 2020, Vol. 40
2020, Vol. 40文章信息
- 张倩, 杨晶, 姚宝辉, 蔡志远, 孙小妹, 王缠, 郭怀亮, 谭宇尘, 苏军虎
- ZHANG Qian, YANG Jing, YAO Baohui, CAI Zhiyuan, SUN Xiaomei, WANG Chan, GUO Huailiang, TAN Yuchen, SU Junhu
- 放牧管理模式对高寒草甸鼢鼠鼠丘群落演替的影响
- Effects of different grazing strategies on zokor mounds plant community succession in alpine meadow
- 生态学报. 2020, 40(8): 2802-2811
- Acta Ecologica Sinica. 2020, 40(8): 2802-2811
- http://dx.doi.org/10.5846/stxb201902120249
-
文章历史
- 收稿日期: 2019-02-12
- 网络出版日期: 2019-12-31





2. 甘肃农业大学-新西兰梅西大学草地生物多样性研究中心, 兰州 730070;
3. 甘肃农业大学资源与环境学院, 兰州 730070
2. Gansu Agricultural University-Massey University Research Centre for Grassland Biodiversity, Gansu Agricultural University, Lanzhou 730070, China;
3. College of Resource and Environmental Science, Gansu Agricultural University, Lanzhou 730070, China
放牧是草地最基本且尤为重要的利用方式之一, 放牧制度作为放牧管理中的利用体系, 从时间和空间角度出发, 规定牛、羊等对草地的使用强度和频率, 保持植物生长和家畜数量之间达到平衡[1]。合理有效的放牧制度对增加草地物种多样性具有重要意义, 也能促进草地健康可持续发展[2]。青藏高原草地退化日益加剧, 过度放牧是导致草地退化的重要原因之一[3]。而家畜主要通过采食、践踏和排泄粪便影响草地植物群落, 以及对物种竞争等产生不同程度的干扰[4]。放牧优化假说认为, 当放牧强度处于中等程度时, 草地初级生产力最大, 即通过优化的放牧制度能够增加草地初级生产力[5], 现有的放牧制度如划区轮牧是放牧强度在空间和时间上调整, 而生长季休牧的放牧制度是放牧强度在时间上的调整, 而连续放牧的管理制度并没有考虑放牧强度的时间和空间效应[6]。不同的放牧制度对群落的优势种、植被特征和草地生产力等产生不同的影响[7], 禁牧有利于调节食草动物与植物的关系, 也是草地管理最常用的手段, 可以在短期内恢复草地建群种的盖度及促进草地生态系统良性循环[8], 但长期禁牧会导致草地生产力降低等[9], 不同的放牧制度, 如划区轮牧、生长季休牧和连续放牧等对草地的影响不同, 客观准确评价放牧制度对生态、生产之间的权衡, 具有重要的现实指导意义。
高寒草甸放牧利用下高原鼢鼠(Eospalax baileyi)等危害的发生是草地管理的关键难题之一, 高原鼢鼠是青藏高原高寒草地优势害鼠之一, 主要采食地表以下3—20 cm的植物根、茎[10]。高原鼢鼠的挖掘、造丘活动是草地退化的重要驱动因子之一, 其所形成的鼠丘是影响草地的重要方式之一, 主要表现在对草地原有植被的覆盖、降低群落中物种多样性和改变植被空间结构, 同时也影响草地植物生产力和地上植被组成等[11]。Koide等[12]研究表明, 鼢鼠扰动会降低优势禾草的地上和地下生物量。不同放牧制度下, 家畜的践踏、排泄物及摄食方式均会对啮齿动物的种群数量产生不同影响[13]。中度干扰假说认为, 中等程度的干扰, 一方面会加速竞争力较弱的物种获得资源, 提高草地物种多样性[14];另一方面会导致时间和空间的异质性, 有利于生态位分化[15]。研究发现, 休牧和轮牧下显著降低禾本科和杂类草的生物量和丰富度, 而高原鼢鼠干扰对草地植物的物种丰富度、Shannon-wiener指数和Simpson指数等均无显著影响[16], 但轻度或中度高原鼢鼠干扰能够提高群落物种多样性, 以及物种多样性指数(Simpson指数和Shannon-wiener指数)随鼠丘群落演替的增加而增加[17-18]。不同放牧制度对高原鼢鼠鼠丘的影响不同, 不同的放牧制度也会导致不同的生态机遇, 给鼠丘植被演替增加机会, 可能会有不同的演替情况, 以及使群落中物种间的竞争发生变化。而鼠丘植被的演替变化, 直接影响草地的利用以及高原鼢鼠危害的发生情况等, 从放牧制度对鼠丘植被演替影响的认知中能获得更有效的草地管理模式[19]。然而迄今未见该方面的报道, 在草地持续退化的态势下, 从鼠丘植被特征以及优化放牧管理策略的角度入手, 对退化草地的综合治理和有效管理具有重要意义。基于此, 本研究通过放牧管理模式对高原鼢鼠鼠丘群落结构影响的研究, 探讨放牧和高原鼢鼠扰动下鼠丘植被生物量(地上和地下)、物种多样性和群落演替, 旨在为高原鼢鼠放牧管理, 以及高寒草甸资源的合理利用和退化草地的恢复与重建提供理论依据。
1 研究区域与方法 1.1 研究区域概况研究区位于甘肃省天祝县抓喜秀龙镇甘肃农业大学天祝高山草原试验站(37°11′24″—37°12′ N, 102°45′—102°46′48″ E), 平均海拔在2900—3100 m, 年均气温1℃, 年均日照时数2500—2700 h, 降水主要集中在6—8月, 年均降雨量400 mm, 年均蒸发量1600 mm, 无绝对无霜期, 亚高山土壤类型[20]。草地类型为高寒草甸, 主要优势种为冰草(Agropyron cristatum)、苔草(Carex supina)、黄花棘豆(Medicago falcata)、二裂萎菱菜(Potentilla bifurca)、鹅绒萎菱菜(Potentilla anserina)、密花香薷(Elsholtzia densa)等杂类草为主。
1.2 样地设置2018年8月初在试验站附近选择划区轮牧(Rotational grazing, RG, 面积0.95 hm2, 放牧时间从2012年到2018年, 轮牧时间为180 d, 轮牧次数为3次, 轮牧强度为5.38羊hm2/a, 轮牧起始时间为5月15日, 结束时间为11月15日)、连续放牧(Continuous grazing, CG, 面积1.44 hm2, 全年连续放牧, 放牧率为8.37羊hm2/a)、生长季休牧(Growing season grazing, GSG, 面积1.47hm2, 时间为2015年到2018年, 生长季放牧率为0, 非生长季放牧率为4.53羊hm2/a, 每年在4月15日到9月15日间休牧, 其余时间自由放牧)和禁牧(Prohibition grazing, PG, 面积5.2 hm2, 时间为2011年到2018年, 全年禁牧), 每个草地利用模式样地仅设置1个样地。4种放牧制度样地, 在每个样地中选择高原鼢鼠鼠丘样地以及在鼢鼠鼠丘周围未干扰的草地中选定对照(CM)植物群落, 以空间序列代替演替时间序列的方法[17], 研究不同放牧制度对不同演替阶段高原鼢鼠土丘植物群落结构的影响。结合土壤性状、植被盖度及物种组成等判断鼠丘年限, 具体为一年鼠丘(ZM1), 即1次越冬, 土壤较疏松, 植被总盖度大于20%, 一年植物占90%以上;两年鼠丘(ZM2), 即2次越冬, 土壤较紧实, 植被总盖度大于60%, 多年禾本科植物小于15%;三年鼠丘(ZM3), 即3次越冬, 土壤紧实, 植被总盖度大于75%, 一年植物小于10%;多年鼠丘(ZMM), 即4次以上越冬, 土壤紧实, 植被总盖度大于85%, CM样地, 土壤紧实, 植被总盖度大于90%[21]。
1.3 取样及计算方法 1.3.1 群落特征和生物量取样分别在每个样地中选择12个鼠丘(每个演替阶段选择3个, 鼠丘可近似看成椭圆形, 各年限鼠丘的长轴×短轴均在45 cm×45 cm — 65 cm×68 cm范围之内)样方(50 cm×50 cm), 以及在鼢鼠鼠丘周围未干扰的草地(距离鼠丘至少50 cm以外)中随机选取3个50 cm×50 cm的自然植被样方作为对照(CM), 用针刺法测定物种总盖度;用卷尺测量物种的高度;用样圆测定物种频度, 样圆面积0.25 m2。用最大值标准化法计算物种高度、盖度和频度的相对值。在每个50 cm×50 cm样方内, 齐地面刈割样方内所有植物放入信封。在烘箱中恒温70℃烘干48 h至恒量后, 称重记录。地下生物量测定采用20 cm×20 cm的铁丝框, 在地上部分刈割完的样方内, 分0—10 cm、10—20 cm和20—30 cm层取出地下土壤样品装入60目的网袋, 抖落松散土壤并用清水冲洗干净, 在70℃的烘箱内烘干48 h至恒量, 并称重记录, 生物量都以g/m2表示。
1.3.2 物种重要值、丰富度指数、Pielou均匀度指数和Shannon-wiener指数计算方法
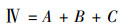
|
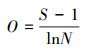
|
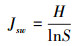
|
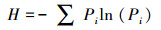
|
式中, IV为物种的重要值, A为相对高度, B为相对盖度, C为相对频度;O为物种丰富度指数, S为样方中物种总数, N代表样方各物种重要值之和;Pi为物种i的相对重要值, Jsw为物种Pielou均匀度指数;H为物种Shannon-wiener指数[22]。
1.4 数据分析应用Excel 2007对所得数据进行简单记录和处理并制作相关图表;采用SPSS 19.0软件对同一放牧方式下不同年限鼠丘的群落特征进行单因素方差分析。
对不同放牧制度下群落演替的驱动因子进行主成分分析, 主要步骤为:(1)将选取的指标变量建立原始数据矩阵;(2)原始数据矩阵的标准化;(3)依据特征值≥ 1的原则提取公因子, 确定公因子数, 并运用Varimax旋转法对初始因子进行旋转, 以便给予明确的因子解释[23]。
2 结果与分析 2.1 两种因素影响下群落特征变化在交互作用下, 群落的盖度、高度、重要值、丰富度指数O、Shannon-wiener指数H、地上生物量均有显著影响(P < 0.05), Pielou均匀度指数Jsw和地下生物量交互作用不显著(P>0.05)(表 1)。
| 指标 Index |
因素A(年限鼠丘) Factor A (different zokor mounds) |
因素B(放牧方式) Factor B (grazing method) |
交互作用(A/B) Interaction (A/B) |
||||||||
| df | F | P | df | F | P | df | F | P | |||
| 盖度Coverage | 4 | 84.678 | 0.000 | 3 | 14.665 | 0.000 | 12 | 2.863 | 0.006 | ||
| 高度Height | 4 | 4.310 | 0.005 | 3 | 40.665 | 0.000 | 12 | 3.073 | 0.004 | ||
| 重要值Important value | 4 | 5.342 | 0.002 | 3 | 2.995 | 0.042 | 12 | 2.163 | 0.034 | ||
| 丰富度指数Richness index | 4 | 0.652 | 0.629 | 3 | 2.963 | 0.043 | 12 | 3.269 | 0.002 | ||
| 均匀度指数Uniformity index | 4 | 1.152 | 0.346 | 3 | 2.663 | 0.061 | 12 | 1.493 | 0.167 | ||
| Shannon-wiener指数 | 4 | 1.327 | 0.277 | 3 | 4.416 | 0.009 | 12 | 4.519 | 0.000 | ||
| 地上生物量AGB | 4 | 86.400 | 0.000 | 3 | 21.169 | 0.000 | 12 | 3.181 | 0.003 | ||
| 地下生物量UGB | 4 | 2.420 | 0.064 | 3 | 1.842 | 0.155 | 12 | 0.128 | 1.000 | ||
| AGB:地上生物量, Above ground biomass; UGB:地下生物量, Under ground biomass | |||||||||||
同一放牧管理模式下, 分析不同年限鼠丘植被群落总盖度、重要值、丰富度、均匀度和优势种等群落特征的变化, 结果发现:GSG和PG下鼠丘物种平均高度均高于CG和RG;RG和PG下ZM3和ZMM物种重要值均高于CM, 但差异不显著(P>0.05), CG和GSG下CM物种重要值均最高, RG和PG下ZM3物种重要值最高, RG和PG下CM与所有年限鼠丘物种重要值之间无显著性差异(P>0.05);RG和PG下ZM3物种Jsw高于CM, CG和GSG下CM物种Jsw最高, PG、CG和GSG下所有年限鼠丘物种Jsw与CM物种Jsw之间均无显著性差异(P>0.05), RG下CM物种Jsw显著低于ZM1、ZM2和ZMM物种Jsw(P < 0.05);RG下ZM1物种O显著高于CM物种O(P < 0.05), 且ZM3物种O显著高于ZM1和ZM2物种O(P < 0.05), PG和GSG下ZM2和ZM3物种O均高于CM物种O, 但差异不显著(P>0.05), CG下CM物种O显著高于ZM1物种O(P < 0.05)(表 2)。
| 样地 Plots |
鼠丘年限 Zokor mounds |
总盖度/% Total coverage |
平均高度 Average height |
重要值 Important value |
均匀度指数 Uniformity index |
丰富度指数 Richness index |
优势种 Dominant species |
| RG | ZM1 | 40.00±7.00c | 3.46±0.71ab | 0.96±0.05a | 0.92±0.01a | 3.56±0.05a | 冰草Agropyron cristatum 苔草Carex supina 二裂萎菱菜Potentilla bifurca |
| ZM2 | 55.67±6.39bc | 2.10±0.22b | 1.07±0.03a | 0.92±0.00a | 3.30±0.05ab | 冰草A. cristatum 兰石草Lancea tibetica 黄花棘豆Oxytropis ochrocephala |
|
| ZM3 | 63.00±5.86b | 2.60±0.29b | 1.21±0.04a | 0.90±0.02ab | 2.42±0.09c | 黄花棘豆O. ochrocephala 苔草C. supina 二裂萎菱菜P. bifurca |
|
| ZMM | 89.67±1.20a | 3.49±0.24ab | 1.15±0.16a | 0.91±0.00a | 3.11±0.35abc | 冰草A. cristatum 苔草C. supina 黄花棘豆O. ochrocephala |
|
| CM | 99.00±0.58a | 4.61±0.41a | 1.06±0.11a | 0.85±0.02b | 2.72±0.36bc | 苔草C. supina 冰草A. cristatum 黄花棘豆O. ochrocephala |
|
| PG | ZM1 | 46.33±3.84c | 9.07±3.45b | 1.38±0.23a | 0.90±0.03a | 2.12±0.19a | 冰草A. cristatum 黄花棘豆O. ochrocephala 紫菀Aster tataricus |
| ZM2 | 76.33±11.68b | 11.78±0.79b | 0.98±0.04a | 0.88±0.03a | 3.10±0.44a | 冰草A. cristatum 密花香薷Elsholtzia densa 节裂角茴香Hypecoum leptocarpum |
|
| ZM3 | 89.00±3.00ab | 10.84±1.04b | 1.47±0.19a | 0.95±0.02a | 2.67±0.65a | 紫菀A. tataricus 苔草C. supina 二裂萎菱菜P. bifurca |
|
| ZMM | 100.00±0.00a | 22.37±3.94a | 1.40±0.17a | 0.91±0.06a | 2.57±0.23a | 冰草A. cristatum 苔草C. supina 臭蒿Artemisia hedinii |
|
| CM | 100.00±0.00a | 14.33±1.63b | 1.31±0.13a | 0.94±0.02a | 2.64±0.20a | 冰草A. cristatum 苔草C. supina 黄花棘豆O. ochrocephala |
|
| CG | ZM1 | 33.33±6.17d | 9.00±1.70a | 1.04±0.13b | 0.83±0.09a | 2.27±0.14b | 冰草A. cristatum 二裂萎菱菜P. bifurca 紫菀A. tataricus |
| ZM2 | 53.67±1.20c | 7.04±0.76a | 1.12±0.11b | 0.86±0.04a | 2.55±0.40ab | 冰草A. cristatum 兰石草L. tibetica 多裂萎菱菜P. anserina |
|
| ZM3 | 85.00±4.04b | 6.69±0.35a | 1.10±0.12b | 0.92±0.02a | 2.58±0.24ab | 冰草A. cristatum 二裂萎菱菜P. bifurca 黄花棘豆O. ochrocephala |
|
| ZMM | 100.00±0.00a | 8.73±0.12a | 1.56±0.07a | 0.96±0.00a | 3.52±0.29a | 冰草A. cristatum 鹅绒萎菱菜P. anserina 苔草C. supina |
|
| CM | 100.00±0.00a | 8.73±0.12a | 1.56±0.07a | 0.96±0.00a | 3.52±0.29a | 冰草A. cristatum 苔草C. supina 多裂萎菱菜P. mulatifida |
|
| GSG | ZM1 | 43.67±22.51b | 11.8±1.38a | 0.65±0.33b | 0.64±0.32a | 2.46±1.26a | 黄花棘豆O. ochrocephala 冰草A. cristatum 密花香薷E. densa |
| ZM2 | 82.67±9.13a | 8.67±0.96a | 1.03±0.08ab | 0.93±0.01a | 3.50±0.59a | 冰草A. cristatum 黄花棘豆O. ochrocephala 二裂萎菱菜P. bifurca |
|
| ZM3 | 93.33±3.67a | 9.70±2.00a | 1.09±0.04ab | 0.95±0.00a | 3.77±0.30a | 冰草A. cristatum 黄花棘豆O. ochrocephala 兰石草L.tibetica |
|
| ZMM | 99.33±0.67a | 9.14±0.80a | 1.48±0.04a | 0.90±0.01a | 2.28±0.19a | 冰草A. cristatum 黄花棘豆O. ochrocephala 苔草C. supina |
|
| CM | 100.00±0.00a | 11.43±0.39a | 1.52±0.03a | 0.95±0.01a | 2.62±0.17a | 苔草C. supina 冰草A. cristatum 黄花棘豆O. ochrocephala |
|
| RG:划区轮牧, Rotational grazing; PG:禁牧, Grazing prohibition; CG:连续放牧, Continuous grazing; GSG:生长季休牧, Growing season grazing; ZM1:一年鼠丘, One year zokor mounds; ZM2:二年鼠丘, Two years zokor mounds; ZM3:三年鼠丘, Three years zokor mounds; ZMM:多年鼠丘, Multi-year zokor mounds; CM:对照, Control meadow; 不同字母表示同一放牧方式下, 不同年限鼠丘之间的差异性(P < 0.05) | |||||||
以α多样性(Shannon-wiener指数H)为物种多样性指标, 分析不同放牧管理模式对不同年限鼠丘物种多样性的影响, 发现CG下物种H最低, GSG下H较高;RG下ZM1和ZM2物种H显著高于CM物种H(P < 0.05);GSG下ZM1和ZM3物种H显著高于CM物种H(P < 0.05);PG下ZM3物种H高于CM物种H, 但差异不显著(P>0.05);CG下CM物种H显著高于ZM1、ZM2、ZM3和CM物种H(P < 0.05)(图 1)。

|
| 图 1 不同放牧管理模式在不同鼠丘年限下Shannon-wiener指数的比较 Fig. 1 Comparison of Shannon-Weiner index in different grazing management modes under different zokor mounds year RG:划区轮牧, Rotational grazing; PG:禁牧, Grazing prohibition; CG:连续放牧, Continuous grazing; GSG:生长季休牧, Growing season grazing; ZM1:一年鼠丘, One year zokor mounds; ZM2:二年鼠丘, Two years zokor mounds; ZM3:三年鼠丘, Three years zokor mounds; ZMM:多年鼠丘, Multi-year zokor mounds; CM:对照, Control meadow; 不同字母表示同一放牧方式下, 不同年限鼠丘之间的差异性(P < 0.05) |
PG、CG和GSG下, 随着鼠丘年限的增加地上生物量也增多, 但CG和GSG下CM地上生物量最高, PG下ZMM地上生物量最高, RG下地上生物量大小为:ZM1 < ZM3 < ZM2 < ZMM < CM, CG和RG下各年限鼠丘的地上生物量均高于PG和GSG下各鼠丘的生物量;所有样地的地下生物量在10 cm土层分布最多, 随着土层深度增加地下生物量减少, 对照样地中地下总生物量最高, 鼢鼠的扰动减少了地下生物量, 但各放牧管理模式下大小为:GSG>PG>CG>RG。GSG下CM地下总生物量接近3500 g/m2, 而PG、RG和GSG下ZM2地下总生物量均为最小, CG下ZM1地下总生物量最小(图 2)。

|
| 图 2 不同放牧管理模式在不同鼠丘年限下生物量分配 Fig. 2 Biodiversity allocation in different grazing management modes under different zokor mounds year RG:划区轮牧, Rotational grazing; PG:禁牧, Grazing prohibition; CG:连续放牧, Continuous grazing; GSG:生长季休牧, Growing season grazing; ZM1:一年鼠丘, One year zokor mounds; ZM2:二年鼠丘, Two years zokor mounds; ZM3:三年鼠丘, Three years zokor mounds; ZMM:多年鼠丘, Multi-year zokor mounds; CM:对照, Control meadow; AGB:地上生物量, Above ground biomass |
主成分分析结果表明, RG下2个公因子对样本方差的贡献率为74.91%, 即2个公因子包含了原始数据信息总量的74.91%, 可以用这2个公因子代表原来的8个指标, 第一主成分上, 具有高载荷因子的指标有地上生物量(0.939)、盖度(0.889), 第二主成分上, 高载荷因子的指标有O(0.874)、重要值(-0.829), 各指标在主成分中的载荷量, 代表与主成分关系的密切程度, 值越大则与主成分关系越密切, 越能代表鼠丘群落演替的变化;PG下3个公因子对样本方差的贡献率为77.23%, 第一主成分上, 具有高载荷因子的指标有地上生物量(0.934)、盖度(0.915);第二主成分上, 高载荷因子有O(0.914)、H(0.906), 第三主成分上, 高载荷因子有Jsw(0.889);CG下2个公因子对样本方差的贡献率为75.71%, 第一主成分上, 具有高载荷因子的指标有O(0.925)、H(0.871)、盖度(0.812);第二主成分上, 高载荷因子有高度(0.858)、重要值(0.860);GSG下3个公因子对样本方差的贡献率为85.99%, 第一主成分上, 具有高载荷因子的指标有H(0.974)、O(0.849), 第二主成分上, 高载荷因子有盖度(0.914)、地上生物量(0.901), 第三主成分上, 高载荷因子有高度(0.890)(表 3)。
| 指标 Index |
RG | PG | CG | GSG | |||||||||
| PC1 | PC2 | PC1 | PC2 | PC3 | PC1 | PC2 | PC1 | PC2 | PC3 | ||||
| 地上生物量AGB | 0.939 | 0.036 | 0.934 | 0.071 | 0.081 | 0.830 | 0.391 | -0.303 | 0.901 | 0.083 | |||
| 丰富度指数Richness index | 0.344 | 0.874 | -0.032 | 0.914 | -0.376 | 0.925 | -0.152 | 0.849 | -0.420 | -0.207 | |||
| 盖度Coverage | 0.889 | -0.090 | 0.915 | 0.135 | 0.002 | 0.775 | 0.354 | -0.013 | 0.914 | -0.217 | |||
| Shannon-wiener指数 | -0.520 | 0.753 | 0.262 | 0.906 | 0.240 | 0.871 | 0.440 | 0.974 | -0.166 | -0.047 | |||
| 均匀度指数Uniformity index | -0.792 | 0.355 | 0.040 | 0.138 | 0.889 | 0.390 | 0.761 | 0.800 | -0.171 | 0.418 | |||
| 高度Height | 0.613 | -0.055 | 0.774 | 0.062 | 0.263 | -0.148 | 0.858 | 0.208 | -0.052 | 0.890 | |||
| 重要值Important value | 0.001 | -0.829 | 0.294 | -0.384 | 0.716 | 0.305 | 0.860 | -0.455 | 0.837 | 0.238 | |||
| 地下生物量UGB | 0.513 | 0.714 | 0.379 | -0.208 | 0.313 | 0.579 | 0.029 | -0.415 | 0.107 | 0.580 | |||
| 特征值Eigenvalues | 3.328 | 2.665 | 2.611 | 1.893 | 1.675 | 3.507 | 2.550 | 2.825 | 2.594 | 1.459 | |||
| 方差贡献率VCR/% | 41.596 | 33.306 | 32.633 | 23.660 | 20.938 | 43.840 | 31.872 | 35.319 | 32.429 | 18.239 | |||
| 累计贡献率CCR/% | 41.596 | 74.906 | 32.633 | 56.293 | 77.232 | 43.840 | 75.712 | 35.319 | 67.748 | 85.987 | |||
| RG:划区轮牧, Rotational grazing; PG:禁牧, Grazing prohibition; CG:连续放牧, Continuous grazing; GSG:生长季休牧, Growing season grazing; PC1:第一主成分, First principal component; PC2:第二主成分, Second principal component; PC3:第三主成分, Third principal component; AGB:地上生物量, Above ground biomass; UGB:地下生物量, Under ground biomass; VCR:方差贡献率, Variance contribution rate; CCR:累计贡献率, Cumulative contribution rate | |||||||||||||
由于生物和非生物因素的综合作用, 造成草地生态系统破坏愈加严重, 而不合理的放牧制度是造成这一现象的主要原因, 其不仅降低了草地生态系统的生产力和恢复力, 也降低畜产品产量, 从而影响当地牧民的基本生存[24]。合理有效的放牧方式, 能促进草地生态系统的结构和功能向有利方向发展, 尤其对群落的基本特征(盖度、高度和丰富度等)和群落多样性等会造成直接影响[25]。中度干扰假说认为, 中等强度的干扰会阻碍生长旺盛植物对资源的利用, 有利于生长能力较弱的物种对资源的吸收利用, 使群落物种多样性增高[26]。有研究发现, 中等程度的干扰下物种丰富度指数和Shannon-wiener指数高于禁牧区[27], 长期禁牧会增加家畜喜食植物的竞争能力, 阻碍群落中某些植物幼苗的生长, 导致群落不能充分进行物种更新, 进而降低群落物种多样性[28]。有研究表明, 禁牧和全生长季休牧能增加植物密度、生物量以及改善草地植被组成[1]。本研究中, GSG、PG和RG下地下总生物量都高于CG(表 2), 符合中度干扰假说。原因可能由于家畜啃食和践踏的强度较大, 使植物自身不能进行自我调节导致浅根系植物直接死亡, 群落结构单一, 从而导致地下生物量降低。
3.2 啮齿动物对草地群落特征和生物量的影响啮齿动物在草地生态系统中扮演着重要的角色[29]。啮食优良牧草, 挖掘活动损失牧草, 挖洞推土影响土壤肥力, 降低植被盖度, 促使土壤水分蒸发, 改变植被成分, 引起群落演替等, 同时也影响到高寒草地植物的生产力和草地地上植被组成、群落结构的复杂性等[30]。研究发现, 物种丰富度随高原鼢鼠新土丘数的增加呈先增加后降低的趋势, 也会随鼢鼠种群密度增加呈降低趋势, 出现这种差异的原因可能与其采用空间序列代替时间序列的方法有关[31]。有研究表明, 高原鼢鼠种群增长的过度放牧样地, 可以通过适度禁牧等休牧措施来提高草地群落多样性和生产力[32], 以及群落盖度随鼠丘密度的增加显著降低[18], 且植物高度随鼠丘年限的增加越来越大[33], 这些与本研究结果相似(图 1、图 2)。原因可能由于过度放牧会使草地退化越加严重, 进而使草地高原鼢鼠种群数量增加, 而适度放牧会改善草地退化, 高原鼢鼠种群数量也会降低, 以及随鼠丘年限增加, 鼠丘上土壤水分、养分等趋于稳定并逐渐增大, 对植物的生长提供良好的条件。
3.3 放牧管理模式对鼠丘群落演替的影响青藏高原高寒草甸对放牧、鼢鼠干扰等因素十分敏感, 微小波动都会使草地生态系统产生强烈的响应, 进而导致群落特征、植物多样性和生产力都发生很大的改变[34]。生态位理论认为物种能够共存是由于物种在生态位中的位置不同、生态位重叠较小和生态位分化等[35]。中性理论认为扩散限制是决定群落结构的重要因素, 且也受到空间尺度、环境因子、物种互利和密度等因子的影响[36]。Turnbull等[37]资源生态位模型研究表明, 生态位差异在物种共存时会在物种间产生互补效应, 但其不利于物种的稳定共存。有研究表明, 禁牧和自由放牧下物种重要值、多样性指数和优势度指数均小于季节性放牧地[19], 与本研究结果相同(图 1、表 2), 说明放牧强度对鼠丘群落的恢复有一定的影响, 由于高强度放牧下, 家畜长时间采食和践踏导致植物的根系受到一定的影响。有研究表明, 禁牧和划区轮牧对快速生长的一年生植物有利, 对多年生植物可能会产生一定的限制作用[38], 这很好的解释了本研究中RG和PG下ZM3物种重要值最高的原因。也有研究发现, 合理的放牧制度能够减弱优势种的竞争力, 加速弱势物种的存活, 提高草地物种多样性[5]。
不同放牧制度下, 植物的生活型和繁殖方式也不同, 资源比率/异质性假说认为, 异质性环境中两种或多种限制性资源比率与单一限制性资源相比, 其生态位分化更丰富[39]。放牧制度的差异性决定不同的物种组成和更替, 是构成群落演替的基础[40]。物种丰富度指数指群落内植物种类的多少, Shannon-wiener指数是反映群落物种均匀度和丰富度的综合指标, 均匀度是指群落中每一物种数量的分配比例[41]。因此, Shannon-wiener指数、均匀度指数和丰富度指数能较全面地反映物种的多样性和植物个体在种群中分布的均匀程度。本研究主成分分析表明, RG和PG下, 地上生物量与第一主成分的关系最密切, CG下, 第一主成分上物种丰富度的载荷最高, GSG下, 第一主成分上物种Shannon-wiener指数的载荷最高(表 3), 说明不同放牧制度会导致鼠丘的群落演替因子的不同, 可能由于不同放牧管理模式下土壤水分、热量等的利用和维持不同, 供给植被生长、发育所吸收和利用的养分也不同, 因此, 鼠丘上植被的生长恢复也会表现出一定的差异性[27]。
综上所述, 不同放牧管理模式下影响鼠丘群落演替的因素不同, RG下鼢鼠扰动增加了物种Shannon-wiener指数, 而CG下鼢鼠扰动降低了物种Shannon-wiener指数和地上生物量;ZM2物种总盖度、丰富度指数和Shannon-wiener指数, 均表现为GSG高于RG、PG和CG, 且CG下均最小;PG和GSG下物种平均高度、地上生物量均高于CG和RG。因此, 合理的放牧管理模式可以有效改善高寒草甸草地的群落特征, 从而优化高原鼢鼠的管理策略。
| [1] |
彭祺, 王宁. 不同放牧制度对草地植被的影响. 农业科学研究, 2005, 26(1): 27-30. DOI:10.3969/j.issn.1673-0747.2005.01.007 |
| [2] |
Kotzé E, Sandhage-Hofmann A, Meinel J A, du Preez C C, Amelung W. Rangeland management impacts on the properties of clayey soils along grazing gradients in the semi-arid grassland biome of South Africa. Journal of Arid Environments, 2013, 97: 220-229. DOI:10.1016/j.jaridenv.2013.07.004 |
| [3] |
Miao F H, Guo Z G, Xue R, Wang X Z, Shen Y Y. Effects of grazing and precipitation on herbage biomass, herbage nutritive value, and yak performance in an alpine meadow on the Qinghai-Tibetan Plateau. PLoS One, 2015, 10(6): e0127275. DOI:10.1371/journal.pone.0127275 |
| [4] |
侯扶江, 杨中艺. 放牧对草地的作用. 生态学报, 2006, 26(1): 244-264. DOI:10.3321/j.issn:1000-0933.2006.01.031 |
| [5] |
McNaughton S J. Grazing as an optimization process:grass-ungulate relationships in the Serengeti. The American Naturalist, 1979, 113(5): 691-703. DOI:10.1086/283426 |
| [6] |
Cui X F, Graf H F. Recent land cover changes on the Tibetan Plateau:a review. Climatic Change, 2009, 94(1/2): 47-61. |
| [7] |
蒙荣, 赛音吉雅, 张小芬. 不同放牧制度大针茅草原群落特征的比较. 内蒙古草业, 2003, 15(2): 10-12. |
| [8] |
Meissner R A, Facelli J M. Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrublands of South Australia. Journal of Arid Environments, 1999, 42(2): 117-128. DOI:10.1006/jare.1999.0515 |
| [9] |
Li W, Liu Y Z, Wang J L, Shi S L, Cao W X. Six years of grazing exclusion is the optimum duration in the alpine meadow-steppe of the north-eastern Qinghai-Tibetan Plateau. Scientific Reports, 2018, 8(1): 17269. DOI:10.1038/s41598-018-35273-y |
| [10] |
苏军虎, 南志标, 纪维红. 家畜放牧对草地啮齿动物影响的研究进展. 草业学报, 2016, 25(11): 136-148. DOI:10.11686/cyxb2015587 |
| [11] |
施大钊, 钟文勤. 2000年我国草原鼠害发生状况及防治对策. 草地学报, 2001, 9(4): 248-252. |
| [12] |
Koide R T, Huenneke L F, Mooney H A. Gopher mound soil reduces growth and affects ion uptake of two annual grassland species. Oecologia, 1987, 72(2): 284-290. DOI:10.1007/BF00379280 |
| [13] |
Rebollo S, Milchunas D G, Stapp P, Augustine D J, Derner J D. Disproportionate effects of non-colonial small herbivores on structure and diversity of grassland dominated by large herbivores. Oikos, 2013, 122(12): 1757-1767. DOI:10.1111/j.1600-0706.2013.00403.x |
| [14] |
Pickett S T A. Non-equilibrium coexistence of plants. Bulletin of the Torrey Botanical Club, 1980, 107(2): 238-248. DOI:10.2307/2484227 |
| [15] |
Schaffer W M, Fox G A. Plant strategies and the dynamics and structure of plant communities. Bulletin of Mathematical Biology, 1989, 51(3): 409-411. |
| [16] |
刘双双.家畜与高原鼢鼠对高寒草甸植物多样性和生产力的影响[D].兰州: 兰州大学, 2018. http://cdmd.cnki.com.cn/Article/CDMD-10730-1018979798.htm
|
| [17] |
江小雷, 张卫国, 杨振宇, 杜国祯. 不同演替阶段鼢鼠土丘群落植物多样性变化研究. 应用生态学报, 2004, 15(5): 814-818. DOI:10.3321/j.issn:1001-9332.2004.05.017 |
| [18] |
于海玲, 樊江文, 李愈哲, 史文娇. 高原鼢鼠干扰对三江源区高寒草甸群落特征的影响. 应用生态学报, 2018, 29(6): 1902-1910. |
| [19] |
王玉琴, 鲍根生, 王宏生, 曾辉, 李继鹏. 不同放牧制度对高原鼢鼠活动区高寒草地群落结构影响. 草地学报, 2018, 26(1): 134-141. |
| [20] |
姬万忠, 王庆华. 补播对天祝高寒退化草地植被和土壤理化性质的影响. 草业科学, 2016, 33(5): 886-890. |
| [21] |
杨莹博, 辛小娟, 艾得协措, 杨雪, 王绍美, 储诚进, 任正炜, 王刚. 鼢鼠土丘植被恢复演替过程中的物种多样性变化. 草业学报, 2010, 19(1): 14-20. |
| [22] |
鲍根生, 王宏生, 曾辉. 有机硅助剂对"狼毒净"防除狼毒效果及草地植物群落多样性的影响. 草业科学, 2015, 32(2): 263-268. |
| [23] |
陈琳, 赵陟峰, 赵廷宁, 王娟. 基于因子与聚类分析的不同林分土壤理化性质评价. 水土保持研究, 2011, 18(5): 191-196. |
| [24] |
朱桂林, 山仑, 卫智军, 韩国栋. 放牧制度对短花针茅群落植物生长的影响研究. 中国生态农业学报, 2004, 12(4): 181-183. |
| [25] |
杨利民, 韩梅, 周广胜, 王玉辉. 草地群落物种多样性维持机制的研究Ⅲ.物种分布格局. 吉林农业大学学报, 2002, 24(1): 58-61. DOI:10.3969/j.issn.1000-5684.2002.01.013 |
| [26] |
Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199(4335): 1302-1310. DOI:10.1126/science.199.4335.1302 |
| [27] |
Proulx M, Mazumder A. Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient- rich ecosystems. Ecology, 1998, 79(8): 2581-2592. DOI:10.1890/0012-9658(1998)079[2581:ROGIOP]2.0.CO;2 |
| [28] |
闫玉春, 唐海萍. 围栏禁牧对内蒙古典型草原群落特征的影响. 西北植物学报, 2007, 27(6): 1225-1232. DOI:10.3321/j.issn:1000-4025.2007.06.026 |
| [29] |
Qin Y, Chen J J, Yi S H. Plateau pikas burrowing activity accelerates ecosystem carbon emission from alpine grassland on the Qinghai-Tibetan Plateau. Ecological Engineering, 2015, 84: 287-291. DOI:10.1016/j.ecoleng.2015.09.012 |
| [30] |
Klein J A, Harte J, Zhao X Q. Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecological Applications, 2007, 17(2): 541-557. DOI:10.1890/05-0685 |
| [31] |
黄彬.高原鼢鼠种群密度与植被、土壤因素的关系[D].兰州: 兰州大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10730-1014305427.htm
|
| [32] |
张卫国, 江小蕾, 王树茂, 杨振宇. 鼢鼠的造丘活动及不同休牧方式对草地植被生产力的影响. 西北植物学报, 2004, 24(10): 1882-1887. DOI:10.3321/j.issn:1000-4025.2004.10.020 |
| [33] |
张燕堃.高寒草甸鼢鼠土丘年限对植物功能特征及生产力的影响[D].兰州: 兰州大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10730-1014305425.htm
|
| [34] |
赵新全, 马玉寿, 周华坤. 三江源区退化生态系统恢复及可持续管理化. 北京: 科学出版社, 2011.
|
| [35] |
Mahdi A, Law R, Willis A J. Large niche overlaps among coexisting plant species in a limestone grassland community. Journal of Ecology, 1989, 77(2): 386-400. DOI:10.2307/2260757 |
| [36] |
Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton: Princeton University Press, 2001.
|
| [37] |
Turnbull L A, Levine J M, Loreau M, Hector A. Coexistence, niches and biodiversity effects on ecosystem functioning. Ecology Letters, 2013, 16(S1): 116-127. |
| [38] |
双全, 邢旗, 金玉, 松梅, 巴格那. 草甸草原主要植物种群动态特征对放牧制度的响应. 畜牧与饲料科学, 2010, 31(6/7): 298-300. |
| [39] |
Tilman D. Resource Competition and Community Structure. Princeton: Princeton University Press, 1982.
|
| [40] |
汪诗平, 李永宏. 内蒙古典型草原退化机理的研究. 应用生态学报, 1999, 10(4): 437-441. DOI:10.3321/j.issn:1001-9332.1999.04.015 |
| [41] |
Haynes R J. The decomposition process: mineralization, immobilization, humus formation, and degradation//Haynes R J, ed. Mineral Nitrogen in the Plant-Soil System. London: Academic Press, 1986: 52-126.
|