 2021, Vol. 41
2021, Vol. 41文章信息
- 韩豪, 骆漫, 李涛, 韦小丽
- HAN Hao, LUO Man, LI Tao, WEI Xiaoli
- 贵州闽楠天然种群特征、空间分布格局及空间关联分析
- Natural population characteristics, spatial distribution pattern and spatial correlation analysis of Phoebe bournei in Guizhou Province
- 生态学报. 2021, 41(13): 5360-5367
- Acta Ecologica Sinica. 2021, 41(13): 5360-5367
- http://dx.doi.org/10.5846/stxb201909201967
-
文章历史
- 收稿日期: 2019-09-20
- 修订日期: 2021-04-06





种群不仅是物种进化的基本单位也是群落组成的基本单位。在种群生态学研究领域中, 种群结构与种群空间格局已成为该领域研究的核心内容, 是种群重要的基本特征之一[1-2]。其中, 种群年龄结构是了解种群生存现状以及揭示其潜在生态适应机制的重要手段, 可以进一步推断出环境对种群的资源配置状况;生命表与存活曲线有助于预测种群在整个生活史中的演替趋势和掌握特定种群结构的发展规律;而种群空间格局在一定程度上可以反映出种群的生物、生态学特性以及种内、种间关系[3-5]。通过点格局分析方法, 前人分析了吉林长白山[6], 四川峨眉山[7], 西藏色季拉山[8], 北京东灵山[9]等地区的植物种群空间分布格局及其关联性。研究结果发现, 不同种群的树种在空间分布格局与年龄结构上会表现出不同的差异性[10-11], 同一树种也会因其生境, 发育阶段以及群落类型的不同其表现也会不相同[4, 12-13]。因此, 分析种群在生命过程中的数量特征、分布格局及变化规律, 对于濒危植物的保护、生存和生态恢复具有十分重要的科学意义[14]。
闽楠(Phoebe bournei), 别名楠木, 樟科(Lauraceae) 常绿乔木, 国家二级保护植物。因其材质优良, 纹理精美, 导致闽楠野生资源遭到大规模的偷窃和破坏, 使其种群资源数量快速下降。前人对于闽楠种群生态位[15-17]、种群遗传多样性[18-19]、群落学特征[20-21]等方面已有一些研究, 但对闽楠天然种群的年龄结构特征、空间分布、空间关联性等科学问题的研究鲜有报道, 而且研究地点主要集中在我国福建, 江西两省, 对于位于西南地区的贵州省闽楠天然种群的研究资料却未见报道。台江县拥有目前贵州省发现分布面积最大的闽楠天然群落, 面积达8 hm2。本研究以该天然群落为研究对象, 旨在通过对闽楠天然种群的年龄结构、空间分布格局以及空间关联性的研究, 揭示闽楠天然种群当前的数量动态, 阐明闽楠与环境之间的关系、受干扰状态, 预测其未来发展与演变趋势, 以期建立闽楠种群的适应性保护措施。
1 研究区概况研究区(东经108°20′—108°32′, 北纬26°37′—27°37′)位于贵州省黔东南州台江县境内, 海拔445—1980 m;土壤石砾含量高, 土壤类型以黄壤为主。样地所在属于中亚热带温和湿润气候, 年降水量847.2 mm, 日照时间长, 年均气温16.5 ℃, 雨水充足, 年均降水量1801.7 mm。该天然群落为当地少数民族保存的风水林, 并被划为县级保护区, 保护制度极为严格, 受干扰程度极小, 也未采取任何经营措施。闽楠群落总面积约8 hm2, 乔木层物种较为丰富, 主要有闽楠(Phoebe bournei)、丝栗栲(Castanopsis fargesii)、罗浮槭(Acer fabri)、南酸枣(Choerospondias axillaris)等树种;灌木层物种较少, 以贵州毛柃(Eurya kueichowensis)、栽秧泡(Rubus ellipticus)、白杜(Euonymus maackii)为主;草本层物种较单一, 主要是蕨类植物铁芒箕(Dicranopteris dichotoma)和里白(Hicriopteris glauca)。
2 研究方法 2.1 样地设置与野外调查在经实地踏查后, 选择该区域内闽楠分布集中、代表性强且保护较为完整的地段设置面积60 m×60 m的固定监测样地。将整个样地的东北角设为平面直角坐标系原点, 东西方向设为坐标系X轴, 南北方向设为Y轴, 对样地内的闽楠进行每木检尺并记录其(X, Y)坐标。把整个样地划分为4个30 m×30 m的大样方, 以10 m×10 m为调查单元, 调查记录乔木物种名、树高、胸径等信息;再以5 m×5 m为调查单元, 对每株更新苗(DGH<2.5 cm)编号, 详细调查其地径(DGH)、高度、枝下高等信息, 同时调查样方内海拔、坡度等立地因子(表 1)。
| 样方 Sample |
坡位 Slope positions |
坡向 Directions |
坡度 Slope angles/(°) |
海拔 Altitudes/m |
| S1 | 中坡位 | 阴坡 | 28—34 | 865 |
| S2 | 中坡位 | 阴坡 | 26—30 | 866 |
| S3 | 中下坡位 | 阴坡 | 29—35 | 921 |
| S4 | 中下坡位 | 阴坡 | 32—38 | 925 |
由于很难准确的判定天然濒危物种在不同生长阶段的年龄大小, 于是参照前人用闽楠胸径大小作为判断其年龄大小的方法[22-23]。将胸径2.5 cm以下的个体根据树高划分为:1—33 cm(龄级1)、33—100 cm(龄级2)、>100 cm(龄级3), 再将胸径2.5 cm以上的个体以胸径每增加5 cm设为一级, 以此类推将种群划分为8级, 最后根据划分的闽楠种群龄级结构将其定义为4个生长阶段:第1—3龄级为幼苗阶段、第4龄级为幼树阶段、第5—7龄级为中树阶段、第8龄级为大树阶段。
2.2.2 种群动态指数、生命表和存活曲线分析参照陈晓德[24]的动态量化方法分析种群动态变化指数。根据江洪[25]的方法编制闽楠种群静态生命表和存活曲线的相关参数。利用Hett等[26]的方法检验物种在整个生活史阶段存活状况。
2.2.3 种群空间分布格局分析以二维空间点的Ripley′s K函数为基础, 利用单变量函数Ripley′s L[27]建立不同径级的闽楠空间分布格局。其公式为:
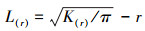
|
(1) |
式中, r表示空间尺度;K(r)表示从某一点起, 距离r内的期望点数;L(r)表示点格局与空间尺度r的关系, 下同。
当L(r)在径级1与径级2两条包迹线之上、两条包迹线之间、两条包迹线之下, 则分别代表植物种群呈聚集分布、随机分布、均匀分布。
2.2.4 种群不同生长阶段的空间分布格局分析考虑到不同径级之间植物个体在r内的数目, 通过双变量L(12)函数[28]探究径级1与径级2之间闽楠空间关联性, 其推演方法与(1)相同。其公式为:
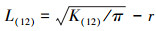
|
(2) |
式中, L(12)表示径级1与径级2之间的点格局与空间尺度r之间的关系;K(12)表示径级1和径级2之间的K(r)估计值。
若L(12)在径级1与径级2两条包迹线之上、两条包迹线之间、两条包迹线之下, 则分别代表径级1与径级2之间空间关联性呈正关联关系、无关联关系、负关联关系。
数据分析利用生态学软件ADE-4完成, 用1000次蒙特卡罗随机拟合计算出99%的置信区间。
3 结果与分析 3.1 闽楠种群年龄结构特征及动态变化分析由闽楠种群龄级结构图(图 1)可知, 闽楠种群的个体数量随着龄级的增加逐级下降, 年龄结构分布呈倒“J”型, 在整个生长阶段, 种群个体数量以第1、2龄级占比最高, 占总体数量的75.9%, 在后期阶段的6、7、8龄级个体数量分布最少, 分别仅占总体数量的2.4%、1.7%、1.1%。可见, 闽楠更新能力强, 有丰富的幼苗贮备资源, 将使闽楠种群更加稳定, 但后期呈现出一定的衰退趋势。

|
| 图 1 闽楠种群的龄级结构 Fig. 1 Age class structure of Phoebe bournei population 1:第1龄级first age class;2:第2龄级second age class;3:第3龄级third age class;4:第4龄级fourth age class;5:第5龄级fifth age class;6:第6龄级sixth age class;7:第7龄级seventh age class;8:第8龄级eighth age class |
种群结构随机干扰风险极大值P极大为0.06, 对外界环境会表现出较高的敏感性。但对闽楠种群8个龄级的动态量化分析发现, 各个年龄级Vn的计算结果都是正值, 再结合Vpi, V′pi的值分别为46.4%, 2.7%, 也为正值(表 2), 表明目前闽楠种群仍趋于稳定状态。
| 龄级 Age class |
种群动态指数 Dynamic index |
动态指数值/% Dynamic index value |
龄级 Age class |
种群动态指数 Dynamic index |
动态指数值/% Dynamic index value |
|
| 1 | V1 | 38.2 | 6 | V6 | 29.0 | |
| 2 | V2 | 66.2 | 7 | V7 | 37.0 | |
| 3 | V3 | 40.5 | Vpi | 46.4 | ||
| 4 | V4 | 42.6 | V′pi | 2.7 | ||
| 5 | V5 | 30.0 | P极大 | 0.06 | ||
| Vn:种群从n到n+1的数量动态变化指数Population dynamic change index from n to n+1;Vpi:无干扰时, 种群结构数量动态变化指数Dynamic change index of population structure when there is no interference;V′pi:考虑外部干扰时, 种群结构的数量动态变化指数Dynamic change index of population structure when considering external interference;P极大:随机干扰风险极大值Random Interference risk maxima | ||||||
闽楠种群的死亡率(qx)和消失率(kx)总体变化趋势基本一致(表 3)。不同龄级个体存活数量分布不同。第1、2龄级个体存活数最多, 能稳定进入第3龄级的幼苗个体存活数仅有第2龄级个体存活数的34%, 死亡率(qx)达到高峰期, 能够成功进入第8龄级的个体存活数仅有第1龄级个体存活数的2.2%。说明虽然闽楠幼苗更新能力较强, 幼苗存活数(lx)较多, 但随着年龄的增加, 可以稳定进入下一生长阶段的数量较少, 存活率显著降低。种群在第5龄级的生命期望(ex)最高, 在第8龄级ex降为0.5, 表明经过前期筛选存活下来的个体在第5龄级的生存条件好, 但随着后期物种对环境资源的利用效率降低、自然灾害、人为干扰等因素的作用ex开始下降。通过比较幂函数Nx=1400.263e-0.545x(R2=0.970)与指数函数Nx=1481.368x-1.892(R2=0.965)两种模型(图 2)的R值, 检验出闽楠种群的存活曲线更趋于DeeveyⅡ型。
| 龄级 Age class |
ax | lx | InIx | dx | qx | Lx | Tx | ex | Kx |
| 1 | 757 | 1000 | 6.908 | 382 | 0.382 | 809 | 1630 | 1.63 | 0.482 |
| 2 | 468 | 618 | 6.426 | 410 | 0.663 | 413 | 821 | 1.33 | 1.008 |
| 3 | 158 | 208 | 5.338 | 84 | 0.404 | 166 | 408 | 1.96 | 0.518 |
| 4 | 94 | 124 | 4.820 | 53 | 0.427 | 98 | 242 | 1.95 | 0.557 |
| 5 | 54 | 71 | 4.263 | 21 | 0.296 | 61 | 144 | 2.03 | 0.351 |
| 6 | 38 | 50 | 3.912 | 16 | 0.320 | 43 | 81 | 1.62 | 0.328 |
| 7 | 27 | 36 | 3.584 | 14 | 0.389 | 29 | 40 | 1.11 | 0.493 |
| 8 | 17 | 22 | 3.091 | — | — | 11 | 11 | 0.50 | — |
| ax:存活数Survivor;lx:标准化存活数The standardized survival;dx:标准化死亡数The standardized deaths;qx:死亡率Mortality;Lx:区间寿命Interval life;Tx:总寿命Total life;ex:生命期望Life expectancy;Kx:消失率Vanish rate | |||||||||

|
| 图 2 闽楠种群存活线曲线 Fig. 2 Survival curve of Phoebe bournei population |
根据种群不同发育阶段株树分布图(图 3)可知, 闽楠在各个年龄阶段的植株密度分布不同。密度由大到小依次为:幼苗、幼树、中树、大树。

|
| 图 3 闽楠种群不同发育阶段株树分布 Fig. 3 Density distribution of Phoebe bournei population at different development stages |
如图 4 L(r)函数分析结果所示, 处于幼苗阶段的闽楠在整个尺度上呈显著集群分布;幼树个体在0—27 m、28—29 m尺度上分别呈集群分布、随机分布的空间分布格局, 聚集强度比幼苗阶段有所下降;中树与大树在所有尺度上分别呈现出集群分布和随机分布的空间格局状态。总体看来, 随着龄级的增加, 闽楠种群空间分布格局由集群分布向随机分布转化。

|
| 图 4 闽楠种群不同发育阶段的空间分布点格局 Fig. 4 Plot pattern of different developmental stages of Phoebe bournei population 两条虚线为拟合的上下包迹线, 实线为L(r)函数 |
闽楠在不同生长阶段之间空间关联性表现不同(图 5)。在0—1 m, 4—29 m尺度上, 大树和幼苗呈正关联, 在其它尺度呈负关联;在0—2 m, 4—8 m和3 m, 9—29 m尺度上, 大树与幼树分别表现出正关联和负关联;在0—4 m尺度上, 大树与中树无关联, 5—29 m尺度上两者呈负关联;中树和幼苗、中树和幼树在0—29 m尺度上都呈负关联且随着空间尺度的增加关联性呈逐渐上升趋势;在0—6 m, 12—29 m尺度上, 幼树与幼苗呈负关联, 在10—11 m尺度, 两者表现出正关联。

|
| 图 5 闽楠种群不同发育阶段的空间关联点格局 Fig. 5 Spatial correlation point pattern of different developmental stages of Phoebe bournei population |
种群的年龄结构是对环境相适应的一种表现, 可以揭示种群未来发展和动态方向[29-30]。对于受保护的闽楠天然群落而言, 闽楠不同个体的实际年龄难以获得, 前人对闽楠天然种群的研究表明, 闽楠的胸径结构能够有效而合理地代表种群年龄结构[23, 31], 因此, 本研究用闽楠胸径大小判断其年龄大小。由闽楠种群年龄结构来看, 其种群年龄结构呈倒“J”型分布, 第1、2龄级(幼苗)植株密度最大, 但伴随年龄增加, 稳定进入后一阶段的个体存活率呈急剧下降的趋势, 这与桂亚可等[31]对江西闽楠种群结构的研究结果相似, 这可能是因为在幼龄阶段闽楠受各种环境因素的影响以及自身抗胁迫能力较弱, 并且随年龄增加, 对林下水分、养分、空间等资源的需求加大, 种内、种间竞争更加激烈, 严重制约幼龄个体成长为大龄个体。虽然闽楠种群对外界环境的敏感性高, 但考虑V′pi=2.7%和闽楠种群的存活曲线趋于Deevey-Ⅱ型, 说明目前闽楠种群处于稳定状态。种群在第5龄级的生命期望(ex)最高, 在第8龄级ex降到最低, 可能是种群个体在第5龄级的生存条件较好以及种群自身的抗性和生存能力增强, 但随着后期物种对环境的资源利用效率下降、自然灾害等因素导致ex开始下降。
4.2 不同生长阶段闽楠种群空间分布格局在本研究中, 闽楠随着龄级的增加, 闽楠种群空间分布格局由集群分布向随机分布转化。这与范忆等[32]对紫楠, 拓峰等[33]对青海云杉种群的研究结果相一致。一方面原因是, 在实际调查中发现, 整个生长阶段, 以幼苗阶段的分布数量最多, 种内竞争明显高于种间竞争, 同时, 该阶段的个体资源利用效率低、抗胁迫能力弱, 集群分布是提高自身竞争力, 更好适应外部环境, 维持种群稳定更新的最好方式。另一方面, 与闽楠种子自身传播特性有关, 闽楠千粒重(346±7.22) g[34], 质量较大, 以重力传播为主, 限制了种子的传播范围, 因此, 种子扩散限制成为种群内不同径级个体呈现聚集分布的主要原因之一[35]。随着后期植株个体对环境资源的需求量提高, 种内、种间竞争强度更加剧烈, 产生自疏效应, 同时, 人类盗伐、自然灾害等原因, 个体死亡率升高, 大树阶段的个体密度急剧降低, 于是种群格局逐渐趋于随机分布[33]。
4.3 不同生长阶段之间闽楠种群空间分布关联性不同生长阶段之间空间关联性表现有所差异。小尺度上, 大树与幼树呈负关联, 大尺度上, 两者呈正关联。在小尺度上, 是由于对空间资源的竞争;大尺度上, 可能是光照等环境因子的作用, 研究显示, 闽楠的光补偿点为16.53 μmol m-2 s-1[36], 表明该物种的耐荫能力较强, 适宜的遮荫环境有助于闽楠幼龄期个体生长发育, 降低高强度环境的胁迫作用。在小尺度上, 幼树与幼苗、大树与幼树都呈正关联, 大树和中树无关联, 前者一般是受物种自身生物学特性影响, 后者是因在大树和中树两个径级之间竞争强度减弱。在大尺度上, 大树和中树、幼树与幼苗、中树与幼苗以及中树和幼树都呈负关联且随着空间尺度的增加负关联性逐渐增强, 这是长期与环境相互作用产生的一种适应性策略, 径级小的个体通过集群作用提高抵御外界环境的胁迫能力、进而提高其生存能力, 保证小径级个体稳定更新, 径级大的个体则通过竞争限制径级小的个体获取空间、养分、光照等资源的数量, 径级小的个体存活率下降, 从而达到维持自身稳定发展[37]。因此, 不同生长阶段之间负关联性加强。
综上所述, 本研究得出结论:台江县天然闽楠种群年龄结构呈倒“J”型分布, 属于稳定型种群;随着龄级的增加, 分布格局由聚集分布转向随机分布, 且不同龄级植株在不同尺度上分布格局不同, 其不同龄级植株间的关系也因尺度不同而异。这是闽楠种子传播特性、生态适应性、种内种间竞争以及环境等多种因素相互影响的结果。闽楠种群目前虽然幼苗更新能力强, 短期内趋于稳定状态, 但最后能进入大龄阶段的个体数量严重不足, 导致后期会呈现衰退趋势。为了更好地保护台江县闽楠天然群落, 有针对性地采取有效措施, 应深入探讨闽楠与环境之间的关系, 特别是与光照的关系, 揭示闽楠植株耐荫性随光环境和年龄变化的规律及其动态响应特征, 找出不同年龄阶段闽楠适生的光环境阈值。在此基础上对闽楠种群采取科学的人工保护措施, 修枝、砍伐一定数量的竞争木, 清除对象木附近杂草、灌木, 为其提供合适的生存空间, 提高闽楠种群个体存活率, 让小径级个体稳定进入林冠层。
| [1] |
张婕, 上官铁梁, 段毅豪, 郭微, 刘卫华, 郭东罡. 灵空山辽东栎种群年龄结构与动态. 应用生态学报, 2014, 25(11): 3125-3130. |
| [2] |
张志祥, 刘鹏, 蔡妙珍, 康华靖, 廖承川, 刘春生, 楼中华. 九龙山珍稀濒危植物南方铁杉种群数量动态. 植物生态学报, 2008, 32(5): 1146-1156. DOI:10.3773/j.issn.1005-264x.2008.05.019 |
| [3] |
Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercus garryana) in British Columbia, Canada. Forest Ecology and Management, 2000, 137(1/3): 209-219. |
| [4] |
涂洪润, 李娇凤, 刘润红, 梁士楚, 兰泽南, 章欣仪, 康馨丹, 姜勇. 桂林岩溶石山檵木种群空间格局及其关联性. 应用生态学报, 2019, 30(8): 2621-2630. |
| [5] |
Druckenbrod D L, Shugart H H, Davies I. Spatial pattern and process in forest stands within the Virginia piedmont. Journal of Vegetation Science, 2005, 16(1): 37-48. DOI:10.1111/j.1654-1103.2005.tb02336.x |
| [6] |
安璐, 吴兆飞, 范春雨, 张春雨, 赵秀海. 长白山次生杨桦林种群空间点格局及密度制约效应. 生态学报, 2021, 41(4): 1461-1471. |
| [7] |
秦爱丽, 马凡强, 许格希, 史作民, 陈其勇. 珍稀濒危树种峨眉含笑种群结构与动态特征. 生态学报, 2020, 40(13): 4445-4454. |
| [8] |
任毅华, 罗大庆, 周尧治, 方江平, 卢杰. 西藏色季拉山急尖长苞冷杉原始林粗木质残体空间格局分析. 生态学报, 2019, 39(21): 8048-8057. |
| [9] |
马芳, 王顺忠, 冯金朝, 桑卫国. 北京东灵山优势种群树木死亡对空间格局与生境的影响. 生态学报, 2018, 38(21): 7669-7678. |
| [10] |
舒兰, 刘兆刚, 董灵波. 帽儿山天然次生林内主要木本植物空间格局及更新特点. 应用生态学报, 2019, 30(6): 1945-1955. |
| [11] |
胡满, 曾思齐, 龙时胜. 青冈栎次生林主要树种空间分布格局及其关联性研究. 中南林业科技大学学报, 2019, 39(6): 66-71. |
| [12] |
刘旻霞, 连依明, 李文. 微地形对优势种群点格局和关联性的影响. 应用生态学报, 2018, 29(5): 1569-1575. |
| [13] |
吴初平, 袁位高, 盛卫星, 黄玉洁, 陈庆标, 沈爱华, 朱锦茹, 江波. 浙江省典型天然次生林主要树种空间分布格局及其关联性. 生态学报, 2018, 38(2): 537-549. |
| [14] |
解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 2014, 34(15): 4272-4279. |
| [15] |
吴大荣. 罗卜岩保护区闽楠等优势植物种群竞争研究初步. 南京林业大学学报, 1998, 22(3): 35-38. |
| [16] |
林建勇, 李俊福, 何应明, 唐复呈, 李娟, 梁瑞龙. 人为干扰(采集)对闽楠群落优势种群生态位的影响. 广西林业科学, 2020, 49(1): 60-65. DOI:10.3969/j.issn.1006-1126.2020.01.012 |
| [17] |
张俊钦. 福建明溪闽楠天然林主要种群生态位研究. 福建林业科技, 2005, 32(3): 31-35. DOI:10.3969/j.issn.1002-7351.2005.03.008 |
| [18] |
李娟, 董利军, 林建勇, 刘雄盛, 梁瑞龙. 闽楠不同地理种群遗传多样性的ISSR分析. 分子植物育种, 2019, 17(23): 7822-7828. |
| [19] |
江香梅, 温强, 叶金山, 肖复明, 江梅. 闽楠天然种群遗传多样性的RAPD分析. 生态学报, 2009, 29(1): 438-444. DOI:10.3321/j.issn:1000-0933.2009.01.052 |
| [20] |
苏良佺. 闽楠群落物种多样性与乔木层主要树种生态位研究. 林业勘察设计, 2017, 37(4): 39-43. |
| [21] |
邱生龙. 明溪县闽楠群落分布特征研究. 安徽农业科学, 2017, 45(24): 153-154, 160-160. DOI:10.3969/j.issn.0517-6611.2017.24.051 |
| [22] |
何亚平, 费世民, 蒋俊明, 陈秀明, 张旭东, 何飞. 不同龄级划分方法对种群存活分析的影响——以水灾迹地油松和华山松种群生存分析为例. 植物生态学报, 2008, 32(2): 448-455. DOI:10.3773/j.issn.1005-264x.2008.02.024 |
| [23] |
吴大荣, 朱政德. 福建省罗卜岩自然保护区闽楠种群结构和空间分布格局初步研究. 林业科学, 2003, 39(1): 23-30. DOI:10.3321/j.issn:1001-7488.2003.01.004 |
| [24] |
陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. DOI:10.3321/j.issn:1000-0933.1998.02.017 |
| [25] |
江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992: 7-13.
|
| [26] |
Hett J M, Loucks O L. Age structure models of balsam fir and eastern hemlock. Journal of Ecology, 1976, 64(3): 1029-1044. DOI:10.2307/2258822 |
| [27] |
Nguyen H, Wiegand K, Getzin S. Spatial patterns and demographics of Streblus macrophyllus trees in a tropical evergreen forest, Vietnam. Journal of Tropical Forest Science, 2014, 26(3): 309-319. |
| [28] |
Diggle P J, Besag J, Gleaves J T. Statistical analysis of spatial point patterns by means of distance methods. Biometrics, 1976, 32(3): 659-667. DOI:10.2307/2529754 |
| [29] |
李先琨, 苏宗明, 向悟生, 宁世江, 唐润琴, 欧祖兰, 李瑞棠. 濒危植物元宝山冷杉种群结构与分布格局. 生态学报, 2002, 22(12): 2246-2253. DOI:10.3321/j.issn:1000-0933.2002.12.031 |
| [30] |
Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres Del Paine national park, Chile. Vegetatio, 1992, 98(1): 13-22. DOI:10.1007/BF00031633 |
| [31] |
桂亚可, 潘萍, 欧阳勋志, 臧颢, 国瑞, 李杨. 赣中闽楠天然种群数量特征及分布格局. 生态学杂志, 2019, 38(10): 2918-2924. |
| [32] |
范忆, 楼一恺, 库伟鹏, 戴其林, 王铮屹, 赵明水, 余树全. 天目山紫楠种群年龄结构与点格局分析. 浙江农林大学学报, 2020, 37(6): 1027-1035. |
| [33] |
拓锋, 刘贤德, 刘润红, 赵维俊, 敬文茂, 马剑, 武秀荣, 赵晶忠, 马雪娥. 祁连山大野口流域青海云杉种群空间格局及其关联性. 植物生态学报, 2020, 44(11): 1172-1183. DOI:10.17521/cjpe.2020.0177 |
| [34] |
陈建毅. 闽楠人工林物候观测研究. 安徽农业科学, 2014(20): 6742-6743. DOI:10.3969/j.issn.0517-6611.2014.20.081 |
| [35] |
郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄俞淞, 黄甫昭, 李冬兴, 李先琨. 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析. 生物多样性, 2015, 23(2): 183-191. |
| [36] |
黄宇, 林智勇, 汤行昊, 廖鹏辉, 张娟, 范辉华. 不同造林模式闽楠造林成效研究. 山地农业生物学报, 2019, 38(5): 6-12, 29-29. |
| [37] |
韩豪. 闽楠天然林种群特征及其天然更新研究[D]. 贵州: 贵州大学, 2020.
|