在北坡与西北坡、南坡与西南坡无显著差异, 但其他坡向间差异显著(P < 0.05);南坡与西南坡的Shannon指数无显著性差异(P>0.05), 但与西坡、西北坡及北坡之间存在显著性差异(P < 0.05);不同坡向梯度间的Pielou指数无显著性差异(P>0.05), 其在西北坡和西南坡出现最大值(0.83)和最小值(0.70)(图 1)。

|
| 图 1 南北坡梯度上物种多样性的变化 Fig. 1 Changes of species diversity on the south-north slope gradient S: 南坡South slope; N: 北坡North slope; SW: 西南坡Southwest slope; W: 西坡West slope; NW: 西北坡Northwest slope |
通过对不同坡向一元性状功能多样性(表 2)研究发现, 北坡的PHFDrao明显高于其他坡向(P < 0.05), 南坡与西南坡间差异不显著;LWCFDrao与LCCFDrao的变化一致, 从南坡向北坡逐渐递增, 南坡显著低于西南坡(P < 0.05);SLAFDrao的最大值(1.06)和最小值(0.53)分别出现在西北坡与西南坡, 且西坡与西南坡间有显著差异(P < 0.05);LDMCFDrao的变化趋势与其他指数相反, 由北坡向南坡逐渐增加, 且南坡与北坡差异性显著(P < 0.05);LNCFDrao北坡最高, 西坡最低, 西坡与西南坡、南坡与西北坡的差异不显著;北坡LPCFDrao最大, 南坡最小, 西坡与西南坡的差异较小;LKCFDrao由北坡向南坡递减(P < 0.05), 在西南坡最低。
| 坡向 Slope aspect |
株高功能多样性 PHFDrao |
叶片含水量功能多样性 LWCFDrao |
比叶面积功能多样性 SLAFDrao |
叶干物质含量功能多样性 LDMCFDrao |
叶片碳含量功能多样性 LCCFDrao |
叶片氮含量功能多样性 LNCFDrao |
叶片磷含量功能多样性 LPCFDrao |
叶片钾含量功能多样性 LKCFDrao |
| 北坡(N) | 0.87±0.03a | 1.07±0.07a | 1.03±0.11a | 0.47±0.13c | 0.83±0.07a | 0.99±0.07a | 0.69±0.03a | 0.74±0.03a |
| 西北坡(NW) | 0.72±0.05b | 1.09±0.04a | 1.06±0.12a | 0.64±0.15ab | 0.86±0.06a | 0.69±0.10b | 0.54±0.05b | 0.62±0.04b |
| 西坡(W) | 0.64±0.07c | 0.98±0.08b | 0.81±0.09b | 0.87±0.09b | 0.71±0.06b | 0.54±0.09c | 0.44±0.04c | 0.54±0.04c |
| 西南坡(SW) | 0.35±0.09d | 0.75±0.10c | 0.53±0.07c | 1.05±0.10a | 0.45±0.07c | 0.57±0.07c | 0.46±0.05c | 0.42±0.05d |
| 南坡(S) | 0.34±0.08d | 0.28±0.08d | 0.60±0.12c | 1.14±0.12a | 0.29±0.08d | 0.68±0.08b | 0.28±0.04d | 0.45±0.04d |
| 不同的字母表示差异显著(a=0.05); PHFDrao: 株高功能多样性Functional diversity of plant height; SLAFDrao: 比叶面积功能多样性Functional diversity of specific leaf area; LNCFDrao: 叶片氮含量功能多样性Functional diversity of leaf nitrogen content; LDMCFDrao: 叶干物质含量功能多样性Functional diversity of leaf dry matter content; LKCFDrao: 叶片钾含量功能多样性Functional diversity of leaf kalium content; LWCFDrao: 叶片含水量功能多样性Functional diversity of leaf water content; LCCFDrao: 叶片碳含量功能多样性; Functional diversity of leaf carbon content; LPCFDrao: 叶片磷含量功能多样性Functional diversity of leaf phosphorus content | ||||||||
图 2表明了多元性状功能多样性的变化特征。在从南坡向北坡的变化过程中, 功能丰富度(FRic)与功能离散度(FDis)呈递增趋势, 西北坡略高于北坡, 但差异性不显著(P>0.05), 功能均匀度变化趋势较为平缓, 坡向间差异性不显著(P>0.05)。

|
| 图 2 南北坡梯度上多元性状功能多样性 Fig. 2 Functional diversity of multi-dimensional traits on the south-north slope gradient |
逐步回归分析表明(表 3), 功能丰富度、功能均匀度、功能离散度均与环境因子都达到显著水平(P < 0.05)。由方程(a)可知, 随着土壤pH、含水量的升高, 功能丰富度呈降低趋势, 随着土温和土壤全磷含量的增加而升高;从方程(b)中可知, 功能均匀度随着坡度与土壤全磷的增加而增大, 随土壤pH增加而降低;从方程(c)中可知, 功能离散度随土温、土壤含水量、光照度的升高而增大, 随着土壤pH的升高而降低。
| 坡向 Slope aspect |
环境因子 Environmental factors |
功能多样性 Functional diversity |
||||||||||
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | Y1 | Y2 | Y3 | ||
| 北坡(N) | 76.981 | 19.871 | 31.059 | 6.451 | 22.041 | 56.849 | 5.376 | 0.694 | 2.528 | 0.608 | 0.755 | |
| 西北坡(NW) | 77.581 | 20.651 | 30.439 | 6.681 | 23.441 | 52.860 | 4.830 | 0.703 | 2.588 | 0.626 | 0.784 | |
| 西坡(W) | 77.821 | 21.461 | 25.279 | 6.941 | 26.171 | 48.420 | 3.668 | 0.483 | 2.172 | 0.661 | 0.673 | |
| 西南坡(SW) | 79.001 | 22.221 | 21.259 | 7.251 | 28.021 | 50.762 | 4.731 | 0.562 | 2.114 | 0.688 | 0.584 | |
| 南坡(S) | 81.221 | 23.541 | 19.689 | 7.851 | 28.411 | 50.889 | 4.469 | 0.519 | 1.430 | 0.648 | 0.418 | |
| Y1: 功能丰富度FRic; Y2: 功能均匀度FEve; Y3: 功能离散度FDis; X1: LI(103lx); X2: ST(℃); X3: SWC(%); X4: pH; X5: SG(°); X6: SOC(g/kg); X7: STN(g/kg); X8: STP (g/kg); Y1=2.335-3.213X4+0.997X2+1.929X8-0.008X3; Y2=0.515+0.025X5-0.079X4+0.065X8; Y3=-7.047-1.677X4+0.048X2+0.013X3+0.121X1 | ||||||||||||
由表 4可知, 环境因子对功能丰富度的直接通径系数大小为:pH < SWC < STP < ST, 对功能均匀度的直接通径系数大小为pH < STP < SG; 而对功能离散度的直接通径系数大小为pH < SWC < LI < ST。功能丰富度的环境因子决策系数均为负, 其大小为:pH < ST < STP < SWC; 功能均匀度的环境因子决策系数均为负, 其大小为:pH < SG < STP;功能离散度的环境因子决策系数大小为:ST < pH < LI < SWC, 其中, SWC的决策系数为正, 其余均为负。因此, 影响功能丰富度和功能均匀度的主要环境限定因子均为pH;而功能离散度的主要环境决定因子为SWC, 主要环境限定因子为ST。
| 功能多样性 Functional diversity |
影响因子 Impact factor |
相关系数 Correlation coefficient |
直接通径 Direct path coefficients |
间接通径 Indirect path coefficients |
决策系数 Decision coefficient |
|||||
| pH | ST | STP | SWC | 合计 | R2 | |||||
| 功能丰富度 | pH | -0.968 | -3.788 | 3.045 | -0.297 | 0.073 | 2.823 | -7.028 | ||
| Functional richness(FRic) | ST | -0.950 | 3.058 | -3.771 | -0.3141 | 0.076 | -4.009 | -3.550 | ||
| STP | 0.742 | 0.421 | 2.661 | -2.277 | -0.063 | 0.321 | -0.802 | |||
| SWC | 0.898 | -0.078 | 3.574 | -2.907 | 0.342 | 1.009 | -0.147 | |||
| SG | pH | STP | 合计 | R2 | ||||||
| 功能均匀度 | SG | 0.849 | 2.287 | -1.270 | -0.170 | -1.441 | -1.348 | |||
| Functional evenness (FEve) | pH | 0.572 | -1.386 | 2.098 | -0.143 | 1.957 | -3.501 | |||
| STP | -0.74 | 0.207 | -1.901 | 0.971 | -0.936 | -0.342 | ||||
| pH | ST | SWC | LI | 合计 | R2 | |||||
| 功能离散度 | pH | -0.975 | -6.17 | 4.291 | -0.436 | 1.346 | 5.204 | -26.128 | ||
| Functional dispersion(FDis) | ST | -0.95 | 4.311 | -6.148 | -0.443 | 1.319 | -5.272 | -26.871 | ||
| SWC | 0.937 | 0.460 | 5.829 | -4.136 | -1.211 | 0.481 | 0.651 | |||
| LI | -0.968 | 1.367 | -6.088 | 4.162 | -0.409 | -2.336 | -4.521 | |||
对南北坡梯度植物群落物种多样性与功能多样性的Pearson相关性检验(表 5)表明, 功能丰富度与Margalef指数、Simpson指数间存在显著正相关关系(P < 0.05), 相关性系数>0.8;Margalef指数、Simpson指数、Shannon指数与功能离散度呈显著负相关(P < 0.05), 相关性系数 < -0.9。
| 指数Index | Margalef指数 Margalef index |
Simpson指数 Simpson index |
Shannon指数 Shannon index |
Pielou指数 Pielou index |
| 功能丰富度(FRic) Functional richness | 0.894* | 0.895* | 0.868 | 0.766 |
| 功能均匀度(FEve) Functional evenness | 0.883 | 0.886 | -0.676 | -0.760 |
| 功能离散度(FDis) Functional dispersion | -0.933* | -0.935* | -0.936* | 0.862 |
| *.P < 0.05, **.P < 0.01 | ||||
坡向是重要地形因子之一, 其通过影响土壤养分、水分等生境条件可以改变群落物种的组成和分布[21]。本研究结果表明, 不同坡向间环境因子差异显著(表 1), 从南坡到北坡, 坡度、光照度、土温、pH均呈递减趋势, 而SWC、SOC、STN、STP逐渐增加。这主要是因为南坡坡度大, 受光照照射时间长, 导致植被稀少, 裸露土地面积大, 使土壤温度偏高, 蒸发量大, 土壤含水量降低, 土壤偏碱性, 这与前人的研究结果一致[22-23]。一般来讲, 坡度越大, 土壤养分含量越低, 这是由于受到地表径流的冲刷作用, 甘南地区雨季集中降水造成南坡土壤养分流失比较严重, 这也是导致其土壤养分含量比其他坡向低的原因之一[24]。此外, 北坡土壤环境较为湿润, 腐殖化程度较高, 具有较高的土壤养分与含水量。西坡的土壤养分也相对其他坡向较低, 这是由于西坡处于南北坡生境过渡的山脊通道地带, 受干扰较南坡与北坡大, 且土壤风化相对严重, 导致其土壤养分含量相对较低[25]。
3.2 不同坡向物种多样性的变化特征从南坡到北坡梯度上, 物种多样性(Margalef、Simpson和Shannon指数)明显增大(图 1), 这主要是因为土壤养分与含水量增加(表 1), 使得北坡能够容纳更多的物种生存, 导致该区域多样性增大。坡向在转变的过程中也会受降水、地表径流等的影响, 使得物种组成、原有的生态位发生变化, 继而出现新的物种来维持生态系统的平衡[26]。Pielou指数在不同坡向的变化不显著, 这与其他的研究结果一致, 即不同坡向存在不同的功能群, 且不同坡向的种间性状差异较小[26-28]。
3.3 一元性状功能多样性在南北坡梯度上的变化南北坡梯度上植物群落一元性状功能多样性的变化体现了植物对环境的适应性[29], 这也是环境筛选和物种间相互作用的结果[30]。本研究结果显示北坡的PHFDrao、LWCFDrao、SLAFDrao均高于其他坡向(表 2)。这是由于北坡(阴坡)具有相对较高的土壤营养资源, 植物能够采取快速获取外部资源和快回报的生存策略, 使得植物能够很好的生长[25]。而南坡的LDMCFDrao显著高于北坡, 相关研究表明[31], LDMCFDrao与SWC、土壤养分的竞争有关即南坡土壤资源匮乏, 植物对所获取的资源有较好的保留能力。北坡的一元性状功能多样性(LCCFDrao、LNCFDrao、LPCFDrao、LKCFDrao)最高, 且各坡向间差异显著, 均随土壤养分含量的增加明显增加, 说明LCCFDrao、LNCFDrao、LPCFDrao、LKCFDrao直接受土壤养分的影响, 而且北坡的土壤资源丰富, 植物具有更好的营养利用空间, 使得植物能够很好的生长, 植物性状的分布范围也相应的增加, 这与刘旻霞等[31]对甘南高寒草甸坡向梯度植物群落功能多样性的研究结果一致。
3.4 植物群落多元性状功能多样性对坡向的响应植物群落功能多样性是通过植物功能性状来反映植物在生态系统中的功能差异及其分布范围, 反映群落物种的生存策略及共存机制[29]。本研究发现, 从南坡到北坡功能丰富度显著增加(图 2), 这可能是由于土壤pH在南坡偏高, 土壤偏碱性, 且土壤pH是影响功能丰富度的主要环境限定因子(表 3), 而引起pH变化的因素很多, 比如土壤含水量, 它们是影响功能丰富度随坡向变化显著的主要因素。相关研究表明土壤pH与土壤含水量呈负相关, 即土壤水分的减少会引起土壤碱性增强, 使得植物的生长发育受到其生存环境的限制, 使得功能丰富度在南坡偏低。功能均匀度反映了物种功能性状分布的均匀程度[20]。本研究中功能均匀度在各坡向间的差异性不显著(图 1), 这与李谆[33]的研究结果一致, 尽管土壤pH对功能均匀度产生负效应, 但pH与土壤养分相比对功能均匀度解释能力较低, 不足以对其产生影响, 使得北坡和南坡物种功能性状分布均匀差异程度相近。功能离散度反映的是功能群或者植物功能性状在生态位空间的离散程度, 若某地区具有较高的功能离散度, 则说明该地区的资源利用效率高, 从而可以提高生态系统生产力, 进一步增强了植物对外界干扰的防御能力[31-32]。在本研究中南坡的功能离散度较低, 主要是因为南坡具有较高的土温和较低的土壤含水量, 而土壤含水量对功能离散度产生直接正效应, 土温产生负效应, 土壤含水量和土温是影响功能离散度的主要环境决定因子(表 4), 使得此坡向的物种丰富度降低, 生态位空间未得到充分的利用, 同时也限制了植物对资源的利用效率, 生态系统稳定性减弱[31]。总之, 功能多样性是生态系统功能最强的预测因子, 功能多样性越高, 其生态系统生产力也越高[20]。
3.5 植物群落物种多样性与功能多样性关系物种多样性与功能多样性是生态系统功能的重要基础, 阐明两者之间的相互关系, 对于揭示其对生态系统功能的相对作用具有重要的生态学意义[13]。本研究结果表明, 物种多样性与功能多样性间的关系存在显著差异(表 5)。功能丰富度与Margalef指数、Simpson指数呈显著正相关(P < 0.05), 且功能丰富度随着物种多样性的增加而增大, 这与我们的假设部分一致。这是因为功能丰富度主要衡量群落中的物种对生态位空间占据的大小, 若群落中的物种数目越多, 则功能性状分布的范围就越广, 因此物种占据的功能生态位也就越大, 这与许多学者的研究结果一致[34-35]。功能均匀度与物种多样性(Margalef、Simpson、Shannon、Pielou指数)无显著相关性, 这与我们的假设部分不一致, 但和以往的研究结果相一致[36]。一方面, 物种多样性与功能多样性的相关性分析发现, 功能均匀度指数具有相对独立性, 另一方面, 由功能多样性在不同坡向的变化趋势可知所有功能性状轴上物种分布均匀, 因此物种多样性的增加基本不会引起功能均匀度的变化。功能离散度与Margalef指数、Simpson指数、Shannon指数呈显著负相关(P < 0.05), 这是因为随着物种多样性的增加, 功能性状的范围会相应的增加, 但一定程度上缩小了性状差异。
本研究阐释了高寒草甸物种多样性与功能多样性在微生境梯度上的生物关系。植物群落物种多样性指数(Margalef、Simpson、Shannon及Pielou) 与功能多样性指数(FRic、FEve及FDis) 均在北坡偏高。物种多样性指数均与功能丰富度呈正线性相关(P < 0.05), 与功能离散度呈负线性相关(P < 0.05), 而与功能均匀度无相关关系。这体现了植物群落适应不同生境的生存策略, 在资源较为丰沛的北坡, 植物具有大而薄的叶片, 从而可以快速获取外部资源;相反, 南坡的资源比较贫瘠, 植物的生存策略转换为有效的资源保存即“慢回报”的方式。本研究结果也进一步证实了土壤养分、水分差异决定物种多样性, 而功能多样性的变化主要取决于物种多样性即由于环境筛选作用引起物种多样性的降低, 这将间接导致功能多样性降低;同时, 该研究结论也为甘南地区退化高寒草甸修复、生物多样性保护与管理提供理论依据。
| [1] |
Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741-758. DOI:10.1111/j.1461-0248.2006.00924.x |
| [2] |
张建贵, 王理德, 姚拓, 李海云, 高亚敏, 杨晓玫, 李昌宁, 李琦, 冯影, 胡彦婷. 祁连山高寒草地不同退化程度植物群落结构与物种多样性研究. 草业学报, 2019, 28(5): 15-25. |
| [3] |
向琳, 陈芳清, 官守鹏, 王玉兵, 吕坤. 井冈山鹿角杜鹃群落灌木层功能多样性及其随海拔梯度的变化. 生态学报, 2019, 39(21): 8144-8155. |
| [4] |
Cadotte M W, Cavender-Bares J, Tilman D, Oakley T H. Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity. PLoS One, 2009, 4(5): e5695. DOI:10.1371/journal.pone.0005695 |
| [5] |
Huang X B, Su J R, Li S F, Liu W D, Lang X D. Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Scientific reports, 2019, 9(1): 6979. DOI:10.1038/s41598-019-43475-1 |
| [6] |
Flynn D F B, Mirotchnick N, Jain M, Palmer M I, Naeem S. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships. Ecology, 2011, 92(8): 1573-1581. DOI:10.1890/10-1245.1 |
| [7] |
Flynn D F B, Gogol-Prokurat M, Nogeire T, Molinari N, Richers B T, Lin B B, Simpson N, Mayfield M M, DeClerck F. Loss of functional diversity under land use intensification across multiple taxa. Ecology Letters, 2009, 12(1): 22-23. DOI:10.1111/j.1461-0248.2008.01255.x |
| [8] |
Cadotte M W, Carscadden K, Mirotchnick N. Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 2011, 48(5): 1079-1087. DOI:10.1111/j.1365-2664.2011.02048.x |
| [9] |
秦大河. 三江源区生态保护与可持续发展. 北京: 科学出版社, 2014.
|
| [10] |
张中华, 周华坤, 赵新全, 姚步青, 马真, 董全民, 张振华, 王文颖, 杨元武. 青藏高原高寒草地生物多样性与生态系统功能的关系. 生物多样性, 2018, 26(2): 111-129. |
| [11] |
邓清月, 张晓龙, 牛俊杰, 秦浩. 晋西北饮马池山植物群落物种多样性沿海拔梯度的变化. 生态环境学报, 2019, 28(5): 865-872. |
| [12] |
Bai X H, Zhang J T. Functional diversity research of forest communities in the Xiaowutai Mountain National Nature Reserve, Hebei. Acta Ecologica Sinica, 2018, 38(2): 67-75. DOI:10.1016/j.chnaes.2017.05.003 |
| [13] |
董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472-1483. |
| [14] |
臧岳铭, 朱志红, 李英年, 王文娟, 席博. 高寒矮嵩草草甸物种多样性与功能多样性对初级生产力的影响. 生态学杂志, 2009, 28(6): 999-1005. |
| [15] |
孔彬彬. 刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响[D]. 西安: 陕西师范大学, 2016.
|
| [16] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000.
|
| [17] |
刘生涛. 皇甫川流域不同水土治理方式下土壤钾含量的变化. 北方环境, 2011, 23(3): 58-61. |
| [18] |
Laliberte E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. DOI:10.1890/08-2244.1 |
| [19] |
Mason N W H, Mouillot D, Lee W G, Wilson J B, Setälä H. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. DOI:10.1111/j.0030-1299.2005.13886.x |
| [20] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. DOI:10.1890/07-1206.1 |
| [21] |
刘旻霞. 甘南高寒草甸植物元素含量与土壤因子对坡向梯度的响应. 生态学报, 2017, 37(24): 8275-8284. |
| [22] |
陈洪磊, 欧阳炜, 吕凤玲, 宋艳暾, 郝蓉. 官渡河流域植被覆盖变化与地形因子相关性. 水土保持研究, 2019, 26(3): 135-140, 147-147. |
| [23] |
刘旻霞, 马建祖. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应. 应用生态学报, 2012, 23(12): 3295-3300. |
| [24] |
Reth S, Reichstein M, Falge E. The effect of soil water content, soil temperature, soil pH-value and the root mass on soil CO2 efflux-a modified model. Plant and Soil, 2005, 268(1): 21-33. DOI:10.1007/s11104-005-0175-5 |
| [25] |
Gong X Y, Brueck H, Giese K M, Zhang L, Sattelmacher B, Lin S. Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia, China. Journal of Arid Environments, 2008, 72(4): 483-493. DOI:10.1016/j.jaridenv.2007.07.001 |
| [26] |
南笑宁. 甘南高寒草甸不同坡向植物群落功能多样性研究[D]. 西北师范大学, 2020.
|
| [27] |
张倩, 孙小妹, 杨晶, 康宇坤, 姚宝辉, 苏军虎. 坡向对东祁连山高寒草甸群落物种功能群及其多样性的影响. 西北植物学报, 2019, 39(8): 1480-1490. |
| [28] |
刘哲, 李奇, 陈懂懂, 翟文婷, 赵亮, 徐世晓, 赵新全. 青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响. 生物多样性, 2015, 23(4): 451-462. |
| [29] |
朱云云, 王孝安, 王贤, 邓美皎. 坡向因子对黄土高原草地群落功能多样性的影响. 生态学报, 2016, 36(21): 6823-6833. |
| [30] |
葛兆轩, 孙国龙, 袁业, 黄选瑞, 张志东. 河北省森林草原区草本植物物种多样性和功能多样性. 草业学报, 2017, 26(7): 35-44. |
| [31] |
刘旻霞, 张国娟, 南笑宁, 宋佳颖, 蒋晓轩, 夏素娟. 甘南高寒草甸坡向梯度对植物群落功能多样性的影响. 西北植物学报, 2020, 40(8): 1414-1423. |
| [32] |
路兴慧, 臧润国, 丁易, 黄继红, 杨秀森, 周亚东. 抚育措施对热带次生林群落植物功能性状和功能多样性的影响. 生物多样性, 2015, 23(1): 79-88. |
| [33] |
李谆. 青藏高原高寒草甸植物群落功能性状及功能多样性对不同地形的响应[D]. 兰州: 兰州大学, 2016.
|
| [34] |
么旭阳, 胡耀升, 刘艳红. 长白山阔叶红松林典型森林群落功能多样性及其与地形因子的关系. 西北农林科技大学学报: 自然科学版, 2014, 42(10): 95-102. |
| [35] |
陈倩. 秦岭山地天然次生林群落物种多样性与功能多样性研究[D]. 杨凌: 西北农林科技大学, 2019.
|
| [36] |
盛芝露. 退化梯度上高山草甸植物群落功能多样性研究[D]. 昆明: 云南大学, 2015.
|