 2021, Vol. 41
2021, Vol. 41文章信息
- 张作合, 张忠学
- ZHANG Zuohe, ZHANG Zhongxue
- 寒地稻田不同灌溉模式下稻株生长与水分消耗利用
- Rice plant growth and water consumption and utilization under different irrigation modes in cold paddy field
- 生态学报. 2021, 41(11): 4586-4595
- Acta Ecologica Sinica. 2021, 41(11): 4586-4595
- http://dx.doi.org/10.5846/stxb201906221322
-
文章历史
- 收稿日期: 2019-06-22
- 网络出版日期: 2021-04-06

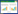




2. 绥化学院农业与水利工程学院, 绥化 152061;
3. 农业农村部农业水资源高效利用重点实验室, 哈尔滨 150030
2. College of Agriculture and Hydraulic Engineering, Suihua University, Suihua 152061, China;
3. Key Laboratory of Effective Utilization of Agricultural Water Resources, Ministry of Agriculture and Rural Affairs, Harbin 150030, China
水稻是我国主要粮食作物, 播种面积大, 约占粮食作物总种植面积的26.7%[1]。稻田生态系统作为重要的农田生态系统之一, 也是半自然的人工湿地生态系统[2]。稻田生态系统是为了满足人类社会生存和发展的需要, 采取积极科学合理的人为干预, 利用稻田生物与非生物环境之间及稻田生物种群之间的关系进行人类所需食物的生产, 具有社会、经济、生态功能和自然、社会双重属性的复合生态系统[3]。水分转化循环是稻田生态系统一个重要过程。作物生长发育过程中吸收的水分, 直接参与光合作用的不足1%, 而99%以上的水分通过作物蒸腾和土壤蒸发而消耗(以蒸腾为主)。水分循环增加了稻田系统环境的湿度、降低了系统环境的气温、改善田间的小气候, 维持了作物的生长和农产品的生产。同时, 水分循环也带动养分循环, 增加水稻对养分的吸收利用。因此, 研究稻田生态系统的水分消耗利用非常重要。
稻田生态系统是一个高度开放的农田生态系统, 其物质循环和能量转换的强度显著高于其他农田生态系统。由于稻田生态系统具有区别于其他农田生态系统的水分消耗过程, 在农田生态系统的研究中受到极大关注[2]。有研究[4]表明, 滴灌旱直播与漫灌旱直播处理的叶、鞘和茎的湿基含水率在全生育期均低于常规插秧淹灌处理, 而根部湿基含水率从分蘖后期开始均高于常规插秧淹灌处理。魏永霞等[5]研究寒地黑土区水稻耗水特性及其对水分利用效率的影响, 发现拔节孕穗期各处理耗水量均值最高为77.25 mm, 同时耗水模系数均值也达到最高为23.09%。进一步研究黑土区水稻生长的耗水规律, 发现水稻生育期内耗水强度呈现先上升后下降的趋势, 在抽穗开花期达到最大值, 为5.66 mm/d;而耗水模系数在分蘖期达到最大, 均值为43.98%[6]。以上研究对稻田生态系统的水分消耗全过程研究较为模糊, 而较为系统完整地阐述分析稻田生态系统的水分消耗规律的研究较少。
本文以常规淹灌作为对比, 采用蒸渗仪与田间小区结合的方法, 在时间尺度上, 分析了三种灌溉模式的湿基含水率、干物质积累、耗水强度及组成和阶段生物量水分利用效率的变化规律, 系统地研究了不同灌溉模式下水稻植株生长与水分消耗利用, 以期为寒地黑土区稻田生态系统水分消耗利用研究提供理论依据和技术支撑。
1 材料与方法 1.1 研究区概况试验于2018年在黑龙江省庆安县和平灌区水稻灌溉试验站(127°40′45″E, 46°57′28″N)进行。试验站位于呼兰河流域中上游, 属于寒地黑土区。寒温带大陆性季风气候, 属第三积温带, 平均日照时数为2600 h, 多年平均降水量500—600 mm, 平均气温为2—3 ℃, 全年无霜期约128 d, 是黑龙江省典型的水稻生产区。供试土壤为黑土, 其基本土壤理化性质:pH值6.40, 有机质41.80 g/kg, 全氮1.51 g/kg, 全磷15.61 g/kg, 全钾19.86 g/kg, 碱解氮148.27 mg/kg, 速效磷24.22 mg/kg, 速效钾156.13 mg/kg。供试水稻品种为绥粳18(Oryza sativa subsp. Keng), 在研究区已大面积推广种植。2018年水稻生育期内试验区最高温度、最低温度和降雨量变化如图 1所示。

|
| 图 1 水稻生育期最高温度、最低温度和降雨量变化情况 Fig. 1 Changes of maximum temperature, minimum temperature and rainfall during the growth period of rice |
本试验设置控制灌溉(C)、间歇灌溉(I)和常规淹灌(F)三种水分管理模式(见表 1)。水稻控制灌溉模式各处理根据控制指标来确定灌水时间和定额, 当田面无水层后, 每天8:00和18:00, 用土壤水分测定仪(TPIME-PICO64/32型)分别测定各小区的土壤含水率, 当土壤含水率接近或低于灌水下限时, 灌水至上限, 并记录各处理的灌水量;间歇灌溉和常规淹灌模式各处理的试验小区在田面无水层时, 由人工灌水至上限。共计3个处理, 每个处理3次重复, 共计9个试验小区, 方形小区面积100 m2采用随机区组排列, 同时对应处理设置蒸渗仪测坑, 蒸渗仪面积1m2, 同一处理的试验小区和蒸渗仪田间管理方式相同。当地施氮肥水平110 kg/hm2, 按基肥∶蘖肥∶促花肥∶保花肥比例为4.5∶2∶1.5∶2分施;K2O施入量为80 kg/hm2, 基肥∶穗肥按1∶1分施;P2O5施入量为45 kg/hm2, 作为基肥一次性施入。供试肥料选择尿素(含N 46.4%)、过磷酸钙(含P2O5 12%)、硫酸钾(含K2O 52%), 并换算成实际化肥施入量。2018年5月19日移栽水稻幼苗, 插秧密度为25穴/m2, 每穴3株, 9月21日收获。水稻生长期注意田间管理, 严格控制病虫草害。
| 灌溉方式 Irrigation mode |
返青期 Rejuvenation period |
分蘖初 Early tillering |
分蘖盛 Tillering period |
分蘖末 Late tillering |
拔节孕穗 Jointing booting |
抽穗开花 Heading and flowering |
乳熟 Milk ripening |
黄熟 Yellow ripening |
| 控制灌溉Control irrigation | 0—30 | 0.7θs—0 | 0.7θs—0 | 晒田 | 0.8θs—0 | 0.8θs—0 | 0.7θs—0 | 落干 |
| 间歇灌溉Intermittent irrigation | 0—30 | 0—40 | 0—40 | 晒田 | 0—30 | 0—40 | 0—40 | 落干 |
| 常规淹灌Flooding irrigation | 0—30 | 0—50 | 0—50 | 晒田 | 0—50 | 0—50 | 0—50 | 落干 |
| θs为根层土壤饱和含水率质量分数。“—”前数据为水分控制下限,“—”后数据位水分控制上限 | ||||||||
水稻各器官干物质积累量和含水率的测定:在分蘖期(T)、拔节孕穗期(JB)、抽穗开花期(HF)、乳熟期(M)和黄熟期(R)从每个小区和蒸渗仪中各随机选取6穴植株, 整株取样时, 挖取40 cm带根土块, 放入尼龙筛网袋(40目)里, 浸泡半个小时后, 用清水冲洗干净, 将植株的叶、茎鞘、穗和根分离后, 分别称量各器官鲜重并记录, 之后将各器官装入信封放入烘箱105 ℃杀青30 min, 然后调至80 ℃烘干后, 用精度0.01 g的电子天平称取植株各器官干质量。
作物耗水量及耗水强度:每天08:00使用称量式有底蒸渗仪获取总体质量(收集称量渗漏水量), 根据前后的质量差(M1)计算水稻植株每天实际耗水量, 试验在自动感应式遮雨棚内进行, 因而不考虑生长季内降雨量和地下水补给量的影响。在各个生育时期的植株干物质进行取样时, 记录取样前、后蒸渗仪的总体质量, 两次质量差值(M2)即为取样引起蒸渗仪的质量变化, 各生育时期取样当天的耗水量为M1-M2的差值。
气象数据由试验站DZZ2型自动气象站(天津气象仪器厂)自动记录。
1.4 计算方法和数据分析水稻植株各器官含水率计算公式:
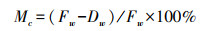
|
式中:Mc为植株各器官湿基含水率, %;Fw为植株各器官鲜质量, g;Dw为植株各器官干质量, g。
作物阶段耗水量计算公式:
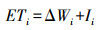
|
式中:ETi为生育期阶段耗水量, mm;i为生育阶段, i为1、2、3、4、5分别代表分蘖期, 拔节孕穗期、抽穗开花期、乳熟期和黄熟期;ΔWi为生育期阶段储水量差值, mm;Ii为生育期阶段灌水量, mm。
作物耗水强度计算公式:
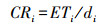
|
式中:CRi为生育期阶段耗水强度, mm/d;di为生育期阶段持续时间, d;
作物耗水模系数计算公式:
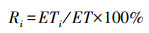
|
式中:Ri为生育期阶段耗水模系数, %;ET为全生育期耗水总量, mm。
阶段生物量水分利用效率:
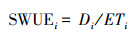
|
式中:Di为各生育期阶干物质增加量, 生育阶段, i为1、2、3、4、5分别代表分蘖期, 拔节孕穗期、抽穗开花期、乳熟期和黄熟期。
2 结果与分析 2.1 不同灌溉模式对水稻各器官水分分布的影响随着水稻生长发育植株体内的水分会发生变化, 并对稻田生态环境作出响应。分析不同灌溉模式下水稻植株体内水分的分布和变化规律, 对研究稻田生态系统水分消耗利用具有重要意义。由图 2可知(图中同组不同小写字母表示在0.05水平上差异显著, 下同), 不同灌溉模式水稻植株各器官湿基含水率随生育期的推进均大致呈下降趋势。三种灌溉模式下水稻叶片的湿基含水率由分蘖期的68.91%、70.38%和71.34%降至黄熟期的57.34%、61.71%和63.57%, 各生育期的叶片湿基含水率均以常规淹灌模式最大, 控制灌溉模式最小, 除拔节孕穗期外, 控制灌溉模式和间歇灌溉模式与常规淹灌模式之间的差异显著(P < 0.05), 分别降低1.51%—9.80%和0.48%—2.93%;各种灌溉模式下水稻茎鞘的湿基含水率与叶片的湿基含水率变化趋势基本一致, 均以常规淹灌模式最大, 控制灌溉模式最小, 除分蘖期外, 控制灌溉模式和间歇灌溉模式与常规淹灌模式之间的差异显著(P < 0.05), 分别降低2.36%—11.11%和0.64%—4.57%。水稻穗部的湿基含水率由拔节孕穗期的88.64%、72.67%和66.45%降至黄熟期的28.03%、25.27%和24.32%, 在拔节孕穗期和抽穗开花期, 控制灌溉模式和间歇灌溉模式的穗部湿基含水率与常规淹灌模式之间的差异显著(P < 0.05), 分别降低21.63%—33.39%和9.36%—12.33%, 而在乳熟期和黄熟期, 控制灌溉模式和间歇灌溉模式的穗部湿基含水率与常规淹灌模式之间的差异不显著(P>0.05);间歇灌溉模式和常规淹灌模式各生育期根的湿基含水率均呈逐渐减小的趋势, 控制灌溉模式呈现出先增大后减小的趋势。从分蘖期到拔节孕穗期, 间歇灌溉模式和常规淹灌模式根的湿基含水率与控制灌溉模式变化趋势相反, 这可能是由于间歇灌溉和常规淹灌模式水稻长期处于有水层状态, 分蘖末期晒田使水稻根部活力减小, 吸水能力能下降, 导致根部湿基含水率较控制灌溉模式显著下降, 对于控制灌溉模式来说, 水稻长期处于无水层状态, 因而分蘖末期的水分亏缺对控制灌溉水稻的根部湿基含水率影响很小。在抽穗开花期、乳熟期和黄熟期, 间歇灌溉模式和常规淹灌模式水稻的根部湿基含水率显著低于控制灌溉模式(P < 0.05), 其中乳熟期和黄熟期分别降低1.48%、2.11%和2.21%、3.98%, 表明控制灌溉模式能够有效地延缓水稻生育后期的根系衰老。

|
| 图 2 不同处理水稻各器官湿基含水率变化曲线 Fig. 2 Variation curves of wet base water content of rice organs under different treatments 图中不同小写字母表示数据在P<0.05水平差异显著 |
水稻植株耗水过程和各器官湿基含水率都与干物质积累过程有直接关系[6], 因此分析不同灌溉模式水稻各器官干物质变化规律对研究稻田生态系统十分重要。由图 3可知, 不同灌溉模式水稻各器官干物质在整个生育期的变化规律大致相同。各处理水稻植株叶片干物质积累量随着生育的推进均呈单峰曲线变化规律, 同时叶片的干物质积累量最大值出现在拔节孕穗期。各处理水稻植株茎鞘干物质积累量在抽穗开花期达到最大, 这样有利于保障后期光合产物向籽粒转移。通常情况下, 水稻植株叶片和茎鞘的干物质积累量达到最大值之后, 开始向穗部转移, 保证穗部干物质积累量在后期迅速增加。各处理水稻植株根部干物质积累量的变化规律与叶片和茎鞘大致相同, 均呈倒“Ⅴ”型曲线, 同时最大值出现在抽穗开花期。

|
| 图 3 不同生育期水稻各器官干物质积累动态曲线 Fig. 3 Dynamic curves of dry matter accumulation of rice various organs at different growth stages |
不同灌溉模式在各生育期的控水标准不同, 从而导致水稻植株各器官干物质积累过程出现差异。除抽穗开花期外, 三种灌溉模式之间的水稻植株叶片干物质积累量差异性均显著(P < 0.05)。在分蘖期, 叶片干物质积累量大小关系为常规淹灌模式>间歇灌溉模式>控制灌溉模式;在拔节孕穗期, 叶片干物质积累量大小关系为常规淹灌模式>控制灌溉模式>间歇灌溉模式;在抽穗开花期和乳熟期, 叶片干物质积累量大小关系为控制灌溉模式>间歇灌溉模式>常规淹灌模式;在黄熟期, 叶片干物质积累量大小关系为间歇灌溉模式>控制灌溉模式>常规淹灌模式;从抽穗开花期到黄熟期由叶片向穗转运的干物质量分别为2.67 g/穴、1.90 g/穴、2.40 g/穴。不同灌溉模式之间的水稻植株茎鞘干物质积累量差异性均显著(P < 0.05)。在分蘖期和拔节孕穗期, 茎鞘干物质积累量大小关系为常规淹灌模式>间歇灌溉模式>控制灌溉模式;在抽穗开花期、乳熟期和黄熟期, 茎鞘干物质积累量大小关系为控制灌溉模式>间歇灌溉模式>常规淹灌模式;从抽穗开花期到黄熟期由茎鞘向穗转运的干物质量分别为4.68 g/穴、4.53 g/穴、4.08 g/穴。总的来看, 控制灌溉模式下叶片和茎鞘的后期生长补偿效应显著。水稻植株穗部干物质积累量随着生育的推进逐渐增加, 在乳熟期和黄熟期, 不同灌溉模式之间的水稻植株穗部干物质积累量差异性均显著(P < 0.05), 大小关系为控制灌溉模式>间歇灌溉模式>常规淹灌模式。从拔节孕穗期到黄熟期, 三种灌溉模式的水稻植株根部干物质积累量大小关系为控制灌溉模式>间歇灌溉模式>常规淹灌模式, 且各处理之间差异性均显著(P < 0.05)。
2.3 不同灌溉模式对水稻阶段耗水特征的影响水稻耗水量是土壤与大气界面、植株与大气界面和土壤界面的系统水分转化的结果, 它对研究稻田生态系统和水稻生产具有重要的现实意义。由图 4可知, 不同灌溉模式之间的水稻各阶段耗水量差异性均显著(P < 0.05), 大小关系均为常规淹灌模式>间歇灌溉模式>控制灌溉模式, 各处理分蘖期的耗水量均值最大(172.19 mm), 黄熟期的耗水量均值最小(64.29 mm)。在分蘖期, 控制灌溉模式水稻耗水量较常规淹灌模式降幅最大, 达49.27%;而间歇灌溉模式水稻耗水量较常规淹灌模式降低20.59%。在分蘖期, 各灌溉模式之间的水稻各阶段耗水模系数差异性显著(P < 0.05), 大小关系为常规淹灌模式>间歇灌溉模式>控制灌溉模式, 由于分蘖期是水稻营养生长的重要阶段, 持续时间较长(一般为45 d左右), 各处理的耗水模系数均为最大值。从拔节孕穗期到黄熟期, 水稻的耗水模系数大小关系为控制灌溉模式>间歇灌溉模式>常规淹灌模式。拔节孕穗期各灌溉模式耗水模系数大小仅低于分蘖期, 主要是由于这个时期为颖花与枝梗发育的重要时期, 充分的水分能够为水稻孕穗及出穗提供良好的生长环境。随着水稻生育期的推进, 逐步进入籽粒灌浆以及产量形成的关键阶段, 在乳熟期各灌溉模式耗水模系数仍然较高。

|
| 图 4 不同处理水稻阶段耗水量及耗水模系数 Fig. 4 Water consumption and water consumption percentage of rice at different stages under different treatments |
图 5为三种灌溉模式下水稻各生育时期的耗水强度及组成。从全生育期来看, 水稻各处理生育期的叶面蒸腾强度、作物需水强度及田间耗水强度均呈现先上升后下降的趋势。水稻分蘖期历时较长, 且处于营养生长阶段, 后期田间耗水由棵间蒸发为主转向植株蒸腾为主。由于气温持续升高的和生育期的推进, 抽穗开花期耗水强度增至5.64 mm/d、6.59 mm/d和7.71 mm/d, 这个时期是水稻生殖生长和营养生长并进阶段, 水稻植株生长速度加快且冠层覆盖率较高, 使水稻的需水量急剧增大。各灌溉模式水稻进入乳熟期后耗水强度迅速降低, 至黄熟期耗水强度降低为2.66 mm/d、3.19 mm/d和3.80 mm/d。通过分析水稻田间耗水强度的组成要素与田间耗水强度的关系发现控制灌溉模式下, 棵间蒸发强度、叶面蒸腾强度、渗漏强度和作物需水量分别占田间耗水强度的16.31%—48.62%、33.20%—60.46%、18.18%—27.44%和72.56%—81.82%;间歇灌溉模式下, 棵间蒸发强度、叶面蒸腾强度、渗漏强度和作物需水量分别占田间耗水强度的17.75%—44.70%、27.27%—62.37%、19.88%—33.23%和66.77%—80.12%;常规淹灌模式下, 棵间蒸发强度、叶面蒸腾强度、渗漏强度和作物需水量分别占田间耗水强度的19.46%—49.70%、26.85%—59.40%、20.46%—29.47%和70.53%—79.54%。

|
| 图 5 不同处理水稻阶段耗水强度 Fig. 5 Water consumption Intensity of rice at different stages under different treatments |
在时间尺度上, SWUEi反映了不同阶段单位耗水量提高水稻植株干物质积累量的能力。由图 6可知, 控制灌溉模式和间歇灌溉模式在整个生育期SWUEi的曲线出现多峰, 其中分蘖期、抽穗开花期和黄熟期出现三个峰值, 而常规淹灌模式在整个生育期SWUEi曲线先下降后上升, 且不同灌溉模式之间的水稻SWUEi差异性均显著(P < 0.05)。在水稻分蘖期, 控制灌溉模式和间歇灌溉模式的SWUEi较常规淹灌模式分别提高了73.71%、15.82%;在水稻拔节孕穗期, 与常规淹灌模式相比, 控制灌溉模式和间歇灌溉模式的SWUEi分别提高了62.80%、24.31%;在水稻拔抽穗开花期, 控制灌溉模式和间歇灌溉模式的SWUEi较常规淹灌模式分别提高了144.35%、79.70%;在水稻乳熟期, 与常规淹灌模式相比, 控制灌溉模式和间歇灌溉模式的SWUEi分别提高了257.10%、90.94%;在水稻黄熟期, 控制灌溉模式和间歇灌溉模式的SWUEi较常规淹灌模式分别提高了39.84%、22.77%。研究结果表明, 灌溉模式对水稻的SWUEi影响显著(P < 0.05), 其中控制灌溉模式能够有效地提高水稻各生育期的SWUEi值, 改变稻田生态系统的阶段生物量水分利用效率。

|
| 图 6 不同处理水稻阶段生物量水分利用效率 Fig. 6 Biomass water use efficiency of rice for different treatments at different stages |
稻株含水量对作物的形态结构及生理生化过程具有重要影响, 植株体内的水分状况会直接影响作物的生长发育和产量。当然, 稻株的含水量受制于稻田生态系统的水分状态。所以在不同灌溉模式下, 随着稻田生态系统环境的变化, 水分在作物体内各器官会重新分配, 这是水稻对外界条件作出的调整和适应[7]。分析本试验中叶片湿基含水率下降的根本原因可以发现, 在抽穗开花期之前可能是由于随着叶片的生长光合速率增加[4], 气孔开度增大, 蒸腾水分散失增加, 导致湿基含水率的下降[8]。而抽穗开花期之后主要是由于叶片衰老引起的, 叶片衰老时营养物质被转运至籽粒, 这个转运过程导致叶片的湿基含水率不断下降[9]。控制灌溉模式后期的叶片湿基含水率下降速率显著高于常规淹灌模式的原因主要是由于水稻长期在水分胁迫下叶片生长受抑制, 后期衰老的较快[10]。作为水稻吸收水分的重要器官, 分析根系的湿基含水率的变化规律十分重要。本试验研究结果表明, 常规淹灌模式和间歇灌溉模式水稻根系长期处于淹水状态, 导致后期根系活力下降, 光合速率和蒸腾速率下降, 同时营养物质的吸收和运输受到影响, 而控制灌溉模式水稻的生育后期湿基含水率增大, 提高了根系活力和吸水能力[11-12]。
光合作用是稻田生态系统组成的一个重要环节, 光合产物在不同器官中积累与分配构成水稻干物质。本试验研究结果表明:分蘖期耗水量增加能够促进叶片的快速发育, 而拔节孕穗期叶片生长发育是基于分蘖后期叶片良好的发育。在抽穗开花期保证茎鞘的干质量积累, 有利于后期营养物质向籽粒的转运。适当的水分调控改善土壤的通气性, 加速有机质的分解, 从而提高土壤肥力、促进根系的发育[13], 然而太过于追求水稻植株根部干物质的积累会抑制叶片和茎鞘的干物质向穗部转移, 对籽粒的干物质积累产生负效应, 这与吴岳轩等[14]的研究结果一致。控制灌溉模式下, 水稻分蘖期水分亏缺有利于营养物质优先供给根系, 促进根系的生长和干物质的积累[15], 可以促进根系对水分及营养的吸收, 提高根系活力, 有利于延缓叶片衰老, 激发作物生产潜能[16]。
分析作物耗水过程对研究水稻自身水分调节能力及适应干旱环境的能力具有重要作用[17-18]。本研究发现, 水稻生育期的阶段耗水量呈逐渐降低的趋势, 其中分蘖期耗水量占全生育期耗水量的26%—33%, 这可能是由于分蘖期的历时较长, 叶片的覆盖率较低, 棵间蒸发强度增大所致, 而棵间蒸发与表层土壤含水率的高低密切相关。本试验中水稻的耗水强度在抽穗开花期达到最大, 但也有学者研究发现水稻在乳熟期的耗水强度最大[19], 这可能是由于不同品种、气候特征、土壤类型和田间管理方式所引起的。水稻是高耗水作物之一, 研究其耗水量规律对提高水分利用效率和水资源优化配置具有重大意义[20]。当水稻植株的生理活动减弱时, 叶片气孔导度和蒸腾速率均会明显降低[21]。而叶片在受到高温和辐射时气孔会主动降低或关闭, 高温过后叶片气孔导度不能很快地恢复初始状态, 而是存在一定的滞后性[22]。控制灌溉模式产生的水分胁迫使得水稻植物通过关闭部分气孔来调节自身的耗水量, 从而导致气象因子对植株耗水量的影响减弱[5], 植株通过水分的自我调解来适应稻田生态系统变化。
阶段生物量水分利用效率是评价作物生长适宜程度的综合生理生态指标[23], 它能够反映作物各阶段耗水量与干物质生产之间的关系。本试验研究结果表明, 水稻的SWUEi分别在分蘖期和、抽穗开花期和成熟期达到高峰, 表明植株在这三阶段对水分利用能力较强, 其中抽穗开花期是水稻植株是对水分最敏感阶段, 也是生殖生长的关键时期, 这个时期水稻光合作用及新陈代谢最旺盛, 适宜的水分环境有利于水稻植株茎鞘和穗部生长, 促进抽穗扬花, 也是成熟期获得高产的必要条件。这与张亚琦等[24]研究结果存在一定的差异, 可能是由于作物种类、土壤类型及气候特征不同所引起的。综合来看, 稻田生态系统水分消耗利用过程较为复杂需要长期的试验研究。
针对东北寒地黑土区稻田生态系统的特点, 本试验采用蒸渗仪与田间小区结合的方法, 研究了不同灌溉模式下水稻植株生长与水分消耗利用, 但考虑多方面因素, 稻田生态系统水分消耗利用机理还有待进一步验证, 并根据各灌溉模式结合实际降雨量, 来调控稻田灌溉水量, 能够有效地维持SPAC稻田生态系统平衡, 保证农业和生态系统的可持续发展。
4 结论(1) 不同灌溉模式水稻植株各器官湿基含水率随生育期的推进均大致呈下降趋势。与间歇灌溉模式和常规淹灌模式相比, 控制灌溉模式能够有效地延缓水稻生育后期的根系衰老。
(2) 各灌溉模式的水稻植株叶片、茎鞘和根部干物质积累量的变化规律大致相同, 均呈倒“Ⅴ”型曲线, 且最大值出现在抽穗开花期。从抽穗开花期到黄熟期, 由叶片和茎鞘向穗转运的干物质量分别为2.40 g/穴、4.08 g/穴, 远高于间歇灌溉模式和常规淹灌模式。
(3) 不同灌溉模式之间的水稻各阶段耗水量差异性均显著(P < 0.05), 大小关系均为常规淹灌模式>间歇灌溉模式>控制灌溉模式, 从拔节孕穗期到黄熟期, 水稻的耗水模系数大小关系为控制灌溉模式>间歇灌溉模式>常规淹灌模式。从全生育期来看, 水稻各处理生育期的叶面蒸腾强度、作物需水强度及田间耗水强度均呈现先上升后下降的趋势, 从拔节孕穗期开始, 叶面蒸腾强度在水稻总耗水量中占比最大。
| [1] |
国家统计局. 中华人民共和国2016年国民经济和社会发展统计公报. http://www.stats.gov.cn/tjsj/zxfb/201702/t20170228_1467424.html.
|
| [2] |
展茗, 曹凑贵, 汪金平, 蔡明历, 袁伟玲. 复合稻田生态系统温室气体交换及其综合增温潜势. 生态学报, 2008(11): 5461-5468. DOI:10.3321/j.issn:1000-0933.2008.11.030 |
| [3] |
李自民, 宋照亮, 姜培坤. 稻田生态系统中植硅体的产生与积累——以嘉兴稻田为例. 生态学报, 2013, 33(22): 7197-7203. |
| [4] |
魏永霞, 侯景翔, 吴昱, 刘慧, 汝晨, 杨军明. 不同水分管理旱直播水稻生长生理及节水效应. 农业机械学报, 2018, 49(8): 253-264. |
| [5] |
魏永霞, 汝晨, 吴昱, 刘慧, 杨军明, 侯景翔. 寒地黑土区水稻耗水特性及其对水分利用效率的影响. 农业机械学报, 2019, 50(4): 245-254. |
| [6] |
魏永霞, 汝晨, 吴昱, 刘慧, 杨军明, 侯景翔. 黑土区水稻生长生理特性与产量对耗水过程的响应. 农业机械学报, 2018, 49(9): 214-225. |
| [7] |
曹慧, 王孝威, 韩振海, 许雪峰, 王永章. 水分胁迫诱导平邑甜茶叶片衰老期间内肽酶与活性氧累积的关系. 中国农业科学, 2004, 37(2): 274-279. DOI:10.3321/j.issn:0578-1752.2004.02.019 |
| [8] |
要世瑾, 杜光源, 牟红梅, 栾翔宇, 马鸿雁, 刘嘉男, 刘梦达, 齐笑, 何建强. 基于核磁共振技术检测小麦植株水分分布和变化规律. 农业工程学报, 2014, 30(24): 177-186. DOI:10.3969/j.issn.1002-6819.2014.24.021 |
| [9] |
Yoshida S. Molecular regulation of leaf senescence. Current Opinion in Plant Biology, 2003, 6(1): 79-84. DOI:10.1016/S1369526602000092 |
| [10] |
Granier C, Tardieu F. Water deficit and spatial pattern of leaf development. Variability in responses can be simulated using a simple model of leaf development. Plant Physiology, 1999, 119(2): 609-620. DOI:10.1104/pp.119.2.609 |
| [11] |
杨启良, 张富仓, 刘小刚, 王玺, 张楠, 戈振扬. 植物水分传输过程中的调控机制研究进展. 生态学报, 2011, 31(15): 4427-4436. |
| [12] |
胡继超, 姜东, 曹卫星, 罗卫红. 短期干旱对水稻叶水势、光合作用及干物质分配的影响. 应用生态学报, 2004, 15(1): 63-67. DOI:10.3321/j.issn:1001-9332.2004.01.014 |
| [13] |
甄博, 郭相平, 陆红飞. 旱涝交替胁迫对水稻分蘖期根解剖结构的影响. 农业工程学报, 2015, 31(9): 107-113. |
| [14] |
吴岳轩, 吴振球. 播期对亚种间杂交稻根系形态发育和生理活性的影响. 作物学报, 1996, 22(2): 178-184. DOI:10.3321/j.issn:0496-3490.1996.02.010 |
| [15] |
徐国伟, 陆大克, 孙会忠, 王贺正, 李友军. 干湿交替灌溉与施氮耦合对水稻根际环境的影响. 农业工程学报, 2017, 33(4): 186-194. |
| [16] |
张伟明, 孟军, 王嘉宇, 范淑秀, 陈温福. 生物炭对水稻根系形态与生理特性及产量的影响. 作物学报, 2013, 39: 1445-1451. |
| [17] |
郑天翔, 唐湘如, 罗锡文, 黎国喜, 王在满, 舒时富, 陈伟通. 不同灌溉方式对精量穴直播超级稻生产的影响. 农业工程学报, 2010, 26(8): 52-55. DOI:10.3969/j.issn.1002-6819.2010.08.008 |
| [18] |
郝树荣, 郭相平, 张展羽. 水分胁迫及复水对水稻冠层结构的补偿效应. 农业机械学报, 2010, 41(3): 52-55. |
| [19] |
路兴花, 吴良欢, 庞林江. 不同土壤水分含量对水稻水分利用特征的影响. 中国水稻科学, 2009, 23(2): 186-190. DOI:10.3969/j.issn.1001-7216.2009.02.012 |
| [20] |
王唯逍, 刘小军, 田永超, 姚霞, 曹卫星, 朱艳. 不同土壤水分处理对水稻光合特性及产量的影响. 生态学报, 2012, 32(22): 7053-7060. |
| [21] |
刘笑吟, 王冠依, 杨士红, 徐俊增, 王乙江. 不同时间尺度节水灌溉水稻腾发量特征与影响因素分析. 农业机械学报, 2016, 47(8): 91-100. |
| [22] |
Asseng S, Ritchie J T, Smucker A J M. Root growth and water uptake during water deficit and recovering in wheat. Plant&Soil, 1998, 201(2): 265-273. |
| [23] |
曹生奎, 冯起, 司建华, 常宗强, 席海洋, 卓玛错. 植物水分利用效率研究方法综述. 中国沙漠, 2009, 29(5): 853-858. |
| [24] |
张亚琦, 李淑文, 付巍, 文宏达. 施氮对杂交谷子产量与光合特性及水分利用效率的影响. 植物营养与肥料学报, 2014, 20(5): 1119-1126. |