 2021, Vol. 41
2021, Vol. 41文章信息
- 钱灵颖, 黄智洵, 杨盛昌, 曹文志
- QIAN Lingying, HUANG Zhixun, YANG Shengchang, CAO Wenzhi
- 厦门市重点保护植物空间优先保护格局研究
- Study on spatial conservation priority pattern of key protected plants in Xiamen
- 生态学报. 2021, 41(11): 4367-4378
- Acta Ecologica Sinica. 2021, 41(11): 4367-4378
- http://dx.doi.org/10.5846/stxb202101110110
-
文章历史
- 收稿日期: 2020-11-11
- 修订日期: 2021-05-14

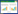




为应对生物多样性的迅速丧失, 我国通过划定一定数量的自然保护地对重点珍稀保护动植物、生物多样性及其所能够提供的生态系统服务进行有效地保护。截至2019年底, 我国共建立超过1.18万个自然保护地, 保护面积占全国国土面积的22.1%[1]。虽然我国自然保护地面积不断增加, 生物多样性下降趋势得到有效遏制, 但徐卫华[2]、杨朝辉[3]、孙思琦[4]等分别对长江经济带的国家级自然保护区、贵州省现有保护地体系、中国东南部自然保护区的生物多样性保护效率的研究结果表明, 大多数自然保护地在发展的早期阶段缺乏科学评估, 导致已有自然保护地体系存在对少数生物保护利用价值不高的物种或生境提供过多的保护, 或与生物多样性受到严重威胁的区域之间存在空间失配问题, 促进了空间保护优先化的发展。
空间保护优先化是一种基于明确的保护目标确定优先保护区网络的定量技术, 能够为保护或环境规划问题生成空间优先级, 为空间保护规划提供决策依据[5]。目前已经开发了多种软件可用于设计生物多样性空间优先保护格局[6]。使用较为广泛的空间保护优先化软件, 包括能够生成使生物多样性保护价值最大化的规划单元组合的Zonation, 使用最小集方法来确定以几乎最小成本实现保护目标的规划单元组合的MARXAN[7]以及通过计算各规划单元在实现保护目标上的不可替代性来确定空间保护优先性的C-Plan[8]。
相较于其它空间保护优先化定量技术, Zonation输出的空间优先保护格局旨在识别出多个物种的核心分布区, 实现生物多样性保护价值最大化, 维持景观的连通性, 而不是获得“以最小成本达到保护目标”的解决方案[9-10]。Zonation模型成功运行首先需要获得物种的生态位信息[11]。生态位模型能够根据物种实际分布及环境变量计算物种生态位, 目前应用较为广泛的生态位模型, 包括最大熵模型(MaxENT)、Bioclim模型和Domain模型[12-13], 每种生态位模型都可以独立预测物种的潜在分布区, 但具有各自的偏好性。Bioclim是最简单的生态位模型, 但模型预测精度与物种分布数据呈正相关, 并且无法处理环境因素之间的相互作用[14], Domain模型能够处理不连续的物种分布点位信息, 但对于每个潜在地点只利用一个点位信息来确定物种分布适宜性, 准确性不高[15], 而MaxENT模型能够在样本量较小、环境变量之间相关性不明的情况下, 通过探寻在物种实际分布点位和环境变量约束下最大熵的可能分布, 直接生成适宜物种生存的连续的物种潜在分布图, 具有所需样本量小、预测结果准确、兼容性好等优势[16-19]。目前MaxENT和Zonation的结合使用已经被广泛应用于划定以保护鸟类[20]、植物[21]、淡水鱼类[22]、海洋生物[23]以及地方特色物种[24]生物多样性为目标的多物种保护规划, 在多物种的空间优先保护区识别中发挥着重要的作用。
城市生物多样性是保证城市生态系统功能持续性的基础, 城市森林多样性可以提高生态系统健康的供给弹性[25]。厦门市快速的城市化导致城市人工环境不断增加, 森林植被遭到严重破坏, 原生森林生态系统逐渐退化[26], 近年来营造的人工森林植被结构单一, 不利于生物多样性形成。此外, 作为海湾型城市, 厦门市曾经拥有广阔的滩涂面积, 非常适合红树林的生长, 然而大规模的围海造田、填海造陆使滨海红树植物遭受破坏, 红树林面积日益减少, 城市生态系统健康受到威胁。本研究以保护厦门市重点保护植物的生物多样性为目标, 使用MaxENT模型作为物种分布模型模拟厦门市重点保护植物的潜在分布, Zonation模型作为空间保护优先化的定量工具, 获得该地重点保护植物的理论空间优先保护格局, 分析厦门市目前生物多样性保护地建设的空缺, 为厦门市生物多样性保护提供决策依据, 促进城市发展的稳定性和可持续性(图 1)。

|
| 图 1 研究流程 Fig. 1 Research process AUC:受试者操作特性曲线ROC(Receiver Operating Characteristic)曲线下方的面积大小Area Under Curve; SSI物种:特殊利益物种Species of Special Interest |
厦门市地处福建省东南部, 是东南沿海重要的中心城市, 总面积1700.61 km2。其中, 陆地面积1699.39 km2, 海域面积约390 km2。地形以滨海平原、台地和丘陵为主, 属于亚热带海洋性季风气候, 年平均气温约20.9℃; 雨量适中, 年降水量1143.5 mm, 降水集中于4-9月, 平均相对湿度较高, 全年适宜植物生长[27]。其位于闽粤沿海植被区, 共有常绿针叶林、常绿阔叶林、针阔叶混交林、红树林等11种植被类型, 马尾松林、杉木林、木荷林、白骨壤群落、秋茄群落、桐花树+白骨壤群落等62个群系, 南亚热带季风常绿阔叶林为厦门市的地带性植被[28]。长期以来, 厦门市的自然生态系统受到很大程度地扰动, 红树林面积日益减少, 森林植被次生性明显, 城市绿地景观多样性指数低, 各绿地类型分布不均匀, 生态功能较弱, 生物多样性保护空间缺乏。
1.2 重点保护植物的选择本研究参考《国家重点保护野生植物名录(第一批)》、《国家重点保护野生植物名录(第二批)》、《福建省国家和省重点保护野生植物目录》(http://lyj.fujian.gov.cn)、《厦门市地方志》(www.fzb.xm.gov.cn)所列保护植物种类, 剔除重复选项, 并根据中国植物主题数据库(http://www.plant.csdb.cn)中所记录的保护植物分布地, 中国数字植物标本馆(https://www.cvh.ac.cn)及教学标本资源共享平台(http://mnh.scu.edu.cn/)的标本信息, 筛选出在厦门市存在并具地理分布信息的39种植物作为本研究中的重点保护植物(表 1)。包括苏铁(Cycas revoluta)、台湾苏铁(Cycas taiwaniana)、水杉(Metasequoia glyptostroboides)、伯乐树(Bretschneidera sinensis)4种国家一级保护野生植物, 樟树(Cinnamomum camphora)、格木(Erythrophleum fordii)、穗花杉(Amentotaxus argotaenia)、福建柏(Fokienia hodginsii)、刺桫椤(Alsophila spinulosa)、金毛狗(Cibotium barometz)6种国家二级保护野生植物, 黄樟(Cinnamomum porrectum)、江南油杉(Keteleeria fortunei)、木榄(Bruguiera gymnorhiza)等8种省重点保护野生植物, 凤凰木(Delonix regia)、细枝叶下珠(Phyllanthus leptoclados)、秋茄(Kandelia candel)等21种地方重点保护野生植物。其中, 秋茄、木榄、白骨壤及桐花树作为红树植物, 所构成的红树林生态系统在保障本地海岸带生物多样性及生态安全等方面起到重要作用[29]。
| 序号 Number |
植物名称 Plants |
首次潜在分布区预测范围 Prediction range of the first potential distribution area |
AUC值 AUC Value |
最终是否视为SSI物种 Considered SSI species |
Zonation权重设置 Weight setting |
序号 Number |
植物名称 Plants |
首次潜在分布区预测范围 Prediction range of the first potential distribution area |
AUC值 AUC Value |
最终是否视为SSI物种 Considered SSI species |
Zonation权重设置 Weight setting |
|
| 1 | 樟树Cinnamomum camphora | 福建 | 0.767 | 否 | 7 | 21 | 松叶耳草Hedyotis pinifolia | 福建 | 0.920 | 否 | 2 | |
| 2 | 格木Erythrophleum fordii | 福建 | 0.987 | 否 | 7 | 22 | 苏铁Cycas revoluta | 厦门 | 0.807 | 否 | 10 | |
| 3 | 江南油杉Keteleeria fortunei | 福建 | 0.782 | 否 | 4 | 23 | 台湾苏铁Cycas taiwaniana | 厦门 | 0.812 | 否 | 10 | |
| 4 | 油杉 Keteleeria fortunei |
福建 | 0.786 | 否 | 4 | 24 | 白桂木 Artocarpus hypargyreus |
厦门 | 0.795 | 否 | 4 | |
| 5 | 长苞铁杉 Tsuga longibracteata |
福建 | 0.845 | 否 | 4 | 25 | 木榄 Bruguiera gymnorhiza |
厦门 | 0.825 | 否 | 6 | |
| 6 | 穗花杉 Amentotaxus argotaenia |
福建 | 0.945 | 否 | 7 | 26 | 马尾松 Pinus massoniana |
厦门 | 0.799 | 否 | 2 | |
| 7 | 青钱柳 Cyclocarya paliurus |
福建 | 0.774 | 否 | 4 | 27 | 细枝叶下珠 Phyllanthus leptoclados |
厦门 | 0.780 | 否 | 2 | |
| 8 | 红锥 Castanopsis hystrix |
福建 | 0.853 | 否 | 4 | 28 | 皱叶忍冬 Lonicera rhytidophylla |
厦门 | 0.784 | 否 | 2 | |
| 9 | 福建含笑 Michelia fujianensis |
福建 | 0.758 | 否 | 4 | 29 | 米槠Castanopsis carlesii | 厦门 | 0.765 | 否 | 2 | |
| 10 | 甜槠Castanopsis eyrei | 福建 | 0.728 | 否 | 2 | 30 | 秋茄Kandelia candel | 厦门 | 0.771 | 否 | 6 | |
| 11 | 黄樟Cinnamomum porrectum | 福建 | 0.715 | 否 | 4 | 31 | 白骨壤Avicennia marina | 厦门 | 0.881 | 否 | 6 | |
| 12 | 凤凰木Delonix regia | 福建 | 0.948 | 否 | 2 | 32 | 车桑子Dodonaea viscosa | 厦门 | 0.882 | 否 | 2 | |
| 13 | 台湾相思Acacia confusa | 福建 | 0.899 | 否 | 2 | 33 | 蓟罂粟Argemone mexicana | - | - | 是 | 2 | |
| 14 | 木麻黄 Casuarina equisetifolia |
福建 | 0.809 | 否 | 2 | 34 | 草海桐 Scaevola sericea |
- | - | 是 | 2 | |
| 15 | 伯乐树 Bretschneidera sinensis |
福建 | 0.724 | 否 | 10 | 35 | 水杉 Metasequoia glyptostroboides |
- | - | 是 | 10 | |
| 16 | 福建柏Fokienia hodginsii | 福建 | 0.64 | 是 | 7 | 36 | 鸡爪簕Oxyceros sinensis | - | - | 是 | 2 | |
| 17 | 刺桫椤 Alsophila spinulosa |
福建 | 0.738 | 否 | 7 | 37 | 黄瑞木 Adinandra yunnanensis |
- | - | 是 | 2 | |
| 18 | 金毛狗Cibotium barometz | 福建 | 0.441 | 是 | 7 | 38 | 柃木Eurya japonica | - | - | 是 | 2 | |
| 19 | 桐花树 Aegiceras corniculatum |
福建 | 0.987 | 否 | 6 | 39 | 木荷Schima superba | - | - | 是 | 2 | |
| 20 | 檵木Loropetalum chinense | 福建 | 0.617 | 是 | 2 | |||||||
| AUC:受试者操作特性曲线ROC(Receiver Operating Characteristic)曲线下方的面积大小Area Under Curve; SSI物种:特殊利益物种Species of Special Interest; - 表示该植物在MaxENT模型模拟物种潜在分布阶段即作为SSI物种, 无AUC值 | ||||||||||||
物种实际地理分布点位信息来自近10年的野外调研工作以及中国数字植物标本馆(https://www.cvh.ac.cn)、教学标本资源共享平台(http://mnh.scu.edu.cn/)、世界生物多样性信息机构(Global Biodiversity Information Facility, GBIF)、相关文献所记录的近60年植物标本信息。涉及到仅提供采集地点信息的标本通过百度地图的坐标拾取系统(http://api.map.baidu.com/lbsapi/getpoint/index.html/)将地名转化为点位信息。利用ArcGIS 10.5对重点保护植物坐标进行筛选, 1 km× 1 km范围内仅保留相同物种的一个点位。研究共获得39种植物的有效坐标。
1.3.2 环境变量研究初步选取19种有关温度和降水的生物气候因子和地形因子数据(海拔、坡度、坡向)作为影响重点保护植物分布的环境变量(表 2)。其中19种生物气候因子与植物生长密切相关, 被认为是进行物种潜在分布模拟的典型环境变量[30], 从WorldClim全球气候数据库(http://www.worldclim.org/)下载当前气候条件下最高分辨率的生物气候因子数据(30"(约1 km)), 通过克里金插值法将分辨率提高至30 m; 从地理空间数据云(http://www.gscloud.cn/)上下载DEM数据(30 m), 获得地形因子数据。使用SPSS软件进行Pearson相关性分析(表 3), 最终选择相关系数小于0.8的14种生物气候因子和3种地形因子构建模型(表 2)[31]。
| 环境变量 Variable |
描述 Description |
是否用于建模 Whether to use for modelling |
||
| 生物气候因子 | Bioclimatic1 | 年均温 | 是 | |
| Bioclimatic factors | Bioclimatic2 | 平均温度日较差 | 是 | |
| Bioclimatic3 | 等温性 | 否 | ||
| Bioclimatic4 | 温度季度变化 | 是 | ||
| Bioclimatic5 | 最热月最高温 | 是 | ||
| Bioclimatic6 | 最冷月最低温 | 是 | ||
| Bioclimatic7 | 年温度变化范围 | 是 | ||
| Bioclimatic8 | 最湿季度均温 | 是 | ||
| Bioclimatic9 | 最干季度均温 | 是 | ||
| Bioclimatic10 | 最暖季度均温 | 否 | ||
| Bioclimatic11 | 最冷季度均温 | 是 | ||
| Bioclimatic12 | 年平均降水量 | 否 | ||
| Bioclimatic13 | 最湿月降水量 | 否 | ||
| Bioclimatic14 | 最干月降水量 | 是 | ||
| Bioclimatic15 | 降水量变异系数 | 是 | ||
| Bioclimatic16 | 最湿季度降水量 | 否 | ||
| Bioclimatic17 | 最干季度降水量 | 是 | ||
| Bioclimatic18 | 最暖季度降水量 | 是 | ||
| Bioclimatic19 | 最冷季度降水量 | 是 | ||
| 地形因子 | dem | 海拔 | 是 | |
| Topographic factors | Pox | 坡向 | 是 | |
| Slope | 坡度 | 是 | ||
| 相关系数 Correla-tion coeffici-ent |
Bio1 | Bio2 | Bio3 | Bio4 | Bio5 | Bio6 | Bio7 | Bio8 | Bio9 | Bio10 | Bio11 | Bio12 | Bio13 | Bio14 | Bio15 | Bio16 | Bio17 | Bio18 | Bio19 |
| Bio1 | 1 | ||||||||||||||||||
| Bio2 | 0.032 | 1 | |||||||||||||||||
| Bio3 | -0.320 | 0.801 | 1 | ||||||||||||||||
| Bio4 | 0.629 | 0.337 | -0.183 | 1 | |||||||||||||||
| Bio5 | 0.665 | 0.263 | -0.123 | 0.730 | 1 | ||||||||||||||
| Bio6 | 0.679 | -0.304 | -0.600 | 0.595 | 0.598 | 1 | |||||||||||||
| Bio7 | 0.446 | 0.661 | 0.262 | 0.694 | 0.569 | 0.146 | 1 | ||||||||||||
| Bio8 | 0.469 | -0.223 | -0.347 | 0.247 | 0.387 | 0.598 | 0.013 | 1 | |||||||||||
| Bio9 | 0.708 | 0.071 | -0.285 | 0.658 | 0.696 | 0.643 | 0.496 | 0.435 | 1 | ||||||||||
| Bio10 | -0.337 | 0.093 | 0.336 | -0.402 | -0.418 | -0.292 | -0.280 | 0.004 | -0.434 | 1 | |||||||||
| Bio11 | 0.707 | 0.012 | -0.325 | 0.637 | 0.656 | 0.763 | 0.391 | 0.527 | 0.687 | -0.259 | 1 | ||||||||
| Bio12 | -0.809 | -0.044 | 0.365 | -0.698 | -0.704 | -0.764 | -0.513 | -0.468 | -0.742 | 0.456 | -0.803 | 1 | |||||||
| Bio13 | -0.759 | -0.030 | 0.358 | -0.699 | -0.711 | -0.810 | -0.455 | -0.525 | -0.698 | 0.351 | -0.796 | 0.900 | 1 | ||||||
| Bio14 | -0.343 | 0.084 | 0.323 | -0.388 | -0.409 | -0.272 | -0.278 | 0.008 | -0.427 | 0.945 | -0.240 | 0.435 | 0.337 | 1 | |||||
| Bio15 | -0.681 | -0.235 | 0.074 | -0.554 | -0.591 | -0.480 | -0.535 | -0.169 | -0.564 | 0.053 | -0.535 | 0.569 | 0.600 | 0.065 | 1 | ||||
| Bio16 | -0.767 | -0.048 | 0.305 | -0.688 | -0.828 | -0.764 | -0.488 | -0.492 | -0.744 | 0.508 | -0.765 | 0.871 | 0.864 | 0.484 | 0.587 | 1 | |||
| Bio17 | -0.676 | 0.074 | 0.424 | -0.614 | -0.601 | -0.687 | -0.396 | -0.446 | -0.650 | 0.619 | -0.684 | 0.828 | 0.877 | 0.602 | 0.364 | 0.797 | 1 | ||
| Bio18 | -0.709 | -0.082 | 0.326 | -0.745 | -0.706 | -0.687 | -0.578 | -0.270 | -0.749 | 0.542 | -0.685 | 0.808 | 0.783 | 0.514 | 0.624 | 0.806 | 0.731 | 1 | |
| Bio19 | 0.353 | 0.573 | 0.338 | 0.491 | 0.461 | 0.092 | 0.589 | 0.176 | 0.310 | 0.032 | 0.316 | -0.368 | -0.389 | 0.033 | -0.372 | -0.419 | -0.274 | -0.270 | 1 |
| Bio1-19:表 2中所列的生物气候因子Bioclimatic1-19 | |||||||||||||||||||
Maxent模型根据最大熵原理, 基于该物种已知出现地的环境条件预测物种在研究区域内的所有可能潜在分布地[32]。
2.1.1 模型运行研究使用MaxENT模型3.4.1版本预测厦门市重点保护植物的潜在分布区。将所获得的厦门市重点保护植物有效坐标超过5个的12种物种, 直接输入MaxENT模型中获得厦门市重点保护植物潜在分布[33-35]。对于所获得的厦门市植物样本量小于5的20种物种, 检索物种在整个福建省范围内的分布信息, 计算获得福建省潜在物种分布图后截取厦门市范围的物种潜在分布[36]。福建省检索到的点位数据依然小于5时, 将物种视为特殊利益物种(Species of Special Interest, SSI species), 不再进行物种潜在分布预测。最初便视为SSI物种的共有7种, 这些物种的点位信息将以.txt文本形式直接输入Zonation模型[37-38]。
MaxENT模型每次预测随机选择75%的物种分布点数据作为训练集用于建立模型, 25%的物种分布点数据作为随机测试数据集用于模型验证[39], 设置正则化乘数为1[40], 采集分析底图的10000个数据点进行模拟, 重复运行次数为4, 重复次数运行类型选择Crossvalidate, 建立模型。共进行两轮物种潜在分布区预测, 通过jackknife刀切法分析各个环境变量对物种分布的重要性[41-42], 在第一次模型运行结束后, 剔除对物种贡献率低于1.0%的环境因子[43], 进行二次模拟, 最终获得植物在厦门市的潜在分布栅格图作为空间保护优先化所需的物种分布状态信息。
2.1.2 模型准确性评估使用受试者操作特性曲线ROC(Receiver Operating Characteristic)评估模型模拟结果准确性[44]。ROC曲线下与坐标轴围成的面积AUC(Area Under Curve)越接近1.0, 检测方法真实性越高。根据AUC值将模型预测结果准确性分为5类, AUC<0.6, 预测失败; AUC=0.6-0.7, 较差; AUC=0.7-0.8, 一般; AUC=0.8-0.9, 良好; AUC=0.9-1.0, 优秀[45]。本研究中认为只有当AUC值大于0.7时, MaxENT模型输出的植物潜在分布栅格图才可输入Zonation模型中。而AUC值小于0.7的物种, 被视为SSI物种。
2.2 Zonation模型Zonation以迭代的方式产生由互补性驱动的景观保护等级, 模型首先假设整体景观都得到保护, 接着逐步识别并移除边际损失最小的单元, 将保护价值最高的区域保留到最后, 获得多物种理论空间优先保护格局[46]。Zonation能够综合考虑多物种的潜在分布核心区, 并识别对于维持景观连通性非常重要的区域, 为划定优先保护区以维持较高的生物多样性水平提供一种定量方法。
2.2.1 单元去除规则选择Zonation算法是否有意义完全取决于对边际损失的定义, 而边际损失是由不同的单元去除规则(Removal Rule)决定[47]。研究选择核心单元分区(Core-area Zonation, CAZ)作为单元去除规则, 得到的空间优先保护格局不在物种之间进行取舍, 而是保护所有物种的核心分布区域[11]。
2.2.2 物种权重设置物种特征权重会影响单元从景观中移除的顺序以及单元移除过程中任意位置上所保留的物种分布比例。因此在对多物种进行空间保护优先化前需确定其所属的保护类别, 并在此基础上判断物种的全球稀有性、地方性以及经济社会价值等, 从而赋予相应的权重[48](表 4)。
| 植物保护类别Protection category | 权重设置Weight setting |
| 国家一级保护野生植物National key protected wild plants grade Ⅰ | 10 |
| 国家二级保护野生植物National key protected wild plants grade Ⅱ | 7 |
| 红树植物Mangrove | 6 |
| 省重点保护野生植物Provincial key protected wild plants | 4 |
| 地方重点保护植物Local key protected wild plants | 2 |
将重点保护植物的潜在分布栅格图及SSI植物的点数据输入Zonation模型中, 设置相应的权重, 识别理论上既适宜重点保护植物生存又能够保证景观连通性的区域, 获得本地重点保护植物的景观保护等级以及一组特征曲线。
根据2020年全球生物多样性目标, 设置景观优先等级前17%的区域作为厦门市重点保护植物空间优先保护区[49]。此外, 由于本研究设定的目标之一是保护本地重点保护植物不至灭绝, 因此依据Zonation模型生成的随景观丧失物种加权灭绝风险曲线将移除后可能会导致物种的灭绝风险迅速增大的景观单元进一步视为一级保护区。
3 结果 3.1 模型准确性评估使用MaxENT模型模拟除蓟罂粟、草海桐等7种SSI物种外的其它重点保护植物的潜在分布, 得到每一物种4次重复运行下的平均AUC值(表 1)。两次MaxENT模拟潜在分布结束后, 福建柏、金毛狗、檵木的AUC值依然小于0.7, 因此也被视为SSI物种。最后筛选得的SSI物种共10种, 获得的GIS栅格图层共29个。
3.2 空间优先保护格局将29个重点保护植物的潜在分布栅格图及10个SSI植物的点数据输入Zonation模型中, 获得本地重点保护植物的空间优先保护格局以及随景观丧失物种加权灭绝风险曲线。
随着景观丧失比例的增加, 物种加权灭绝风险随之增加。景观丧失比例低于92%时, 物种加权灭绝风险增长率相对稳定, 当景观丧失比例达到92%时, 物种的平均灭绝风险达到39%。此后, 物种加权灭绝风险锐增至100%, 物种迅速灭绝(图 2)。结合2020年全球生物多样性目标, 共选取研究区域总面积的17%作为重点保护植物的优先保护区, 将保护等级最高的8%区域划为一级保护区, 保护等级在8%-17%范围内的区域划为二级保护区(图 3)。

|
| 图 2 随景观丧失物种加权灭绝风险曲线 Fig. 2 Weighted extinction risk curve with landscape loss |

|
| 图 3 厦门市重点保护植物重点保护区划分 Fig. 3 Division of key protected plant areas in Xiamen |
厦门市重点保护植物的一级保护区主要集中分布在厦门岛内的东部和南部、岛外东南部以及西南部的海岸带区域、西北部的山地。二级保护区主要分布在一级保护区的外围。叠加厦门市现有自然保护地, 结果表明大部分的自然保护地集中分布在厦门市的中部地区, 少部分位于西部山区和海岸带地区, 本研究所划定的厦门市重点保护植物理论空间优先保护区基本尚未得到良好的保护。
4 讨论 4.1 重点保护植物优先保护区分布特点(1) 地形因子为影响植物生长分布的主导环境因子。MaxENT模型通过jackknife刀切法可判断影响各物种分布的主导环境因子, 结果表明地形因子对植物生长分布的影响均大于生物气候因子, 其中海拔为对物种潜在分布概率影响最大的环境因子。主要是由于厦门市受海洋的影响较为显著, 气温年较差较小, 气候温和稳定, 全年均适宜植物生长, 但地势由西北向东南倾斜, 海拔较高的西北部可能会导致植物存在逆境保护机制来保护其免受光损伤[50], 因而重点保护植物优先保护区主要集中于厦门市的东南部, 西北部的优先保护区分布面积较小。
(2) 优先保护区多集中于海岸带区域。厦门市是我国典型的海湾型城市, 其海岸线长达234公里, 海沧湾、马銮湾、杏林湾、同安湾、翔安湾等构成了厦门海岸带, 研究表明重点保护植物优先保护区主要分布在海沧湾以及翔安湾, 其间分布着红树林、河口、沙滩等重要生态系统, 蕴含着丰富的自然资源, 既是植被生长的适宜栖息地, 也是鸟类越冬的良好场所, 具有较高的生态系统服务价值, 为丰富生物多样性提供可能。
(3) 二级保护区分布于一级保护区外围。Zonation模型生成的厦门市重点保护植物空间优先区间具有较好的连通性, 将重点保护植物优先保护区进一步划分为一级保护区和二级保护区, 大部分的二级保护区分布于一级保护区的外围, 类似于目前自然保护区中的核心区、缓冲区, 因此一级保护区内的保护要求应严于二级保护区, 以禁止进一步开发利用为主。
4.2 重点保护植物优先保护区用地现状将研究所得的重点保护植物优先保护区与厦门市现有自然保护地建设情况、厦门市生态功能区规划(1998-2030年)、厦门市土地利用规划(2006-2020年)和厦门市城市总体规划(2011-2020年)对比, 探究优先保护区的生态空间保护利用情况。
(1) 厦门岛外西部、北部的优先保护区得到较好保护。厦门市的森林生态系统主要集中分布于岛外的西部、北部、东部地区, 共同特点为开发利用强度较低。其中, 西部、北部的优先保护区属西北部低山丘陵水土保持与生态林生态功能区与西部低山丘陵水土保持与生态林生态功能小区范畴, 包括了小坪森林公园-莲花国家森林公园, 该区域生物多样性得到较好保护, 以生态保育为主要发展方向。
(2) 海岸带区域的优先保护区被大量占用。快速城市化进程导致有限的土地资源难以承受人口快速增长带来的压力, 促使人类不断地将生态空间转为城镇空间。厦门岛外的西南部及东南部、岛内的东部及南部海岸带的优先保护区均被开发为居住用地、商业服务用地、物流仓储用地和区域交通建设用地等, 适宜植物生长的栖息地被建设用地所占用, 导致厦门东西海域水质下降, 陆地和海洋的生物多样性下降明显, 但在目前的自然保护地建设中将海岸带优先保护区划入其中的仅有五缘湾栗喉蜂虎自然保护区和五缘湾湿地公园, 总体看来海岸带区域的优先保护区保护空缺较大。
(3) 厦门岛外东南部部分优先保护区虽然未被占用, 但规划中属发展备用地, 生态保护不足。相对岛内而言, 岛外东南部的优先保护区生态空间占用程度较低, 留有部分空间作为发展备用地, 为城市未来的发展建设预留弹性。该区域主要为湿地及农田生态系统, 具有较高的生态系统服务价值, 但由于属发展备用地, 未来优先保护区面积可能进一步减少, 区域生物多样性受到威胁。
4.3 沿海湾区城市生态空间保护规划对比中国海岸带由北向南跨越了40个纬度, 自东向西跨越了20个经度, 包括了粤港澳大湾区、北部湾区、环渤海湾区、环杭州湾四大湾区。国内外学者已经在不同区域尺度上开展了生态空间规划及生物多样性保护优先区域识别工作, 其中深圳市土地利用总体规划(2006-2020)将大部分海岸带区域划为限制建设区和禁止建设区, 上海市生态空间专项规划(2018-2035)将长江口大部分沿海区域视为一类、二类生态空间, 在各湾区城市中海岸带区域都具有优先保护性。对比厦门市目前的土地利用规划(2006-2020), 海岸带区域保护力度较弱, 适宜植物生长的理论优先保护区均未受到保护, 在未来的生物多样性保护规划中应予以重视。
5 结论生物多样性保护优先区识别是目前国内外研究的热点问题, 但目前针对海湾型城市植物生物多样性保护优先区识别的研究较少。本文识别厦门市重点保护植物的理论空间优先保护格局, 探究优先保护区的生态空间保护利用现状。研究结果表明本地重点保护植物分布受海拔影响最大, 大部分物种的适宜分布生境集中于海岸带地区。海岸带地区的理论优先保护区被建设用地大量占用, 少部分优先保护区虽未占用, 但生态保护不足, 存在较大的生态空间保护空缺, 而西部山地地区的优先保护区得到较好的保护。为避免优先保护区面积进一步减少, 在未来的土地利用及生物多样性保护规划中, 需重点关注海岸带区域优先保护区的生态保护, 提升海岸带栖息地生境质量, 将目前优先保护区内的发展备用地改划为生态留白空间。对一级和二级优先保护区分别实施刚性和弹性的生态保育措施, 在保护生物多样性的同时, 严控对海岸带区域优先保护区的进一步开发利用, 协调优先保护区内保护与开发利用间的关系, 提升城市的生态安全。
| [1] |
中华人民共和国生态环境部. 2019中国生态环境状况公报. (2020-06-02)[2021-04-21\]. http://www.mee.gov.cn/hjzl/sthjzk/zghjzkgb/202006/P020200602509464172096.pdf.
|
| [2] |
Xu X, Jiang B, Chen M, Bai Y, Yang G. Strengthening the effectiveness of nature reserves in representing ecosystem services: The Yangtze River Economic Belt in China. Land Use Policy, 2020, 96: 104717. DOI:10.1016/j.landusepol.2020.104717 |
| [3] |
杨朝辉. 基于C-Plan技术的贵州生物多样性优先保护规划研究. 贵州大学, 2020. |
| [4] |
孙思琦. 自然保护区对植物保护效果分析. 中央民族大学, 2020. |
| [5] |
Moilanen A, Wilson K, Possingham H. Spatial conservation prioritization: quantitative methods and computational tools. Oxford University Press, 2009, 349. |
| [6] |
Kukkala A S, Moilanen A. Ecosystem services and connectivity in spatial conservation prioritization. Landscape Ecology, 2017, 32(1): 5-14. DOI:10.1007/s10980-016-0446-y |
| [7] |
Delavenne J, Metcalfe K, Smith R J, Vaz S, Martin C S, Dupuis L, Coppin F, Carpentier A. Systematic conservation planning in the eastern English Channel: comparing the Marxan and Zonation decision-support tools. ICES Journal of Marine Science, 2012, 69(1): 75-83. DOI:10.1093/icesjms/fsr180 |
| [8] |
Lu M, Qu Y. Biodiversity conservation development based on Systematic Conservation Planning in urban areas. Energy Procedia, 2018, 153: 484-488. DOI:10.1016/j.egypro.2018.10.027 |
| [9] |
Moilanen A. Landscape zonation, benefit functions and target-based planning: unifying reserve selection strategies. Biological Conservation, 2007, 134(4): 571-579. DOI:10.1016/j.biocon.2006.09.008 |
| [10] |
Moilanen A, Anderson B J, Eigenbrod F, Heinemeyer A, Roy D B, Gillings S, Armsworth P R, Gaston K J, Thomas C D. Balancing alternative land uses in conservation prioritization. Ecological Applications, 2011, 21(5): 1419-1426. DOI:10.1890/10-1865.1 |
| [11] |
Lehtomäki J, Moilanen A. Methods and workflow for spatial conservation prioritization using Zonation. Environmental Modelling & Software, 2013, 47: 128-137. |
| [12] |
段义忠, 王海涛, 王驰, 杜忠毓. 气候变化下濒危植物半日花在中国的潜在分布. 植物资源与环境学报, 2020, 29(2): 55-68. DOI:10.3969/j.issn.1674-7895.2020.02.07 |
| [13] |
段义忠, 王驰, 王海涛, 杜忠毓, 贺一鸣, 柴乖强. 不同气候条件下沙冬青属植物在我国的潜在分布——基于生态位模型预测. 生态学报, 2020, 40(21): 7668-7680. |
| [14] |
段义忠, 鱼慧, 王海涛, 杜忠毓. 孑遗濒危植物四合木(Tetraena mongolica)的地理分布与潜在适生区预测. 植物科学学报, 2019, 37(3): 337-347. |
| [15] |
Tsoar A, Allouche O, Steinitz O, Rotem D, Kadmon R. A comparative evaluation of presence-only methods for modelling species distribution. Diversity and distributions, 2007, 13(4): 397-405. DOI:10.1111/j.1472-4642.2007.00346.x |
| [16] |
胡文佳, 晁碧霄, 王玉玉, 董鹏, 张典, 俞炜炜, 陈光程, 陈彬. 基于最大熵模型的福建省红树林潜在适生区评估. 中国环境科学, 2020, 40(9): 4029-4038. DOI:10.3969/j.issn.1000-6923.2020.09.035 |
| [17] |
崔绍朋, 王泽鑫, 吉晟男. 基于生态位模型预测山西省双条杉天牛的适生区. 农业与技术, 2020, 40(17): 71-73. |
| [18] |
Phillips S J, Dudík M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161-175. DOI:10.1111/j.0906-7590.2008.5203.x |
| [19] |
Zhong Y, Xue Z, Jiang M, Liu B, Wang G. The application of species distribution modeling in wetland restoration: A case study in the Songnen Plain, Northeast China. Ecological Indicators, 2021, 121: 107137. DOI:10.1016/j.ecolind.2020.107137 |
| [20] |
Barik S, Saha G K, Mazumdar S. Potentially suitable habitat, connectivity and priority conservation areas for White-breasted waterhen (Amaurornis phoenicurus) and Bronze-winged jacana (Metopidius indicus). Wetlands, 2021, 41(3): 1-15. DOI:10.1007/s13157-021-01433-6 |
| [21] |
Kaky E, Gilbert F. Allowing for human socioeconomic impacts in the conservation of plants under climate change. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology, 2020, 154(3): 295-305. DOI:10.1080/11263504.2019.1610109 |
| [22] |
VanCompernolle M, Knouft J H, Ficklin D L. Multispecies conservation of freshwater fish assemblages in response to climate change in the southeastern United States. Diversity and Distributions, 2019, 25(9): 1388-1398. DOI:10.1111/ddi.12948 |
| [23] |
Patrizzi N S, Dobrovolski R. Integrating climate change and human impacts into marine spatial planning: A case study of threatened starfish species in Brazil. Ocean & Coastal Management, 2018, 161: 177-188. |
| [24] |
Freire Filho R, Palmeirim P. Potential distribution of and priority conservation areas for the Endangered Caatinga howler monkey Alouatta ululata in north-eastern Brazil. Oryx, 2019, 1-9. |
| [25] |
厦门市人民政府. 厦门概况. [2021-04-30]. http://www.xm.gov.cn/zjxm/xmgk/202011/t20201125_2498295.htm
|
| [26] |
高美蓉. 基于多源遥感数据的厦门城市森林景观格局研究. 中国林业科学研究院, 2013. |
| [27] |
赵娟娟, 宋晨晨, 刘时彦. 城市植物种类构成的特征分析——以厦门市为例. 西南大学学报(自然科学版), 2018, 40(7): 1-8. |
| [28] |
李渊, 林锋, 严泽幸. 基于Landsat遥感影像的城市化演变分析——以厦门市为例. 城市建筑, 2019, 16(25): 138-142. DOI:10.3969/j.issn.1673-0232.2019.25.030 |
| [29] |
刘亚军, 郑秋燕, 郭玉清, 汲珊珊, 陈建春, 李军, 刘爱原. 厦门滨海湿地红树林资源与互花米草入侵分布. 集美大学学报(自然科学版), 2020, 25(2): 88-96. |
| [30] |
Yi Y, Zhou Y, Cai Y, Yang W, Li Z, Zhao X. The influence of climate change on an endangered riparian plant species: The root of riparian Homonoia. Ecological Indicators, 2018, 92: 40-50. DOI:10.1016/j.ecolind.2017.05.004 |
| [31] |
马东旭, 周一, 陆双飞, 殷晓洁, 周斯怡. 基于MaxEnt模型的11种云南常见阔叶树种气候适宜性研究. 西南林业大学学报(自然科学), 2020, 40(5): 64-72. |
| [32] |
Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions. Ecological modelling, 2006, 190(3-4): 231-259. DOI:10.1016/j.ecolmodel.2005.03.026 |
| [33] |
Loiselle B A, Howell C A, Graham C H, Goerck J M, Brooks T, Smith K G, Williams P H. Avoiding pitfalls of using species distribution models in conservation planning. Conservation biology, 2003, 17(6): 1591-1600. DOI:10.1111/j.1523-1739.2003.00233.x |
| [34] |
van Proosdij A S J, Sosef M S M, Wieringa J J, Raes N. Minimum required number of specimen records to develop accurate species distribution models. Ecography, 2016, 39(6): 542-552. DOI:10.1111/ecog.01509 |
| [35] |
Pearson R G, Raxworthy C J, Nakamura M, Townsend Peterson, A. ORIGINAL ARTICLE: Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. Journal of biogeography, 2007, 34(1): 102-117. |
| [36] |
麻亚鸿. 基于最大熵模型(MaxEnt)和地理信息系统(ArcGis)预测藓类植物的地理分布范围. 上海师范大学, 2013. |
| [37] |
Wang G, Wang C, Guo Z, Dai L, Wu Y, Liu H, Li Y, Chen H, Zhang Y, Zhao Y, Cheng H, Ma T, Xue F. Integrating Maxent model and landscape ecology theory for studying spatiotemporal dynamics of habitat: Suggestions for conservation of endangered Red-crowned crane. Ecological Indicators, 2020, 116: 106472. DOI:10.1016/j.ecolind.2020.106472 |
| [38] |
Pearson R G, Raxworthy C J, Nakamura M, Townsend Peterson A. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. Journal of biogeography, 2007, 34(1): 102-117. |
| [39] |
刘振生, 高惠, 滕丽微, 苏云, 王晓勤, 孔芳毅. 基于MAXENT模型的贺兰山岩羊生境适宜性评价. 生态学报, 2013, 33(22): 7243-7249. |
| [40] |
Wakie T T, Evangelista P H, Jarnevich C S, Laituri M. Mapping current and potential distribution of non-native Prosopis juliflora in the Afar region of Ethiopia. PloS one, 2014, 9(11): e112854. DOI:10.1371/journal.pone.0112854 |
| [41] |
Dos Santos L A, Mendes M F, Krüger A P, Blauth M L, Gottschalk M S, Garcia F R M. Global potential distribution of Drosophila suzukii (Diptera, Drosophilidae). Plos one, 2017, 12(3): e0174318. DOI:10.1371/journal.pone.0174318 |
| [42] |
Shadman M, Meshgi B, Fathi S, Hanafi-Bojd A A, Majidi-Rad M, Modabbernia G. Mapping habitat suitability for gastrointestinal nematodiasis of ruminants in southern Caspian Sea littoral: a predicted risk pattern model based on the MaxEnt. Tropical Animal Health and Production, 2020, 52(6): 3843-3854. DOI:10.1007/s11250-020-02423-2 |
| [43] |
李丽鹤, 刘会玉, 林振山, 贾俊鹤, 刘翔. 基于MAXENT和ZONATION的加拿大一枝黄花入侵重点监控区确定. 生态学报, 2017, 37(9): 3124-3132. |
| [44] |
Hernandez P A, Graham C H, Master L L, Albert D L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography, 2006, 29(5): 773-785. DOI:10.1111/j.0906-7590.2006.04700.x |
| [45] |
Swets J A. Measuring the accuracy of diagnostic systems. Science, 1988, 240(4857): 1285-1293. DOI:10.1126/science.3287615 |
| [46] |
Jalkanen J, Vierikko K, Moilanen A. Spatial prioritization for urban Biodiversity Quality using biotope maps and expert opinion. Urban Forestry & Urban Greening, 2020, 49: 126586. |
| [47] |
Leathwick J, Moilanen A, Francis M, Elith J, Taylor P, Julian K, Hastie T, Duffy C. Novel methods for the design and evaluation of marine protected areas in offshore waters. Conservation Letters, 2008, 1(2): 91-102. DOI:10.1111/j.1755-263X.2008.00012.x |
| [48] |
肖静, 崔莉, 李俊清. 基于ZONATION的岷山山系多物种保护规划. 生态学报, 2016, 36(2): 420-429. |
| [49] |
王春晶. 气候变化背景下中国野生植物地理分布格局及优先保护. 北京林业大学, 2017. |
| [50] |
洪滔, 吴承祯, 陈灿, 谢安强, 李键, 林勇明, 范海兰, 林晗. 闽东南海岸带山地几种树种海拔资源位的叶绿素特征. 福建农林大学学报(自然科学版), 2013, 42(1): 35-39. DOI:10.3969/j.issn.1671-5470.2013.01.007 |