 2021, Vol. 41
2021, Vol. 41文章信息
- 叶文建, 杜萍, 寿鹿, 蔡小霞, 陈悦, 汤雁滨, 廖一波, 黄伟, 刘小涯
- YE Wenjian, DU Ping, SHOU Lu, CAI Xiaoxia, CHEN Yue, TANG Yanbin, LIAO Yibo, HUANG Wei, LIU Xiaoya
- 舟山海域大中型浮游动物群落时空变化及受控要素
- Spatio-temporal variation of marco and mesozooplankton communities and the controlling factors around Zhoushan archipelago
- 生态学报. 2021, 41(1): 254-267
- Acta Ecologica Sinica. 2021, 41(1): 254-267
- http://dx.doi.org/10.5846/stxb201912242781
-
文章历史
- 收稿日期: 2019-12-24
- 网络出版日期: 2020-11-19






舟山海域拥有我国最大的近海渔场, 海洋渔业资源十分丰富。浮游动物作为渔业资源的重要饵料, 可以将较低营养级生物的物质和能量传递到渔业资源, 对渔业资源的维持具有重要意义。浮游动物对环境变化敏感, 水质变化、赤潮发生和渔业资源结构变动都可能影响浮游动物数量和结构, 因此浮游动物的群落结构、种类组成和季节变化特征对海洋环境和渔业资源的研究与保护具有重要意义[1-3]。
舟山海域位于长江和钱塘江两大径流交汇区, 陆地径流带来了丰富的陆源营养物质; 另外, 受台湾暖流和沿岸寒流的交汇以及沿岸上升流的作用, 表底层海水的混合使得舟山海域营养丰富、温度稳定, 利于浮游生物的生长, 为鱼类提供了丰富的天然的饵料。舟山及其附近海域岛屿众多, 海水交换状况良好, 多样的生态环境同时也为舟山渔场的海洋生物提供了优良的的产卵场、育幼所和庇护场[4]。然而, 以往的过度捕捞使得渔业资源发生衰退和更替; 随着2011年舟山新区的发展, 各类港口、船舶工程、围填海工程的建设加重了舟山海域的环境污染问题, 该海域海洋生态系统正面临巨大威胁[5-8]。
目前, 关于舟山海域浮游动物的研究报道都主要是针对夏季或其他单个季节[9-13], 且研究区域更多的以嵊泗岛和东极岛为中心[14-15], 对舟山本岛附近海域的浮游动物鲜有报道。因此, 本文根据2014年夏季、秋季, 2016年春季以及2017年冬季对舟山岛附近海域浮游动物的采样调查, 研究舟山岛附近海域浮游动物群落的时空变化特征及主要驱动因素, 为今后舟山海域的海洋生态环境修复和舟山渔业资源的保护等研究提供基础资料。
1 研究区域与方法 1.1 调查海域和站位布设2014年7月和10月、2016年5月、2017年2月在舟山岛附近海域(121.82°—122.35°E, 29.80°—30.22°N)进行4个航次的海洋浮游动物生态调查, 其中夏季(7月)、秋季(10月)各布设25个站位(Z1—Z25), 春季(5月)、冬季(2月)各布设33个站位(Z1—Z32和P2)(图 1)。

|
| 图 1 舟山海域调查站位图 Fig. 1 Sampling stations in the waters nearby Zhoushan Island Z:代表站点 |
调查方法均按照《海洋调查规范—海洋生物调查》(GB12763.6-91, 2007)[16]进行。浮游动物样品采集使用浅水Ⅰ型浮游生物网(网口内径50 cm, 网长145 cm, 筛绢孔径为505 μm, 网口安装德国HYDRO-BIOS-数字网口流量计)进行由底至表层垂直拖网一次采集获得。采集的浮游动物样品装入容积为1000 mL的塑料瓶中, 加入甲醛溶液至体积分数为5%固定保存, 根据流量计转数计算滤水量。实验室内挑去杂物后, 用精度为万分之一的天平称量浮游动物湿重, 在体视显微镜下对样品进行鉴定和计数, 结合湿重、个体数和滤水量分别计算湿重生物量和丰度。Chl a则采集现场水样带回实验室进行过滤, 并将滤膜保存在5 mL体积比为90%的丙酮中, 避光冷冻保存24 h后通过荧光计测定。
环境参数同步测定, 其中盐度(盐度计法)、温度(水温计法)、pH(玻璃电极法)和水深(垂直铅鱼法)由现场仪器直接测得; 化学参数如溶解氧(Dissolved oxygen, DO)采集水样保存于50 mL棕色玻璃瓶内并加入1 mL碘化钾和1 mL氯化锰固定, 磷酸盐(Dissolved inorganic phosphorus, DIP)、无机氮(Dissolved inorganic nitrogen, DIN)、硅酸盐(Dissolved silicate, DSi)、悬浮物则采集水样保存于2000 mL塑料瓶, 所有水样黑暗保存带回实验室按照《海洋监测规范—海水分析》(GB12763.4-91, 2007)[17]方法测定。
1.3 数据分析优势种更替率(R)[18]:
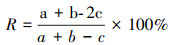
|
式中, a、b为相邻两期优势种种数, c为共同优势种种数。
采用Surfer 11绘制站位图, 浮游动物丰度、生物量和温盐分布图。根据适温或适盐范围将浮游动物分为不同的生态类群[14, 19]。浮游动物优势度和群落多样性指数用Primer 6.0软件处理得到, 并基于物种丰度对各季节浮游动物群落进行相似性分析和单因素方差分析(Analysis of similarities, ANOSIM)。浮游动物优势度和群落多样性计算公式如下[20]:
(1) 浮游动物种类优势度计算公式(Y):
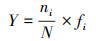
|
式中, ni为第i种浮游动物的个体数, N为浮游动物总个体数; fi为第i种浮游动物在采样站位中出现的频率。
(2) Margalef的种类丰富度指数(d):

|
式中, S为浮游动物总的种类数, N为所有浮游动物总的个体数。
(3) Shannon-Wiener多样性指数(H′):
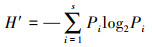
|
式中, Pi为第i种浮游动物的个数与总个体数的比值, S为浮游动物种类数。
(4) Pielou均匀度指数(J′):
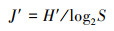
|
式中, S为浮游动物种类数。
采用SPSS 20.0中的单因素非参数方差分析法(Kruskal-Wallis, K-W检验)检验浮游动物生物量、丰度的季节间差异, 当P < 0.05时, 再进行成对比较。采用SPSS 18软件对4个季节环境因子和浮游动物丰度进行Spearman相关性分析; 选取各个季节浮游动物的主要物种(相对丰度≥0.01), 利用CANOCO对主要物种的丰度进行去趋势对应分析(Detrended Correspondence Analysis, DCA)分析, 根据DCA分析结果的最大梯度长度值, 值若小于3, 则选用冗余分析(Redundancy Analysis, RDA)分析方法; 若介于3和4之间, 则RDA和典范对应分析(Canonical Correlation Analysis, CCA)均可; 若大于4, 则选用CCA分析方法分析4个季节的环境理化因子和物种丰度的对应关系, 并且进行RDA或CCA分析前, 通过前项选择(RDA-forward selection)来筛选出影响显著的理化因子(P<0.05)。
2 结果 2.1 种类组成和优势种调查海域四季共鉴定出浮游动物88种和浮游幼体19类, 桡足类是四季的主要生物类群, 共鉴定到42种; 其次是水螅水母类, 为13种。夏季的浮游动物种类四季最多, 共67种, 其中有水螅水母类8种和浮游幼体15类; 春季和秋季其次, 分别有共浮游动物50种和45种, 其中分别包含浮游幼体9类和7类; 冬季种类数共21种, 为四季最少, 且仅有1类浮游幼体。春-夏、夏-秋、秋-冬和冬-春相邻季节的优势种更替率分别为75%、80%、100%和60%。
调查海域春季浮游动物优势种共4种, 中华哲水蚤(Calanus sinicus)为该海域最主要的优势种, 其余依次为捷氏歪水蚤(Tortanus derjugini)、百陶带箭虫(Zonosagitta bedoti)和虫肢歪水蚤(T. vermiculus); 优势种以暖温带近海物种为主, 其次为近岸低盐种。夏季浮游动物优势种共6种, 背针胸刺水蚤(Centropages dorsispinatus)为该海域最主要的优势种, 其余依次为圆唇角水蚤(Labidocera rotunda)、中华哲水蚤(C. sinicus)、精致真刺水蚤(Euchaeta concinna)、百陶带箭虫(Z. bedoti)和真刺唇角水蚤(L. euchaeta); 优势种以近岸低盐和暖温带近海物种为主。秋季浮游动物优势种共6种, 背针胸刺水蚤、百陶带箭虫依然为该海域优势种, 双生水母(Diphyes chamissonis)、瓜水母(Beroe cucumis)为新的优势种, 其他依次为糠虾幼体(Mysidacea larva)和箭虫幼体(Sagittid larva)。冬季浮游动物优势种共3种, 分别为真刺唇角水蚤(L. euchaeta)、捷氏歪水蚤(T. derjugini)和中华哲水蚤(C. sinicus), 与秋季的优势种类完全不同(表 1)。
| 物种 Species |
春季 Spring |
夏季 Summer |
秋季 Autumn |
冬季 Winter |
| 中华哲水蚤Calanus sinicus | 0.511 | 0.079 | — | 0.046 |
| 捷氏歪水蚤Tortanus derjugini | 0.148 | / | — | 0.069 |
| 百陶带箭虫Zonosagitta bedoti | 0.051 | 0.026 | 0.099 | — |
| 虫肢歪水蚤Tortanus vermiculus | 0.026 | / | / | — |
| 背针胸刺水蚤Centropages dorsispinatus | — | 0.404 | 0.255 | / |
| 圆唇角水蚤Labidocera rotunda | / | 0.092 | — | / |
| 真刺唇角水蚤Labidocera euchaeta | — | 0.022 | — | 0.113 |
| 精致真刺水蚤Euchaeta concinna | — | 0.032 | — | / |
| 双生水母Diphyes chamissonis | / | / | 0.097 | / |
| 瓜水母Beroe cucumis | / | — | 0.071 | / |
| 糠虾幼体Mysidacea larva | / | — | 0.058 | / |
| 箭虫幼体Sagittid larva | / | — | 0.048 | / |
| “—”:优势度小于0.02;“/”:未发现 | ||||
对舟山海域浮游动物丰度和生物量进行季节间K-W检验, 结果显示:浮游动物丰度和生物量季节间差异显著(P < 0.001)。经过两两成对比较, 除秋季和冬季间浮游动物的丰度水平无差别(调整后P=1.000)外, 其他季节间均有显著差异(调整后P<0.050);而生物量水平除春季和秋季间无差别(调整后P=1.000)外, 其他季节间均有显著差异(调整后P<0.050)(表 2)。
| 样本1-样本2 Sample1-Sample2 |
丰度Abundance | 生物量Biomass | |||
| P | 调整后P值 Adjusted P |
P | 调整后P值 Adjusted P |
||
| 春季-夏季Spring-Summer | 0.001 | 0.007 | 0.001 | 0.007 | |
| 春季-秋季Spring-Autumn | 0.006 | 0.035 | 0.745 | 1.000 | |
| 春季-冬季Spring-Winter | 0.000 | 0.003 | 0.000 | 0.000 | |
| 夏季-秋季Summer-Autumn | 0.000 | 0.000 | 0.005 | 0.032 | |
| 夏季-冬季Summer-Winter | 0.000 | 0.000 | 0.000 | 0.000 | |
| 秋季-冬季Autumn-Winter | 0.501 | 1.000 | 0.000 | 0.000 | |
舟山海域浮游动物季节平均生物量和平均丰度均为夏季最高, 分别达到176.34 mg/m3和143.97个/m3, 平均生物量冬季最低, 仅7.21 mg/m3, 平均丰度秋季最低, 仅21.38个/m3。
春季丰度和生物量的最高值(760.26个/m3和1164.87 mg/m3)均位于朱家尖岛西南海域的Z26号站, 主要是春季优势种中华哲水蚤在朱家尖附近海域大量聚集所致; 丰度和生物量最低值(10.13个/m3和7.45 mg/m3)位于册子岛和舟山本岛之间的附近海域, 主要物种为捷氏歪水蚤和虫肢歪水蚤。浮游动物丰度和生物量总体分布趋势均为东侧高于西侧海域(图 2, 图 3)。

|
| 图 2 四个季节的浮游动物生物量和盐度平面分布图 Fig. 2 Horizontal distribution of zooplankton biomass and salinity Z:代表站点 |

|
| 图 3 四个季节的浮游动物丰度和温度平面分布图 Fig. 3 Horizontal distribution of zooplankton abundance and temperature Z:代表站点 |
夏季丰度和生物量的高值区(383.76个/m3和349.87 mg/m3)均位于外海侧秀山岛的附近海域, 主要构成物种是背针胸刺水蚤、糠虾幼体、双刺唇角水蚤和中华哲水蚤。整体上夏季舟山海域各站位生物量和丰度相似性较高, 舟山外海侧和内海侧丰度存在差异, 外海侧丰度相对较高, 生物量分布则表现为舟山岛东部海域高于西部海域, 南部与北部差异不明显, 其中册子岛附近海域生物量和丰度都较低(图 2, 图 3)。
秋季丰度和生物量均明显低于夏季, 丰度和生物量的最高值(76.19个/m3和452.86 mg/m3)均位于金塘岛与舟山本岛之间海域的Z16号站, 主要种类为背针胸刺哲水蚤。秋季整个生物量分布特征不明显, 调查海域南部海域生物量偏低, 而金塘岛附近海域生物量相对略高, 空间差异较春、夏季小; 丰度总体上舟山本岛北部、南部以及金塘岛西北部海域浮游动物丰度较低, 舟山本岛东部、金塘岛东南部以及金塘岛册子岛之间的附近海域浮游动物丰度较高(图 2, 图 3)。
冬季平均丰度达到四季最低, 但生物量略高于秋季。丰度和生物量总体上为舟山与宁波之间的内海侧大于外海侧, 金塘岛附近海域浮游动物生物量和丰度最高(图 2, 图 3)。
2.3 群落特征指数舟山海域浮游动物的群落特征指数如表 3所示, 其中平均丰富度指数春季、夏季高于秋季、冬季, 其中夏季最高(5.26), 冬季最低(1.61);平均均匀度指数秋、冬季高于春季、夏季, 其中冬季最高(0.92), 春季最低(0.58);平均多样指数夏季、秋季高于春季、冬季, 其中夏季最高(3.03), 冬季最低(1.71)。
| 季节 Season |
群落特征指数 Community parameters |
最小值 Min |
最大值 Max |
平均值±标准差 Mean ± SD |
| 春季Spring | d | 0.82 | 5.49 | 3.16 ± 1.36 |
| J′ | 0.23 | 0.93 | 0.58 ± 0.21 | |
| H′ | 0.85 | 3.60 | 2.05 ± 0.82 | |
| 夏季Summer | d | 2.55 | 6.59 | 5.26 ± 1.07 |
| J′ | 0.38 | 0.84 | 0.64 ± 0.14 | |
| H′ | 1.80 | 4.27 | 3.03 ± 0.71 | |
| 秋季Autumn | d | 1.49 | 4.12 | 2.83 ± 0.85 |
| J′ | 0.66 | 0.96 | 0.83 ± 0.06 | |
| H′ | 1.92 | 3.70 | 2.82 ± 0.53 | |
| 冬季Winter | d | 0.91 | 2.49 | 1.61 ± 1.52 |
| J′ | 0.69 | 1.00 | 0.92 ± 0.40 | |
| H′ | 0.92 | 2.52 | 1.71 ± 0.87 | |
| d: Margalef种类丰富度指数; J′: Pielou均匀度指数; H′: Shannon-Wiener多样性指数 | ||||
将各季节所有站位共分为A区(Z1—Z8和P2)、B区(Z9—Z16)、C区(Z17—Z25)和D区(Z26—Z32)四个区(图 1)。其中春季和冬季可分为A、B、C和D 4个组, 夏秋可分为A、B、C 3个组。ANOSIM分析结果显示, 春季D区与A、C区空间差异极显著, A、B和C区相互之间空间差异显著; 夏季仅A区与C区空间差异显著; 秋季仅B区与C区空间差异显著; 冬季A区与D区空间差异极显著, 与B、C区空间差异显著, 但B、C和D区相互之间空间差异不显著(表 4)。
| 区组 Groups |
春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | |||||||
| R | P | R | P | R | P | R | P | ||||
| A, B | 0.153 | 0.044 | 0.185 | 0.057 | 0.117 | 0.081 | 0.371 | 0.010 | |||
| A, C | 0.213 | 0.015 | 0.260 | 0.030 | 0.074 | 0.155 | 0.215 | 0.012 | |||
| A, D | 0.957 | 0.002 | 0.673 | 0.006 | |||||||
| B, C | 0.260 | 0.017 | 0.049 | 0.180 | 0.186 | 0.013 | 0.040 | 0.291 | |||
| B, D | 0.203 | 0.077 | 0.238 | 0.059 | |||||||
| C, D | 0.882 | 0.003 | -0.077 | 0.733 | |||||||
| ANOSIM:相似性分析Analysis of similarities | |||||||||||
整合四季所有浮游动物丰度, 分析同一分组在不同季节间的组间差异, 其中春季为组A1、B1、C1和D1, 夏季为组A2、B2和C2, 秋季为A3、B3和C3, 冬季为组A4、B4、C4和D4。结果表明同一分组在不同季节间的组间差异均极显著(表 5)。
| 区组 Groups |
R | P |
| A1, A2 | 0.996 | 0.001 |
| A1, A3 | 0.979 | 0.002 |
| A1, A4 | 0.766 | 0.002 |
| A2, A3 | 0.974 | 0.001 |
| A2, A4 | 0.974 | 0.001 |
| A3, A4 | 0.961 | 0.002 |
| B1, B2 | 0.729 | 0.001 |
| B1, B3 | 0.768 | 0.001 |
| B1, B4 | 0.402 | 0.001 |
| B2, B3 | 0.900 | 0.001 |
| B2, B4 | 0.637 | 0.001 |
| B3, B4 | 0.708 | 0.001 |
| C1, C2 | 0.965 | 0.001 |
| C1, C3 | 0.986 | 0.001 |
| C1, C4 | 0.398 | 0.001 |
| C2, C3 | 0.967 | 0.001 |
| C2, C4 | 0.494 | 0.001 |
| C3, C4 | 0.655 | 0.001 |
| D1, D4 | 0.478 | 0.008 |
| 字母下标数字1、2、3、4分别代表春季、夏季、秋季和冬季 | ||
舟山海域温度空间分布特征为春季南侧高于北侧, 外海侧高于内海侧; 夏、秋季温度分布相似, 均为内海测高于外海测, 冬季各站位温度均较低, 但金塘和朱家尖两岛屿南部的温度均比其他站位要高(图 3, 表 6)。盐度空间分布均为东南侧高于西北侧, 但是冬季各站位盐度相差不大, 西北侧站位盐度较高(图 2, 表 6)。
| 环境因子 Environmental factor |
春季 Spring |
夏季 Summer |
秋季 Autumn |
冬季 Winter |
| 深度Depth/m | 18.2000±13.71 | 15.3500±6.88 | 12.9200±5.26 | 14.6400±11.13 |
| 温度Temp/℃ | 16.6200± 0.50 | 25.1400±0.85 | 20.2200±0.40 | 10.5700±0.40 |
| 盐度Sal/‰ | 25.1000±1.84 | 25.4300±2.27 | 21.3500±1.94 | 23.4000±2.82 |
| pH | 7.9600±0.05 | 7.9800±0.06 | 8.0800±0.01 | 8.2900±0.78 |
| 溶解氧DO/(mg/L) | 8.1389±0.5524 | 6.0515±0.5224 | 7.7301±0.2870 | 9.9844±0.66 |
| 悬浮物SS/(mg/L) | 527.0645±300.3516 | 204.4576±154.8737 | 751.1200±391.8558 | 522.0654±252.2924 |
| 叶绿素a Chl a/(μg/L) | 0.9010±0.7556 | 0.7059±0.2791 | 0.2104±0.0782 | 0.9009±0.0704 |
| 溶解硅酸盐DSi/(mg/L) | 1.3264±0.2419 | 1.2542±0.2687 | 1.7317±0.2303 | 1.9056±2.8642 |
| 溶解无机磷DIP/(mg/L) | 0.0363±0.0081 | 0.0382±0.0069 | 0.0512±0.0055 | 0.6985±3.3003 |
| 溶解无机氮DIN/(mg/L) | 0.7325±0.1409 | 0.6205±0.1582 | 1.0614±0.1474 | 2.9952±10.9218 |
| Temp:温度Temperature; Sal:盐度Salinity; DO:溶解氧Dissolved oxygen; SS:悬浮物Suspended solids; Chl a:叶绿素a Chlorophyll a; DIP:溶解磷酸盐Dissolved inorganic phosphorus; DIN:溶解无机氮Dissolved inorganic nitrogen; DSi:溶解硅酸盐Dissolved silicate | ||||
浮游动物丰度和多种环境因子相关性显著, 其中春季主要与Sal和Chl a极显著正相关、与DSi和DIN极显著负相关性; 夏季与Chl a和盐度极显著正相关, 与Temp和营养盐极显著负相关; 秋季与Temp极显著正相关, 与Sal显著正相关, 与SS、Depth和营养盐显著负相关; 冬季仅与DO显著负相关(表 7)。
| 深度 Depth |
温度 Temp |
盐度 Sal |
pH | 溶解氧 DO |
悬浮物 SS |
叶绿素a Chl a |
溶解硅酸盐 DSi |
溶解磷酸盐 DIP |
溶解无机氮 DIN |
|
| 春季Spring | -0.162 | 0.294 | 0.481 | 0.405 | -0.210 | -0.373 | 0.620 | -0.528 | -0.453 | -0.530 |
| 夏季Summer | -0.074 | -0.596 | 0.528 | 0.482 | -0.307 | -0.239 | 0.559 | -0.628 | -0.636 | -0.387 |
| 秋季Autumn | -0.436 | 0.548 | 0.462 | 0.320 | -0.249 | -0.449 | -0.199 | -0.504 | -0.510 | -0.640 |
| 冬季Winter | -0.378 | 0.187 | 0.165 | -0.095 | -0.390 | -0.210 | 0.131 | -0.161 | -0.072 | -0.125 |
为了发现影响浮游动物四季分布的主要因子, 整合四季的环境因子和主要物种(相对丰度≥0.01)丰度数据并对主要物种丰度进行DCA分析, 最大梯度长度为5.82, 后续进行CCA分析, 同时通过对理化因子进行前项选择并筛选出6个主要的理化因子(P < 0.05)。结果表明:前两轴6个环境因子累积解释了26.96%的物种变量, 其中温度、盐度、Chl a和溶解氧是影响浮游动物四季变化的主要环境因子, 营养盐(DIP和DIN)的影响则相对较低。Temp和DO是春、冬季与夏、秋季分化的主要影响因素, 其次Sal和Chl a是夏季和秋季分化的主要影响因素(图 4)。

|
| 图 4 四季主要浮游动物和站位与环境因子的CCA分析 Fig. 4 CCA ordination of main zooplankton and sample sites with environmental factors |
春季筛选出4个环境因子与7个主要物种进行RDA分析(图 5), 前两轴4个环境因子累积解释了浮游动物79.47%的物种变量, 溶解无机氮、温度和盐度是影响春季浮游动物空间变化的主要环境因子(表 8)。春季分布于B区和D区的站位靠近第一轴的正方向, 与温度和盐度表现出较强的相关性; 而春季主要优势种中华哲水蚤和百陶带箭虫表现出与温度和盐度的强相关性; 分布于A区和C区的站位更靠近第一轴的负方向, 与盐度表现出更强的负相关性, 优势种中捷氏歪水蚤和虫肢歪水蚤是适低盐种类, 也表现出与盐的强负相关性。

|
| 图 5 主要浮游动物与环境因子4个季节的RDA分析 Fig. 5 RDA ordination of main zooplankton with environmental factors in four seasons RDA:冗余分析Redundancy Analysis |
| 特征值Eigenvalue | 相关系数Correlation coefficient | 解释变量Explained variation | ||||||
| 轴一 SPEC AX1 |
轴二 SPEC AX2 |
轴一 SPEC AX1 |
轴二 SPEC AX2 |
轴一 SPEC AX1 |
轴二 SPEC AX2 |
|||
| 春季Spring | 0.7714 | 0.0233 | 0.9847 | 0.5521 | 77.1400 | 79.4700 | ||
| 夏季Summer | 0.2925 | 0.0528 | 0.8355 | 0.7569 | 29.2500 | 34.5400 | ||
| 秋季Autumn | 0.1672 | 0.0595 | 0.7209 | 0.6673 | 16.7200 | 22.6700 | ||
| 冬季Winter | 0.1566 | 0.0968 | 0.6700 | 0.6837 | 15.6600 | 25.3400 | ||
| 四季All Seasons | 0.6033 | 0.3025 | 0.9444 | 0.8161 | 17.9600 | 26.9600 | ||
| RDA:冗余分析Redundancy Analysis;CCA:典范对应分析Canonical Correlation Analysis;SPEC AX:排序轴spec axis | ||||||||
夏季筛选出4个环境因子与12个主要物种进行RDA分析(图 5), 前两轴4个环境因子累积解释了浮游动物34.54%的物种变量, 温度、盐度、溶解无机磷和Chl a是影响夏季浮游动物空间变化的主要环境因子(表 8)。夏季分布在B区和C区的站位靠近第一轴负方向, 与温度和溶解无机磷表现出较强的相关性; 而夏季主要优势种中背针胸刺水蚤和小拟哲水蚤也表现出与温度有较强相关性; 分布在A区的站位靠近第一轴的正方向, 与Chl a和盐度表现出较强相关性; 而主要优势种中的精致真刺水蚤、真刺唇角水蚤、中华哲水蚤、百陶带箭虫和瓜水母幼体等则表现出与Sal和Chl a较强相关性。
秋季筛选出2个环境因子与9个主要物种进行RDA分析(图 5), 前两轴2个环境因子累积解释了浮游动物22.67%的物种变量, 溶解无机氮和Chl a是影响浮游动物空间变化的主要环境因子(表 8)。
冬季筛选出4个环境因子与6个主要物种进行RDA分析(图 5), 前两轴4个环境因子累积解释了浮游动物25.34%的物种变量, 溶解氧、悬浮物、温度和Chl a是影响浮游动物分布的主要环境因子(表 8)。秋、冬季站位多数站位都比较靠近轴心, 与环境因子的相关性不高, 但部分朱家尖岛和金塘岛海域的站位与环境因子(Chl a)表现出极强相关性。
3 讨论 3.1 浮游动物群落特征及季节变化四个航次时间跨度较大, 但是根据舟山以往的相关历史文献发现, 舟山本岛海域浮游动物虽然丰度和生物量呈缓慢下降趋势, 但优势种组成却并无明显变化, 因此2—3年的时间跨度对舟山浮游动物季节变化规律的揭示影响应该不大。根据对舟山海域4个季度浮游动物的调查结果显示, 浮游动物的群落特征存在明显的季节变化。浮游动物CCA分析(图 4)发现温度、盐度、Chl a和溶解氧是影响浮游动物季节分化的重要因素。温度是浮游动物季节变化的主要因素, 其次盐度和Chl a是夏季和秋季分化的主要影响因素。舟山海域浮游动物春季受温度影响浮游幼体开始大量繁殖, 夏季暖流带来大量高温高盐的海水的同时也带来了多种水母, 秋季浮游植物减少不利浮游动物的生长繁殖, 冬季气温全年最低, 所以暖水种骤减, 浮游动物主要为近岸种或河口种。
春季水温上升, 水母及各类浮游动物开始繁殖, 适温更高的近岸低盐种(捷氏歪水蚤等)和暖温带近海种(中华哲水蚤等)在该海域浮游动物中占主导地位, 直接影响着舟山海域浮游动物的丰度和生物量分布。另外有研究表明[21]舟山海域春季浮游植物由硅藻门直链藻属和甲藻门原甲藻属两大主要类群组成, 种类多且数量分布不均衡。春季, 浮游植物开始进入快速繁殖生长的时期, 其分布间接影响浮游动物的种类分布。部分站位中华哲水蚤成为绝对优势种, 造成春季调查海区浮游动物丰度高但却种类少、分布不均匀的情况, 所以平均均匀度指数四季最低。
夏季水温达到四季最高, 浮游植物生长高峰期, 为浮游动物的生长繁殖提供充足的饵料, 幼体种类达到四季中最多。此时, 长江和钱塘江处于丰水期[22-24], 冲淡水增多, 河口区高温低盐, 盐度梯度明显, 该海域同时又受台湾暖流的影响[25-26], 整个海域呈现多样的温盐环境, 高温高盐的种类增加。浮游动物对温盐梯度敏感[27-28], 高温高盐种主要分布在外海区, 而近岸低盐种和高温低盐种主要分布在河口区。夏季浮游动物主要组成除近岸低盐种外, 还有寒暖流交汇区的高温高盐种和河口区的高温低盐种, 主要种类有背针胸刺水蚤、圆唇角水蚤和各类水螅水母。所以夏季各站位浮游动物的种类数、生物量、平均丰富度和平均多样性均为四季中最高。
秋季, 随着水温下降, 浮游植物丰度减小, 浮游动物生长和繁殖速率也随之下降, 夏季的暖水种、水螅水母类和浮游幼体大量减少, 取而代之的是近岸低盐种和暖温带近岸类群, 主要贡献者为百陶带箭虫、双生水母和瓜水母, 浮游动物的种类数、生物量、丰度和均匀度也随之下降。
冬季气温持续下降, 不适宜浮游动物的繁殖和暖水性浮游生物的生存, 所以冬季浮游动物和浮游幼体种类数四季最低, 主要贡献种为中华哲水蚤和真刺唇角水蚤。秋季的平均生物量高于冬季而平均丰度低于冬季, 可能与冬季浮游动物优势类群是生物量较小的种类(如真刺唇角水蚤), 而生物量较大的暖水种(如百陶带箭虫、糠虾幼体等)在冬季消失导致的。
3.2 浮游动物群落分布空间差异本研究将研究海域分为4个区域:A区是舟山本岛外海侧秀山岛区域的站位, 受台湾暖流影响; B区是舟山本岛内海侧的站位, 受近岸上升流影响较大; C区站位是舟山本岛西北部金塘-册子岛附近区域的站位, 主要受两江河口冲淡水影响; D区站位是位于舟山本岛东南部朱家尖岛附近区域的站位。B区和D区未检验到空间差异, 表明从浮游动物群落角度, 朱家尖岛与本岛海域无差异, 即研究海域可分为外海区(A区)、舟山近岸区(B区和D区)和河口区(C区)。ANOSIM结果显示4个季节的浮游生物群落均存在空间差异, 但春季的区域间差异最明显, 其次是冬季, 夏季仅外海区与河口区差异显著, 秋季仅舟山近岸区与河口区差异显著, 表明该海域暖季(夏季和秋季)的水体混合较冷季(春季和冬季)强。主要由于影响河口区(C区)的两江冲淡水和影响外海区(A区)的台湾暖流均在暖季较强[10-11]。受影响海流的季节变化影响, 浮游动物各季节的空间分布特征也不同。
目前已有研究显示温度、盐度、溶解氧、pH会直接影响浮游动物分布, 而SS和营养盐会通过影响浮游植物间接影响浮游动物分布[29]。在已有的舟山浮游动物研究中, 有研究[11]发现浮游动物数量与温度、盐度密切相关, 但没有相关性分析结果验证; 有研究[12]发现浮游动物生物量在春季与叶绿素负相关, 在秋季与叶绿素无相关性。由于舟山海域四季温度变化明显, 海水盐度梯度主要受径流和寒暖流影响, 所以主导浮游动物分布的影响因素每个季节不同。根据本文环境因子的Spearman相关性分析表明:除冬季外, 其它季节浮游动物丰度均与盐度和营养盐显著相关, Chl a在春、夏季显著相关, 温度仅夏、秋季显著相关, 冬季仅与溶解氧显著相关; 而根据四个季节的RDA分析表明春季浮游动物群落空间分布主要受盐度的影响, 夏季受温度、盐度和Chl a的影响, 秋季主要受Chl a的影响, 冬季主要受悬浮物和溶解氧的影响, 而营养盐对每个季节的浮游动物群落分布都有一定的影响。
温盐是影响浮游动物群落结构区域差异的重要因素[30-32]。5月份是东北季风向东南季风转换期, 春季调查期间, 该海域主要受台湾暖流和东南季风的影响, 舟山海域南部温度和盐度高于北部, 所以春季的优势种中华哲水蚤和百陶带箭虫主要分布在舟山南部朱家尖海域, 而捷氏歪水蚤则分布在低温低盐的北部金塘海域范围。如春季中华哲水蚤在朱家尖附近的Z26站位丰度极高, 导致该区域浮游动物生物量和丰度春季均大于夏季。
夏季水温上升, 陆地河流进入汛期, 台湾暖流自南向北楔入直抵沿岸水域, 因此两江冲淡水、寒暖流和近岸流等水团的影响范围均显著扩大, 舟山海域形成了长江河口盐度梯度区、钱江河口盐度梯度区、寒暖流交汇区和沿岸流区等多种独特的海洋物理环境。夏季整体温度梯度从东南到西北逐渐升高, 整体盐度梯度则刚好相反, 此结果与王婕妤[13]等人研究一致。所以夏季在调查海域相对低温高盐的东南部主要为瓜水母幼体、双刺唇角水蚤、精致真刺水蚤及中华哲水蚤, 高温低盐的西北部近岸海域主要为背针胸刺水蚤和小拟哲水蚤, 而高温高盐外海侧主要为百陶带箭虫、真刺唇角水蚤和平滑真刺水蚤等。夏季水温较高、适温种类可在整个调查海域生长繁殖以致于各站位物种相似度增加, 所以导致近岸区(B区)与外海区(A区)的空间差异性不显著, 而盐度则是导致河口区(C区)与外海区(A区)差异显著的主要原因。
秋季适宜广温种类和低温种类生存; 此时季风风向开始由西南风向转为东北风向, 暖流退至南侧, 使得该海域南侧温度高于北侧。而河口径流量减少, 河口冲淡水对海区的影响减小, 各站位盐度有所上升但梯度方向与夏季相同保持不变, 浮游动物分布主要受近岸上升流的作用影响, 背针胸刺水蚤, 百陶箭虫等优势种主要分布在舟山南侧近岸海区。冬季陆地河流处于枯水期, 舟山海域暖流沿岸退却, 外海侧主要受寒流影响, 内海侧受近岸上升流影响。加之季风南下, 海域温度降至全年最低, 浮游动物主要为耐低温或近岸低盐种类, 所以浮游动物的群落结构主要为外海侧与内海侧差异显著。
另外, Chl a和营养盐也是影响浮游动物群落的重要因素[33-35]。Chl a是评估海域初级生产力与浮游植物生物量的重要指标之一。营养盐是影响浮游植物的生长繁殖重要环境因子, 从而间接影响浮游动物。本次调查中, 舟山海域春季和夏季平均氮磷比分别为32.8:1和27.4:1, 且所有站位水质均处于氮富营养化水平。春季溶解无机氮输入主要来源于长江和钱江的径流输入, 有研究[36]表明陆地径流带来的溶解无机氮的量远大于溶解磷酸盐的量, 影响营养元素的平衡。夏季是浮游植物生长繁殖的高峰期, 大量消耗溶解无机氮, 所以夏季氮磷比低于春季, 溶解无机磷成为了限制浮游植物生长的主要影响因子, 有研究[29]表明舟山附近海域春季为富营养水平, 夏季主要为磷潜在限制性富营养。秋、冬季陆源和潮流带来的营养减少, 舟山海域而舟山东部海域靠外海, 水流交换比西部内海要好, 且内海侧处于背风向, 受东北季风相对较小, 海区环境更稳定, 因此人类活动对舟山西部海域的生物群落干扰就变得显著。据报道舟山海域海洋环境受污染严重[37-39], 所以在部分受污染的海区容易出现相对富营养状态, 形成小范围的聚集区, 从而导致冬季大多数站位浮游生物丰度太低而个别站位丰度高, 如金塘岛附近浮游动物丰度和生物量极高很可能是由于该处港口码头众多, 人类活动干扰所致。
4 结论(1) 四季共鉴定浮游动物88种和浮游幼体19类, 桡足类是四季的主要生物类群; 夏季的浮游动物种类(67种)、浮游幼体(15类)和水母类种类数(8种)均为四季最多; 四季间优势种更替明显, 春-夏、夏-秋、秋-冬和冬-春相邻季节的优势种更替率分别为75%、80%、100%和60%。
(2) 浮游动物平均生物量和平均丰度四季均为夏季最高, 分别为176.34 mg/m3和143.97个/m3。浮游动物的群落结构四季稳定性较好, 其中夏季最为稳定。
(3) 浮游动物季节变化明显, 温度、盐度、Chl a和溶解氧是影响浮游动物季节分化的重要因素。其中温度和溶解氧是四季分化的主要影响因素, 盐度和Chl a是夏季和秋季分化的主要影响因素。
(4) 四个季节的浮游生物群落均存在空间差异, 但春季的区域间差异最明显, 其次是冬季, 夏季仅外海区与河口区差异显著, 秋季仅舟山近岸区与河口区差异显著。温度和盐度梯度直接影响浮游动物的分布, 营养盐、悬浮物和Chl a则是通过浮游植物间接影响浮游动物的空间分布。春季浮游动物群落空间分布主要受盐度的影响, 夏季主要受温度、盐度和Chl a的影响, 秋季主要受Chl a的影响, 冬季主要受悬浮物和溶解氧的影响, 而营养盐对每个季节的浮游动物群落分布都有一定的影响。
| [1] |
徐兆礼, 王荣, 陈亚瞿. 黄海南部及东海中小型浮游桡足类生态学研究Ⅰ.数量分布. 水产学报, 2003, 27(S1): 1-8. |
| [2] |
陈亚瞿, 郑国兴, 朱启琴. 长江口区浮游动物初步研究. 东海海洋, 1985(3): 53-61. |
| [3] |
陈亚瞿, 徐兆礼, 王云龙, 胡方西, 韩明宝, 严宏昌. 长江口河口锋区浮游动物生态研究Ⅱ种类组成、群落结构、水系指示种. 中国水产科学, 1995, 2(1): 59-63. DOI:10.3321/j.issn:1005-8737.1995.01.004 |
| [4] |
金显仕, 单秀娟, 郭学武, 李显森. 长江口及其邻近海域渔业生物的群落结构特征. 生态学报, 2009, 29(09): 4761-4772. DOI:10.3321/j.issn:1000-0933.2009.09.021 |
| [5] |
吴鹏, 倪勇强, 孙毛明, 毛珊珊, 张广之, 汝少国. 舟山钓梁围垦一期工程对附近海域生态系统影响研究. 海洋通报, 2013, 32(5): 540-548552. |
| [6] |
俞存根, 张平, 郭朋军, 邓小艳, 郑基, 徐娜娜. 围填海区渔业生态损害的补偿标准定量研究——以舟山近岸海域为例. 生态学报, 2019, 39(4): 1416-1425. |
| [7] |
徐娜娜, 邱颖, 姚炎明, 朱明栋, 黄大吉. 2002-2011年舟山近岸海域生态环境变化. 安徽农业科学, 2015, 43(3): 292-296. DOI:10.3969/j.issn.0517-6611.2015.03.098 |
| [8] |
施辰阳, 白炜炜, 陈李庆, 陈柏峰, 李伟东, 杨华云. 舟山海洋渔业资源现状分析及其可持续发展对策探讨. 环境科学导刊, 2017, 36(6): 11-16. |
| [9] |
金敬林, 方添坤, 朱四喜. 2011年夏季舟山近岸海域浮游动物调查研究. 环保科技, 2015, 21(4): 11-15. DOI:10.3969/j.issn.1674-0254.2015.04.003 |
| [10] |
骆鑫, 曾江宁, 徐晓群, 杜萍, 廖一波, 刘晶晶. 舟山海域夏、秋季浮游动物的分布特征及其与环境因子的关系. 生态学报, 2016, 36(24): 8194-8204. |
| [11] |
何舟洋, 许永久, 俞存根, 谢旭, 张平, 朱明星, 何江楠, 张亚洲. 舟山近岸海域秋季浮游动物多样性与环境因子的关系. 浙江海洋学院学报:自然科学版, 2017, 36(2): 144-150. DOI:10.3969/j.issn.1008-830X.2017.02.008 |
| [12] |
章菁, 杨关铭, 王春生, 张东声. 舟山群岛邻近海域浮游动物生态研究Ⅰ.种类组成与数量分布. 海洋学研究, 2008, 26(4): 20-28. DOI:10.3969/j.issn.1001-909X.2008.04.003 |
| [13] |
王婕妤, 黄备, 贾海波, 魏娜. 2011年春、秋季浙江舟山海域浮游动物群落结构和多样性现状. 浙江海洋学院学报:自然科学版, 2014, 33(4): 311-316. DOI:10.3969/j.issn.1008-830X.2014.04.004 |
| [14] |
孟庆超, 赵淑江. 舟山东极岛海域浮游动物群落结构特征. 农村经济与科技, 2019, 30(11): 63-66. |
| [15] |
俞存根, 陈小庆, 胡颢琰, 黄备, 郑基, 覃涛, 宁平. 舟山渔场及邻近海域浮游动物种类组成及群落结构特征. 水生生物学报, 2011, 35(1): 183-193. |
| [16] |
国家标准化管理委员会.GB/T12763.6-2007海洋调查规范第6部分: 海洋生物调查.北京: 中国标准出版社, 2007.
|
| [17] |
国家标准化管理委员会.GB/T12763.4-2007海洋调查规范第4部分: 海水化学要素调查.北京: 中国标准出版社, 2008.
|
| [18] |
徐兆礼, 沈新强, 马胜伟. 春、夏季长江口邻近水域浮游动物优势种的生态特征. 海洋科学, 2005, 29(12): 13-19. DOI:10.3969/j.issn.1000-3096.2005.12.004 |
| [19] |
刘镇盛, 王春生, 张志南, 刘诚刚, 杨关铭. 三门湾浮游动物的季节变动及微型浮游动物摄食影响. 生态学报, 2006(12): 3931-3941. DOI:10.3321/j.issn:1000-0933.2006.12.003 |
| [20] |
沈国英, 施并章. 海洋生态学(第二版). 北京: 科学出版社, 2002: 37-39.
|
| [21] |
李振华, 周永东, 张亚洲, 朱增军, 徐开达. 2014年春季舟山海域浮游植物群落特征. 浙江海洋大学学报:自然科学版, 2018, 37(5): 405-411. DOI:10.3969/j.issn.1008-830X.2018.05.005 |
| [22] |
蒋巧丽, 许永久, 俞存根, 张浩良, 肖祎, 唐逸铮, 张亚洲. 2016年夏季长江口邻近海域浮游动物群落结构. 应用生态学报, 2018, 29(9): 3078-3084. |
| [23] |
Zhou F, Xue H J, Huang D J, Xuan J L, Ni X B, Xiu P, Hao Q. Cross-shelf exchange in the shelf of the East China Sea. Journal of Geophysical Research:Oceans, 2015, 120(3): 1545-1572. DOI:10.1002/2014JC010567 |
| [24] |
Gao X L, Song J M, Li X G. Zooplankton spatial and diurnal variations in the Changjiang River estuary before operation of the Three Gorges Dam. Chinese Journal of Oceanology and Limnology, 2011, 29(3): 591-602. DOI:10.1007/s00343-011-0098-3 |
| [25] |
洪旭光, 张锡烈, 俞建銮, 孟凡. 东海北部黑潮区浮游动物的多样性研究. 海洋学报, 2001, 23(1): 139-142. DOI:10.3321/j.issn:0253-4193.2001.01.019 |
| [26] |
Umezawa Y, Yamaguchi A, Ishizaka J, Hasegawa T, Yoshimizu C, Tayasu I, Yoshimura H, Morii Y, Aoshima T, Yamawaki N. Seasonal shifts in the contributions of the Changjiang River and the Kuroshio Current to nitrate dynamics in the continental shelf of the norther East China Sea based on a nitrate dual isotopic composition approach. Biogeosciences, 2014, 11(4): 1297-1317. DOI:10.5194/bg-11-1297-2014 |
| [27] |
Telesh I V, Khlebovich V V. Principal processes within the estuarine salinity gradient:A review. Marine Pollution Bulletin, 2010, 61(4/6): 149-155. |
| [28] |
Zakaria H Y, Radwan A A, Said M A. Influence of salinity variations on zooplankton community in El-mex Bay, Alexandria, Egypt. Egyptian Journal of Aquatic Research, 2007, 33(3): 52-67. |
| [29] |
李磊, 夏培艳, 唐峰华, 沈新强. 舟山附近海域富营养化的时空分布及其与环境因子的关系. 生态学杂志, 2011, 30(4): 771-777. |
| [30] |
徐晓群, 曾江宁, 陈全震, 刘晶晶, 杜萍, 王桂忠. 乐清湾海域浮游动物群落分布的季节变化特征及其环境影响因子. 海洋学研究, 2012, 30(1): 34-40. DOI:10.3969/j.issn.1001-909X.2012.01.005 |
| [31] |
杜明敏, 刘镇盛, 王春生, 张东声, 章菁. 中国近海浮游动物群落结构及季节变化. 生态学报, 2013, 33(17): 5407-5418. |
| [32] |
Shi Y Q, Zuo T, Yuan W, Sun J Q, Wang J. Spatial variation in zooplankton communities in relation to key environmental factors in the Yellow Sea and East China Sea during winter. Continental Shelf Research, 2018, 170: 33-41. DOI:10.1016/j.csr.2018.10.004 |
| [33] |
章飞燕, 唐静亮, 李道季, 方涛, 王彪. 夏、秋季长江口及毗邻海域浮游动物的分布与变化. 水生生物学报, 2009, 33(6): 1219-1225. |
| [34] |
Zhou S C, Jin B S, Guo L, Qin H M, Chu T J, Wu J H. Spatial distribution of zooplankton in the intertidal marsh creeks of the Yangtze River Estuary, China. Estuarine, Coastal and Shelf Science, 2009, 85(3): 399-406. DOI:10.1016/j.ecss.2009.09.002 |
| [35] |
Zhang S Y, Zhou Q H, Xu D, Lin J D, Cheng S P, Wu Z B. Effects of sediment dredging on water quality and zooplankton community structure in a shallow of eutrophic lake. Journal of environmental sciences, 2010, 22(2): 218-224. DOI:10.1016/S1001-0742(09)60096-6 |
| [36] |
王保栋. 长江冲淡水的扩展及其营养盐的输运. 黄渤海海洋学报, 1998, 16(2): 42-48. |
| [37] |
余静, 孙英兰, 张越美, 张燕, 蔡惠文. 宁波-舟山海域入海污染物环境容量研究. 环境污染与防治, 2006, 28(1): 21-24. DOI:10.3969/j.issn.1001-3865.2006.01.007 |
| [38] |
刘琴, 朱婧睿, 钟志. 舟山朱家尖附近海域海洋渔业环境质量评价(英文). 农业科学与技术, 2016, 17(11): 2513-2515. |
| [39] |
李铁军, 郭远明, 薛彬, 李佩佩, 朱敬萍, 金衍健. 舟山电厂运行对附近海域环境影响分析. 广州化工, 2013, 41(1): 121-123, 141-141. DOI:10.3969/j.issn.1001-9677.2013.01.045 |