 2021, Vol. 41
2021, Vol. 41文章信息
- 蔡建堤, 徐春燕, 马超, 叶孙忠, 庄之栋, 刘勇, 谢少卿, 沈长春
- CAI Jiandi, XU Chunyan, MA Chao, YE Sunzhong, ZHUANG Zhidong, LIU Yong, XIE Shaoqing, SHEN Changchun
- 闽南渔场游泳动物优势种种群聚集特性
- Population aggregation characteristics of the main dominant species of nekton in Minnan Fishing Ground
- 生态学报. 2021, 41(12): 4818-4826
- Acta Ecologica Sinica. 2021, 41(12): 4818-4826
- http://dx.doi.org/10.5846/stxb202001070052
-
文章历史
- 收稿日期: 2020-01-07
- 修订日期: 2021-05-24





2. 福建省海洋生物增养殖与高值化利用重点实验室, 厦门 361013
2. Key Laboratory of Cultivation and High-value Utilization of Marine Organisms in Fujian Province, Xiamen 361013, China
闽南渔场是福建海区重要渔场, 为鱼类、虾类、蟹类和头足类等游泳动物的产卵、索饵与越冬的重要场所[1]。近年来, 由于环境污染、气候变化和渔业资源的过度捕捞等造成闽南渔场渔业资源不断衰退[2], 但有的种类资源衰退明显, 如大黄鱼(Larimichthys crocea)[3], 而有的种类资源衰退程度相对较轻, 如二长棘犁齿鲷(Evynnis cardinali)[4], 造成不同种类资源衰退程度出现分化现象的原因多种多样, 如个体发育阶段、生理状况、生活习性等[5-6]。其中, 生活习性是种群资源变动的主要因素之一, 而种群集群行为是生活习性最基本的要素, 通过研究闽南渔场游泳动物优势种种群聚集的变化, 有助于揭示闽南渔场渔业资源渔业变动因素, 了解鱼类洄游及鱼群形成机制[6], 对于确定种群特征、种群间相互关系以及种群与环境之间的关系具有非常重要的作用。
有关鱼群聚集特性的研究, 已有学者开展大量的研究工作[7-11], 主要研究方法有实验室水槽观测法、潜水观测法、标志放流法、超声波影像分析法、数学模拟和仿真法。用数学模型进行模拟逐渐成为一种帮助认识鱼群行为内在规律的有效手段, 蔡建堤等[12-13]利用格局强度指标分析了闽东北外海假长缝拟对虾(Parapenaeus fissuroides)和中华管鞭虾(Solenocera crassicornis)的种群聚集特性, 认为种群分布格局强度适用于研究海洋游泳动物聚集特性, 这种集群行为的研究方法开辟了自然海域海洋游泳动物集群行为研究的新思路。目前对闽南渔场鱼群聚集状况不甚了解, 缺乏系统化研究。为此, 本文应用格局强度指标分析了闽南渔场主要优势种聚集特性, 比较不同的类群、种类和季节种群聚集状况, 探讨影响集群的主要因子, 旨在揭露闽南渔场游泳动物优势种种群聚集的特征、内在规律和形成机理, 为闽南渔场渔业资源的科学研究和管理提供依据。
1 材料与方法 1.1 材料2016年4月和10月, 在闽南渔场开展了游泳动物渔业资源的专项调查, 共设10个调查渔区(图 1)。调查船为闽东渔61792底拖网船(渔船吨位51 t, 主机功率183 kW, 网口宽度10.0 m, 网口高度5.5 m, 囊网网目1.8 cm)。渔业资源调查按《海洋渔业资源调查规范》(SC/T 9403—2012)进行, 每航次在每个调查渔区拖曳一次, 拖曳时间均为1 h, 拖速2.0 kn。环境因子(水深、温度、盐度、PH和溶解氧)调查按《海洋监测规范》(GB 17378.1—2007)进行。本次调查取全部渔获物, 带回福建省水产研究所生态实验室进行种类鉴定和生物学测定, 对每个种类分别进行称重和计数。物种鉴定主要参考《福建鱼类志(上、下卷)》、《中国动物志(头足纲)》、《中国海洋动物图集(第六册)》、《南海对虾类等》。生物拉丁文名与世界海洋生物种记录(WoRMS)进行核对。

|
| 图 1 调查站位图 Fig. 1 Survey station map 图中数字为渔区的编号, ★为调查站点 |
优势种的计算采用相对重要性指数[14]。
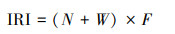
|
IRI表示游泳动物在群落中的相对重要性指数, N为某一种类的尾数占总尾数的百分比, W为一种类的重量占总重量的百分比, F为一种类的出现样方数占总样方数的百分比。当IRI值大于1000时将该物种定义为优势种;当IRI值介于100—1000时将该物种定义为重要种。
1.2.2 Lloyd的平均拥挤度指数(m*)[15]
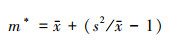
|
m*为平均拥挤度指数, 表示每个个体的平均拥挤程度。平均拥挤度越大, 则个体的平均拥挤程度越大。
1.2.3 Cassie指标[16-17]
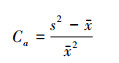
|
Ca为Cassie指标, 用来表示种群的聚集强度, s2为各个渔区CPUE的方差, x为平均CPUE。Ca值愈大, 则种群聚集强度愈强。
1.2.4 聚块指数[18-21]
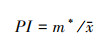
|
PI为聚块指数, 表示斑块组成形式。PI较大, 则代表主要种群数量集中分布于少数团聚的大斑块, 种群聚集度较高, PI值较小, 则代表主要种群数量分散分布于多数斑块, 种群聚集度较低。
1.2.5 Mrisita指数(I)[22]
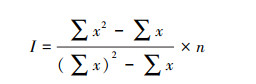
|
I值为Mrisita指数, 表示种群聚集强度, n为渔区数量, x为渔区CPUE。I值越大, 种群聚集强度越大。
2 结果 2.1 游泳动物优势种根据2016年4、10月在闽南渔场游泳动物渔业资源的调查资料, 由相对重要性指数计算不同类群的优势种, 游泳动物优势种共17种, 其中鱼类有6种, 分别为六指马鲅(Polydactylus se-xfilis)、带鱼(Trichiurus lepturus)、蓝圆鲹(Decapterus maruadsi)、叫姑鱼(Johnius grypotus)、多鳞鱚(Sillago sihama)、二长棘犁齿鲷;虾类有3种, 分别为哈氏仿对虾(Parapenaeopsis hardwickii)、鹰爪虾(Trachypenaeus curvirostris)、须赤虾(Metapenaeopsis barbata);蟹类有4种, 分别为拥剑梭子蟹(Portunus gladiator)、日本蟳(Charybdis japonica)、三疣梭子蟹(P. trituberculatus)、善泳蟳(Charybdis natator);头足类有4种, 分别为火枪乌贼(Loligo beaka)、柏氏四盘耳乌贼(Euprymna morsei)、短蛸(Octopus ocellatus)、杜氏枪乌贼(L. duvaucelii)。
2.2 鱼类优势种的种群聚集特性春季, 六指马鲅的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其余依次为带鱼、蓝圆鲹、叫姑鱼、多鳞鱚, 而二长棘犁齿鲷的种群聚集强度最低。二长棘犁齿鲷的平均拥挤度最高, 其余依次为六指马鲅、带鱼、蓝圆鲹, 而叫姑鱼和多鳞鱚均较低(图 2)。

|
| 图 2 春季闽南渔场鱼类主要优势种聚集特性 Fig. 2 Population aggregation characteristics of dominant species of fishes in Minnan Fishing Ground in spring 二长棘犁齿鲷(Evynnis cardinali), 叫姑鱼(Johnius grypotus), 蓝圆鲹(Decapterus maruadsi), 带鱼(Trichiurus lepturus), 六指马鲅(Polydactylus se-xfilis),多鳞鱚(Sillago sihama) |
秋季, 六指马鲅的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其余依次为多鳞鱚、叫姑鱼、带鱼、二长棘犁齿鲷, 而蓝圆鲹的种群聚集强度最弱。带鱼的平均拥挤度最高, 其余依次为六指马鲅、二长棘犁齿鲷、多鳞鱚、叫姑鱼、蓝圆鲹(图 3)。

|
| 图 3 秋季闽南渔场鱼类主要优势种聚集特性 Fig. 3 Population aggregation characteristics of dominant species of fishes in Minnan Fishing Ground in autumn |
春季, 哈氏仿对虾的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其次为鹰爪虾、须赤虾。鹰爪虾的平均拥挤度明显最高, 其余依次为哈氏仿对虾、须赤虾(图 4)。

|
| 图 4 春季闽南渔场虾类主要优势种聚集特性 Fig. 4 Population aggregation characteristics of dominant species of shrimps in Minnan Fishing Ground in spring 须赤虾(Metapenaeopsis barbata), 哈氏仿对虾(Parapenaeopsis hardwickii), 鹰爪虾(Trachypenaeus curvirostris) |
秋季, Cassie指标、聚块指数和Mrisita指数以鹰爪虾最高, 种群聚集强度最强, 其次为须赤虾, 而哈氏仿对虾最低。鹰爪虾的平均拥挤度最高, 其余依次为须赤虾、哈氏仿对虾(图 5)。

|
| 图 5 秋季闽南渔场虾类主要优势种聚集特性 Fig. 5 Population aggregation characteristics of dominant species of shrimps in Minnan Fishing Ground in autumn |
春季, 拥剑梭子蟹的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其次为日本蟳、三疣梭子蟹、善泳蟳。三疣梭子蟹的平均拥挤度最高, 其余依次为日本蟳、善泳蟳、拥剑梭子蟹(图 6)。

|
| 图 6 春季闽南渔场蟹类主要优势种聚集特性 Fig. 6 Population aggregation characteristics of dominant species of crabs in Minnan Fishing Ground in spring 拥剑梭子蟹(Portunus gladiator),日本蟳(Charybdis japonica),三疣梭子蟹(P. trituberculatus), 善泳蟳(Charybdis natator) |
秋季, 日本蟳的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其余依次为三疣梭子蟹、善泳蟳、拥剑梭子蟹。拥剑梭子蟹的平均拥挤度最高, 其余依次为日本蟳、善泳蟳、三疣梭子蟹(图 7)。

|
| 图 7 秋季闽南渔场蟹类主要优势种聚集特性 Fig. 7 Population aggregation characteristics of dominant species of crabs in Minnan Fishing Ground in autumn |
春季, 火枪乌贼的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其次为柏氏四盘耳乌贼、短蛸、杜氏枪乌贼。火枪乌贼的平均拥挤度最高, 其余依次为杜氏枪乌贼、短蛸, 柏氏四盘耳乌贼明显最低(图 8)。

|
| 图 8 春季闽南渔场头足类主要优势种聚集特性 Fig. 8 Population aggregation characteristics of dominant species of cephalopods in Minnan Fishing Ground in spring 杜氏枪乌贼(Loligo duvaucelii),短蛸(Octopus ocellatus), 柏氏四盘耳乌贼(Euprymna morsei), 火枪乌贼(Loligo beaka) |
秋季, 火枪乌贼的Cassie指标、聚块指数和Mrisita指数最高, 种群聚集强度最强, 其余依次为杜氏枪乌贼、柏氏四盘耳乌贼、短蛸。杜氏枪乌贼的平均拥挤度明显最高, 其余依次为短蛸、火枪乌贼、柏氏四盘耳乌贼(图 9)。

|
| 图 9 秋季闽南渔场头足类主要优势聚集群特性 Fig. 9 Population aggregation characteristics of dominant species of cephalopods in Minnan Fishing Ground in autumn |
种群聚集强度是度量一个种群分布格局的聚集程度, 是渔业资源科学管理和利用的基础。种群聚集形成的斑块形状、大小和间隙等不尽相同, 斑块反映的生态信息十分复杂, 聚集现象及其内在机制不能依靠单个的指标来解释。聚块指数主要以种群主要数量所占据的斑块的数量衡量种群聚集强度[12-13], 如春季, 80%的叫姑鱼种群数量集中在3个渔区, 聚块指数为2.40, 秋季, 82%的叫姑鱼种群数量集中在1个渔区, 聚块指数较高, 为6.84, 种群聚集强度较强。Cassie指标主要根据种群在单一斑块聚集度判断种群聚集强度[12-13], 如春季, 38%的叫姑鱼种群数量集中在1个渔区, Cassie指标为1.40, 而秋季, 82%的叫姑鱼种群数量集中在1个渔区(283渔区), Cassie指标较高, 为5.84, 种群聚集程度较强。Mrisita指数由渔区数量和各个渔区CPUE两个参数组成, 其值不受均值影响[12-13]。因此, Cassie指标适用于分析和比较种群在单一斑集中程度, 聚块指数适合于衡量种群聚集形成斑块的特征, Mrisita指数不受样方数的影响, 更适合比较样方数不同的种群聚集强度。闽南渔场游泳动物优势种同一类群中不同种类分布的样方数变化不大, 平均CPUE受样方数的影响比较小, 因此, 三者研究相同类群中不同种类的种群聚集特性结果基本一致(图 1—图 9)。而不同类群间分布的样方数往往差别大, 平均CPUE受样方数多寡影响较大, 因此三者研究不同类群聚集强度的结果有时并不一致。本研究应用聚块指数、Cassie指标和Mrisita指数分析不同类群聚集强度结果不同。如秋季, 鱼类、蟹类和头足类的聚块指数分别为5.75、4.73和5.50, 种群聚集强度呈现出鱼类大于蟹类和头足类(图 10), 而Mrisita指数分别为3.33、4.43和4.97, Cassie指标分别为3.24、3.77和4.46, 种群聚集强度呈现出鱼类小于蟹类和头足类(图 11, 图 12)。头足类、虾类和蟹类团聚在一个渔区的种群数量比例大, 且分布渔区数量较少, 而鱼类分布渔区数量较多, 且主要种群数量分布于多数渔区。因此, Cassie指标和Mrisita指数更适合于分析闽南渔场头足类、虾类和蟹类的种群聚集强度, 聚块指数比较适合于分析闽南渔场鱼类的聚集强度。其中, Mrisita指数不受均值影响, 用于比较类群间的聚集强度更为恰当。

|
| 图 10 闽南渔场不同类群的聚块指数 Fig. 10 The Poly block index of different faunas in Minnan Fishing Ground |

|
| 图 11 闽南渔场不同类群的Cassie指标 Fig. 11 The Cassie index of different faunas in Minnan Fishing ground |

|
| 图 12 闽南渔场不同类群的Mrisita指数 Fig. 12 The Mrisita index of different faunas in Minnan Fishing Ground |
平均拥挤度是指每个个体在一个样方中的平均个体数, 反映个体平均拥挤程度[12-13]。个体所经历的种内竞争强度并不真的由种群密度决定, 而是取决于邻近个体造成拥挤程度或者对个体的抑制程度[23]。贝根[24]研究种内竞争对死亡率的影响, 认为种群初始密度的增加会增大种群死亡率, 死亡率是密度依赖性的。春季, 二长棘犁齿鲷幼鱼群体数量众多, 渔获个体共6020尾, 个体平均拥挤度较高, 为5819.78, 种内竞争激烈, 增大了幼鱼群体死亡率, 只有少部分幼鱼个体成长为大个体, 秋季二长棘犁齿鲷的个体比较大, 种群数量明显较少, 渔获个体共202尾。可见, 平均拥挤度可体现种群内部竞争程度, 对于研究种内竞争有重要参考价值。
个体平均拥挤程度越高, 种群数量越多, 渔获渔业资源量越丰富。平均拥挤度和渔获资源量关系为:
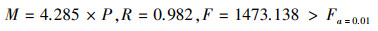
|
式中M为渔获物种的渔业资源量(g), P为平均拥挤度, R为相关系数。平均拥挤度越大, 则渔业资源越丰富, 而平均拥挤度越小, 则渔业资源越少。如2015年春季, 二长棘犁齿鲷种群的平均拥挤度指数为5819.78, 秋季, 二长棘犁齿鲷种群的平均拥挤度指数为2808.14, 而1998年春季, 平均拥挤度指数为24543.25, 秋季, 平均拥挤度指数为4330.17。二长棘犁齿鲷渔业资源2015年比1998年有明显下降。可见, 平均拥挤度在渔业资源开发和渔业管理等方面具有实用价值。
一般情况下, 平均拥挤度越大, 种群聚集强度越强, 而平均拥挤度越小, 种群聚集强度越弱(图 5)。但平均拥挤度的大小并不一定总与种群聚集强度的强弱一致, 如杜氏枪乌贼为闽南渔场重要的经济种类, 春季, 种群聚集强度最弱, 而平均拥挤度却最大(图 8);拥剑梭子蟹是闽南渔场重要经济种类, 春季, 种群聚集强度最强, 而平均拥挤度却最小(图 6)。表明平均拥挤度和种群聚集强度是衡量种群聚集特性的不同指标, 其生态学意义不同, 平均拥挤度主要体现渔业资源密度和种群竞争状况, 而种群聚集强度主要反映种群聚集程度。
3.3 闽南渔场游泳动物优势种聚集特性从类群角度分析, Mrisita指数分析表明:春季, 种群聚集强度以蟹类较强, 其余依次为虾类、头足类和鱼类, 秋季, 虾类的种群聚集强度最强, 其余依次为头足类、蟹类和鱼类(图 12)。虾类、头足类和鱼类的种群聚集强度秋季大于春季, 而蟹类的聚集强度秋季小于春季。蟹类的种群聚集强度不同季节出现明显差异, 春季, 分布范围小, 比较集中, 聚集强度强, 而秋季分布范围广, 聚集强度弱(图 12)。
蟹类聚集强度的分析结果符合闽南渔场蟹类分布特征[25]。总体上, 虾类和蟹类的聚集强度大于鱼类、头足类, 主要原因是甲壳类活动速度慢、空间较小, 从而表现出聚集强度较强的特征。春季, 鱼类、蟹类、头足类和虾类的平均拥挤度分别为1428.75、310.89、280.42和56.47, 平均拥挤度以鱼类最高, 资源量最丰富, 其次为蟹类、头足类和虾类(图 13)。秋季, 头足类、鱼类、蟹类和虾类的平均拥挤度分别为4186.14、2101.88、1704.85和274.62, 平均拥挤度以头足类最大, 资源量最丰富, 其余依次为鱼类、蟹类、虾类(图 13)。总体上, 闽南渔场虾类资源量最小, 鱼类资源量比较丰富, 蟹类和头足类的资源量秋季大于春季, 闽南渔场渔业资源量秋季明显大于春季。不同类群平均拥挤度的研究结果符合闽南渔场单船拖网作业渔获物的组成特征[26]。

|
| 图 13 闽南渔场不同类群的平均拥挤度 Fig. 13 The mean crowding of different faunas in Minnan Fishing Ground |
从种类角度分析, 本文分别采用上述3种聚集强度指标分析了闽南渔场17个种类(6种鱼类、4种蟹类、4种头足类和3种虾类)种群聚集强度结果基本一致, 说明3种聚集强度指标均可用于分析和比较同一类群中不同种类的聚集强度。春季, 拥剑梭子蟹是聚集强度最强的种类, 其次为六指马鲅、火枪乌贼。秋季, 鹰爪虾为聚集强度最强的种类, 其次为六指马鲅、火枪乌贼。秋季的杜氏枪乌贼是平均拥挤度最高的种类, 渔业资源量最高, 其次为秋季的带鱼和二长棘犁齿鲷。表明杜氏枪乌贼、带鱼和二长棘犁齿鲷是闽南渔场秋季主捕的经济种类。
种群聚集特性由种群本身的生境条件、生物学特性以及二者之间的相互作用所决定[27-28]。生殖活动是影响种群聚集强度的重要因素。如拥剑梭子蟹繁殖高峰期为2—4月, 集中繁殖[29], 因而春季种群聚集强度最强。生活习性也会影响种群聚集强度。如日本蟳属沿岸定居性种类, 分布范围小, 主要分布于沿海岛礁区及浅海水域[2], 呈现出种群聚集强度较大, 蟹类中日本蟳秋季聚集强度最大。环境条件对种群聚集特性有重要影响。如鹰爪虾主要分布在45—65m水深的海域, 鹰爪虾在盐度33—34的海域分布数量较多[29], 种群分布比较密集, 呈现出种群聚集强度较大, 秋季鹰爪虾有较多分布[30], 资源丰富, 在虾类中平均拥挤度最大。季节变化也会对平均拥挤度有较大的影响, 秋季比春季鱼类、虾类、蟹类和头足类优势种的平均拥挤程度均较高。
集群动能也用于自然海域鱼类集群行为的研究[5], 根据1998年闽南-台湾浅滩渔场开展二长棘犁齿鲷资源的专项调查资料, 采用集群动能研究二长棘犁齿鲷的集群性, 认为春季幼鱼群体集群性较强, 而秋季二长棘犁齿鲷群体分散。采用格局强度指标分析二长棘犁齿鲷的聚集特性, 1998年春季, 幼鱼种群的Cassie指标和平均拥挤度指数分别为1.28和24543.25, 而秋季二长棘犁齿鲷种群的Cassie指标和平均拥挤度指数分别为0.31和4330.17, 说明春季比秋季种群聚集强度更强, 平均拥挤度更高。可见, 春季幼鱼群体集群性较强, 而秋季二长棘犁齿鲷群体分散。集群动能和格局强度指标研究种群集群性原理不同, 但二者研究结果一致, 符合二长棘犁齿鲷群体结构特征[31-32], 进一步佐证了应用格局强度指标研究种群聚集特性是科学的。与集群动能相比, 格局强度指标有明确的生态学意义, 能细致地反映种群聚集强度、斑块的组成形式、个体平均拥挤度、种群内部的竞争程度和种群扩散程度等生态学信息, 是一种帮助认识鱼群集群行为内在规律和研究渔场形成机制的有效手段。
| [1] |
林裕江, 陈文勇. 闽南渔场渔业资源利用现状和管理对策. 现代渔业信息, 2000, 15(2): 6-7. |
| [2] |
沈长春, 蔡建堤, 戴天元, 刘勇, 马超, 徐春燕, 庄之栋. 福建海区渔业资源可持续利用. 厦门: 厦门大学出版社, 2018, 1. |
| [3] |
徐开达, 刘子藩. 东海区大黄鱼渔业资源及资源衰退原因分析. 大连水产学院学报, 2007, 22(5): 392-396. DOI:10.3969/j.issn.1000-9957.2007.05.015 |
| [4] |
叶孙忠. 闽南-台湾浅滩二长棘鲷群体数量的时空分布. 福建水产, 2004, 4: 36-39. |
| [5] |
蔡建堤, 马超, 姜双城, 吴建绍, 蔡玉婷, 刘勇. 闽南-台湾浅滩渔场二长棘鲷群体集群行为宏观量化与分析. 水生生物学报, 2013, 37(2): 185-190. |
| [6] |
周应祺, 王军, 钱卫国, 曹道梅, 张仲秋, 柳玲飞. 鱼类集群行为的研究进展. 上海海洋大学学报, 2013, 22(5): 734-743. |
| [7] |
赵建. 鱼群集群行为的建模与仿真. 太原: 太原科技大学, 2008. |
| [8] |
徐盼麟. 基于视频的鱼类行为自动观测系统[D]. 上海: 上海海洋大学, 2011.
|
| [9] |
H Yoshioka. Mathematical analysis and validation of an exactly solvable model for upstream migration of fish schools in one-dimensional rivers. Mathematical Biosciences, 2016, 281: 139-148. DOI:10.1016/j.mbs.2016.09.014 |
| [10] |
Wu C J, Wang L. Numerical simulations of self-propelled swimming of 3D bionic fish school. Science in China Series E: Technological Sciences, 2010, 52(3): 658-669. |
| [11] |
Paramo J, Bertrand S, Villalobos H, Gerlotto F. A three-dimensional approach to school typology using vertical scanning multibeam sonar. Fisheries Research, 2007, 84(2): 171-179. DOI:10.1016/j.fishres.2006.10.023 |
| [12] |
蔡建堤, 徐春燕, 叶孙忠, 庄之栋, 马超, 刘勇, 陈梅芳, 沈长春. 闽东北海域假长缝拟对虾种群分布格局强度. 生态学报, 2017, 37(17): 5790-5797. |
| [13] |
蔡建堤, 徐春燕, 马超, 刘勇, 庄之栋, 陈洁, 沈长春. 闽东北海域中华管鞭虾种群聚集特性. 生态学报, 2017, 37(6): 1844-1850. |
| [14] |
刘惠, 俞存根, 郑基, 邓小艳, 张平, 颜文超, 谢旭. 杭州湾灰鳖洋海域秋季主要游泳动物生态位与种间联结. 生态学报, 2019, 39(11): 3828-3836. |
| [15] |
Lloyd M. 'Mean crowding'. Journal of Animal Ecology, 1967, 36(1): 1-30. DOI:10.2307/3012 |
| [16] |
Cassie R M. Frequency distribution models in the ecology of plankton and other organisms. Animal Ecol, 1962, 31: 65-92. DOI:10.2307/2333 |
| [17] |
陈艳华, 龙岳林, 彭重华. 湖南省阳明山山顶苔藓矮林优势种空间分布格局. 生态学报, 2019, 39(18): 6690-6700. |
| [18] |
覃林. 统计生态学. 北京: 中国林业出版社, 2009, 58-59. |
| [19] |
兰国玉, 雷瑞德. 植物种群空间分布格局研究方法概述. 西北林学院学报, 2003, 18(2): 17-21. DOI:10.3969/j.issn.1001-7461.2003.02.006 |
| [20] |
杨持. 植物种群分布格局的强度与纹理. 内蒙古大学学报(自然科学版), 1988(04): 662-668. |
| [21] |
武玉珍, 张峰. 山西桑干河流域湿地植被优势种群分布格局研究. 植物研究, 2006, 26(6): 735-741. DOI:10.3969/j.issn.1673-5102.2006.06.019 |
| [22] |
Morisita M. Measuring the dispersion of individuals and analysis of the distributional patterns. Mem. Fac. Sci. Kyusha Univ. Ser. E(Biol.), 1959, 2: 215-235. |
| [23] |
边疆晖. 根田鼠种群密度制约扩散的生态学过程及遗传效应[D]. 杭州: 浙江大学, 2005.
|
| [24] |
贝根, 汤森, 哈珀. 生态学-从个体到生态系统. 李博, 张大勇, 王德华, 译. 4版. 北京: 高等教育出版社, 2016: 144.
|
| [25] |
叶孙忠. 闽南、台湾浅滩渔场蟹类种类组成及分布特征. 海洋渔业, 2004, 26(4): 249-254. |
| [26] |
沈长春. 闽南-台湾浅滩渔场单船拖网作业调查与分析. 渔业研究, 2012, 34(4): 302-308. |
| [27] |
李圣法, 程家骅, 严利平. 东海大陆架鱼类群落的空间结构. 生态学报, 2007, 27(11): 4377-4386. DOI:10.3321/j.issn:1000-0933.2007.11.001 |
| [28] |
余为, 陈新军. 西北太平洋柔鱼栖息地环境因子分析及其对资源丰度的影响. 生态学报, 2015, 35(15): 5032-5039. |
| [29] |
黄美珍. 福建海区拥剑梭子蟹、红星梭子蟹. 和锈斑虫寻的食性与营养级研究台湾海峡, 2004, 23(2): 159-161. |
| [30] |
宋海棠, 俞存根, 姚光. 东海鹰爪虾的数量分布和变动. 海洋渔业, 2004, 26(3): 184-188. |
| [31] |
叶孙忠. 闽南-台湾浅滩二长棘鲷的生长特性. 水产学报, 2004, 28(6): 663-668. |
| [32] |
叶孙忠, 肖方森, 陈文勇. 闽南-台湾浅滩二长棘鲷群体结构特征. 福建水产, 2004, 1: 23-30. |