 2020, Vol. 40
2020, Vol. 40文章信息
- 王凌菲, 徐霞, 江红蕾, 关梦茜, 田晓宇
- WANG Lingfei, XU Xia, JIANG Honglei, GUAN Mengxi, TIAN Xiaoyu
- 内蒙古温带典型草原围封十年草灌景观格局动态
- The dynamics of landscape pattern within a ten-year exclosure in temperate typical grassland in Inner Mongolia
- 生态学报. 2020, 40(7): 2234-2241
- Acta Ecologica Sinica. 2020, 40(7): 2234-2241
- http://dx.doi.org/10.5846/stxb201901170144
-
文章历史
- 收稿日期: 2019-01-17
- 网络出版日期: 2019-12-26





2. 北京师范大学地理科学学部自然资源学院, 北京 100875;
3. 中国农业科学院农业资源与农业区划研究所, 北京 100081
2. School of Natural Resources, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China;
3. Institute of Agricultural Resources and Regional Planning of Chinese Academy of Agricultural of Sciences, Beijing 100081, China
草地作为中国占比最大的土地利用类型, 主要分布于年降水量小于400 mm的干旱半干旱地区[1], 具有较强的生态脆弱性, 在气候变化、人类活动和植被间的相互作用等多种因素综合影响下表现出的灌丛化是其退化或荒漠化的显著特征[2], 威胁草原及畜牧业生产的可持续发展。
全球大部分地区有变暖变干的趋势, 降水波动引起的土壤水分的改变必然会对植被分布格局产生影响[3-4]。研究发现, 凡能够减少草本生物量或减弱草本植物利用土壤表层水分的能力, 并引起土壤深层水分增加的因素均能促进木本植物的生长[5-7]。刘峻杉等人发现, 降雨的长期波动和干旱的影响能够增强灌木作为群落优势种的竞争性, 从而延缓草本植物在群落演替后期成为优势种的格局的出现[8];李巧燕等人在对鄂尔多斯高原草地灌丛水分利用特征的研究中表明, 灌木植物在遭受水分胁迫时表现出更强的可塑性, 能够根据土壤水分的可利用性改变对不同土层水分的利用[9]。此外, 降水的季节分布以及极端降雨事件的发生均会对草本、灌木的竞争能力产生影响[10], 从而改变草原原有的景观格局。
景观格局变化引起的草地生态过程的改变受到众多学者的关注[11-14]。景观破碎化加剧、异质性增强会导致生态系统不稳定、草本生产力降低[15]。灌丛化会改变土壤资源的空间分布格局, 影响水分、养分等资源的循环过程, 导致关键性资源分布的异质性, 进而影响生态系统功能[16-18]。目前对灌丛化背景下景观格局的特征研究多是采用空间代替时间的方法, 缺少长时间序列的精确测量。本研究采用差分GPS法对植被斑块进行空间精确定位, 结合景观生态学的研究方法, 试图揭示在排除人为放牧干扰以后研究区植被格局动态表现, 更好地理解降水事件与草原景观格局和灌丛化过程间的关系, 为我国干旱半干旱地区可持续发展提供参考。
1 研究区概况研究区位于内蒙古锡林郭勒盟太仆寺旗境内(114°51′E—115°49′E, 41°35′N—42°10′N), 海拔1300—1800 m。属半干旱大陆性气候, 冬季寒冷漫长, 夏季凉爽短暂, 常年多风, 年均温2.1℃, 年均降雨量384.5 mm。地带性土壤是较为贫瘠的栗钙土。研究区属内蒙古典型草原区, 草地占据最大的土地利用数量比例[19], 草原植被主要有克氏针茅(Stipa krylovii)、糙隐子草(Cleistogenes squarrosa)、冷蒿(Artemisia frigida)、银灰旋花(Convolvulus ammannii)等, 镶嵌着小叶锦鸡儿(Caragana microphylla)为主的灌丛斑块。属典型的农牧交错生态过渡区。
2 研究方法 2.1 样地设置于2007年10月在太仆寺旗东北黑山庙设置了草原灌木观测研究样地(图 1), 沿西北—东南向设置长400 m, 宽50 m的围栏(115°27.743′E—115°27.978′E, 42°06.934′N—42°07.100′N), 平均坡度为4.06°。围栏区域在2003年以前放牧利用至重度退化阶段, 2003年后全年轻度放牧利用, 围栏内部全年禁牧。将围封区域内划分20个样地, 每个样地为20 m×50 m(图 1)。

|
| 图 1 样地设置 Fig. 1 The setting of researching plot |
样地植被群落调查:实验观测于2007、2008、2010、2012、2013和2016年的8月进行, 在每个样地随机选取5个1 m×1 m的样方, 调查草本种类、总盖度、分盖度(针刺法)和高度等, 并且在每个样地设置3个5 m×5 m的灌木样方, 调查灌木的盖度和高度。草本和灌木生物量通过张勇等[20]在该样地建立的估算预测模型得出:
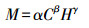
|
(1) |
式中, M为草本生物量(g/m2)时, α=68.259, C为草本盖度, H为草本高度(cm);M为灌木生物量(g/m2)时, α=62.5549, C为灌木冠幅(m2), H为灌木高度(cm)。
样地景观格局调查:按照斑块的种类和形态特征将景观类型要素分为小叶锦鸡儿灌丛斑块和草本斑块。实验观测于2010、2012、2013和2016年的8月进行, 利用差分GPS(中海达, 中国)测量样地灌丛斑块垂直投影形态, 并通过Fragstat4.2软件计算各期植被景观指数。
土壤水分的获取:每个样地中心位置安装土壤水分监测系统(ECH2O, USA), 测定土壤体积含水量。测量的土层深度分为5层, 0—15、15—30、30—45、45—60、60—75 cm。土壤表层含水量为0—30 cm平均值, 深层为30—75 cm平均值。同时, 在围栏区域的中间位置, 装有自动气象站(Dynamax, UAS), 记录相关气象数据。
土壤样品按照水分测量深度分层采样, 分别在2009年和2016年8月进行, 使用元素分析仪(PE2400, UAS)测定土壤元素含量。
试验数据的统计分析采用SPSS 23.0(SPSS Inc, Chicago, USA)完成, 统计检验方法主要为配对T检验, Pearson相关性检验, 显著水平为P < 0.05。
3 结果 3.1 降水变化如图 2所示, 研究区近10年降水整体呈增加趋势;2007—2009年为干旱年;2010—2012年总降水量较多;2013—2016年降水先减少后增加。2015年降水较少, 但仍高于近10年的平均值。在季节分配上, 降水主要集中在夏季, 而2010年秋季占全年降水的47.9%;2012年夏季降水最多, 2014年之后年降水量由于夏季降水的增加而呈现上升趋势。

|
| 图 2 季节降水和年降水变化 Fig. 2 The changing of seasonal and annual precipitation |
随着围封年份的增加, 围栏内植被盖度和生物量的变化如图 3所示:排除人为干扰后, 观测围栏内2007—2016年样地植被平均盖度和平均生物量, 发现草本的盖度增加率为207.2%, 生物量增加率为58.5%;灌木的盖度从2007到2016年增加了73.65%, 灌木生物量增加了34.93%。草本在2007—2010年, 盖度和生物量降低, 2012—2016年, 盖度和生物量增加;而灌木的盖度和生物量表现出先增加而后趋于平稳的趋势。围封10年后, 灌木和草本的盖度和生物量均显著增加, 样地植被总生产力呈现较快增加而又趋于平稳的趋势。

|
| 图 3 围封10年草本和灌木的盖度和生物量的变化 Fig. 3 The changing of coverage and biomass of grass and shrubs in different years |
按照斑块的种类将景观类型要素分为灌木(小叶锦鸡儿)和草本2种斑块类型。从图 4可知, 研究区景观类型以草本所占面积比例最大, 整体呈现以草本为基质, 小叶锦鸡儿为主体的交错景观格局。本研究选取斑块密度、斑块面积比、最大斑块指数等斑块结构指标, 景观形状指数等形状指标, 以及均匀度指数和蔓延度指数等形态指标, 探究不同年份植被景观格局的动态变化。

|
| 图 4 研究区景观格局 Fig. 4 The landscape pattern of researching area |
表 1反映了研究区域景观水平上不同景观指数的年际变化趋势。总的来说, 斑块密度和景观形状指数以2012年为拐点呈现先增加后减少的趋势, 最大斑块指数和蔓延度指数下降, 均匀度指数有所增加。
| 年份 Year |
斑块密度 PD(个/100hm2) |
最大斑块指数 LPI |
景观形状指数 LSI |
均匀度指数 SHEI |
蔓延度指数 CONTAG |
| 2010 | 65083.09 | 77.62 | 19.40 | 0.78 | 53.78 |
| 2012 | 109698.22 | 76.85 | 22.06 | 0.78 | 52.10 |
| 2013 | 93793.39 | 75.69 | 20.75 | 0.80 | 51.53 |
| 2016 | 89663.43 | 75.56 | 19.64 | 0.80 | 51.85 |
| PD, 斑块密度Patch Density; LPI, 最大斑块指数largest patch index; LSI, 景观形状指数landscape shape index; SHEI, 均匀度指数Shannon′s Evenness Index; CONTAG, 聚集度指数Contagion | |||||
在斑块类型尺度, 小叶锦鸡儿灌丛斑块和草本斑块在格局上的变化对整体景观格局的变化具有指示意义[21]。由表 2可知, 2016年小叶锦鸡儿灌丛斑块面积比相较于2010年增加了9.16%, 斑块密度和景观形状指数具有先增后减的特点。草本斑块的面积比由于灌木的增加而减少, 斑块密度和景观形状指数变化趋势与灌木类似。
| 年份 Year |
小叶锦鸡儿灌丛斑块 Caragana microphylla Lam. |
草本斑块 Grass |
|||||
| 斑块密度 PD (个/100hm 2) |
斑块面积比 PLAND/% |
景观形状指数 LSI |
斑块密度 PD(个/100hm 2) |
斑块面积比 PLAND/% |
景观形状指数 LSI |
||
| 2010 | 64837.50 | 22.38 | 36.34 | 2451.60 | 77.62 | 22.00 | |
| 2012 | 105440.53 | 23.13 | 41.74 | 4257.69 | 76.87 | 24.88 | |
| 2013 | 91536.33 | 24.31 | 38.10 | 2257.06 | 75.69 | 23.52 | |
| 2016 | 87090.08 | 24.43 | 35.50 | 2573.35 | 75.57 | 22.26 | |
| PLAND, 斑块面积比Percentage of Landscape | |||||||
干旱半干旱地区, 水分是植被生长的主要限制因子。地下水位埋藏深, 表层土壤水主要靠降水补给[22]。本研究中表层土壤含水量(d=0.079)显著大于深层(d=0.065), 如图 5所示, 使用广义最小二乘法得出二者关系为Ws(30—75)=-0.23+1.11Ws(0—30), 相关系数Φ=0.84, P < 0.01。近10年来研究区降水整体呈增加趋势, 表层(0—30 cm)和深层(30—75 cm)土壤水分动态与降水的年际变化呈现出相似的波动[23], 但是地表植被和地貌构成的微环境会影响土壤垂直方向含水量对于降水的响应[24-25], 如2011年和2013年降水量减少而深层土壤水分增加(图 5)。地表植被能够有效截流地表径流, 增加土壤水分下渗能力, 涵养水源, 提高土壤含水量[24-25];同时不同的植被覆盖类型会影响土壤水分动态, 如Qiao等的研究表明, 灌木入侵导致土壤径流显著减少, 水分下渗过程增加[26], 改变土壤水分分布;彭海英等人通过染色剂示踪法发现, 相对于草地斑块而言, 灌丛斑块下土壤水分入渗速率更快, 导水率更强[27]。植被的蒸腾作用同时也会影响降水在垂直方向的再分配[28];灌木深根系不仅能够将表层土壤水传递到深层土壤中, 同时也能将深层水和养分提升至表层, 供给草本利用[29]。

|
| 图 5 土壤表层含水量和深层含水量年均动态和回归关系 Fig. 5 The dynamics of yearly average top soil water content and bottom soil water, the regression relationship |
灌木与草本表现的差异性可能与降水有关。2007—2010年为干旱的时期, 夏季降水少(图 2)。灌木与草本在形态结构与生理活动上具有较大的差异, 灌木具有多枝的形态特征, 在受到水分胁迫时有利于降低导管栓塞, 增加水流路径[30], 且小叶锦鸡儿的网状叶脉有利于水分在叶片系统中的运输[31], 表现出更强的耐旱性, 同时草本茎干的木质化程度低于灌木, 木质部结构的安全性更差, 在发生干旱事件时, 草本比灌木更容易由于水分胁迫而死亡, 长期的干旱会降低草本的盖度和生物量。2010年的降水主要在秋季, 灌木生长季较长, 从而有利于灌木的建群[3, 32]。并且有研究表明, 在围封初期, 草本还没来得及恢复到生产力较高的组分, 为灌木的扩张提供了契机[33-35];草本和灌木之间的竞争会导致系统中水分和养分等的利用效率降低, 从而抑制草本生产力的恢复[12]。本研究对20个观测点2007年与2010年两年草本和灌木的盖度增加量做相关分析, 发现两者的相关系数为r=-0.28(P=0.23), 不能很好的解释0.1—10 m这个尺度上草本和灌木盖度变化之间的关系, 二者并未形成绝对竞争, 但是显示了灌、草此消彼涨的大致趋势。
2011—2016年期间, 研究区生长季降雨量增多, 处于一个较为湿润的时期。草本生物体的构建需要较少的物质和能量, 对降水响应敏感迅速, 能够快速吸收表层土壤水, 出现草本盖度的剧增。2012年降水比较充分, 促进了土壤种子库短命植物种子的萌发, 植被盖度增大, 生物量增加[13]。Westoby认为干旱区草地特殊的降雨脉冲事件可能会驱动群落演替的跃迁[36]。围封6年后, 草本和灌木呈现同步增长, 出现了各居其地共存的现象。有研究表明草本的盖度和生物量均随着表层土壤含水量的增加而显著增加, 灌木的盖度和生物量均随着深层土壤含水量的增加而增加, 因“两层水分假设”而共存[37], 并且这种关系受到年际降水量的波动的影响[38]。
有研究表明小叶锦鸡儿的扩张对草本的生长有促进作用:一是小叶锦鸡儿的滞雪作用增加了周围土壤含水量, 供给植物春季返青生长的需求[39]。二是豆科植物在恢复早期能改善土壤养分, 植被枯枝落叶归还土壤, 加强土壤氮固持[40-43]。本研究也发现围封禁牧后, 研究区土壤总氮含量显著增加(图 6), 为草本的恢复创造了稳定的土壤环境。土壤水分条件良好时有利于增强小叶锦鸡儿根系根瘤的活性, 增强小叶锦鸡儿固定大气中的氮的能力, 从而减弱与草本植物对土壤中的氮的竞争。灌木植物根系固氮后通过凋落物以及根系周转而在表层土壤富集[44], 并且小叶锦鸡儿增加的固氮量能够通过根系转移到与其相邻的草本植物根系周围, 从而促进草本植物的生长[45], 再通过“沃岛”向周围传播和散布[46]。

|
| 图 6 2009和2016年土壤总氮含量的比较 Fig. 6 The comparing of total nitrogen between 2009 and 2016 |
植物种群的分布格局随着环境的变化而变化, 并且受其自身的生物学特征(如繁殖方式等)、围封放牧干扰、种间关系(如竞争)等因素的影响[47]。灌丛斑块分布格局是灌木对干旱缺水的生存环境进行自我调节、适应和选择的外在表现[15]。景观尺度上, 优势斑块草本的面积逐渐减小且斑块间的连通度降低, 2010—2012年破碎化加剧, 2012年后破碎化程度有所下降。从小叶锦鸡儿灌丛斑块类型水平上看, 小叶锦鸡儿灌丛斑块的扩张过程导致了样地景观格局的变化, 提高了样地的均匀度。2012年之前小叶锦鸡儿克隆生长, 破碎化程度加剧, 蔓延度下降, 但提高了景观水平的均匀度, 灌丛斑块和草本斑块边界复杂, 物质和能量交换最为频繁[21];2012年之后小叶锦鸡儿由点及面, 灌丛斑块不断扩大, 逐渐形成更聚集的空间分布方式, 成为“沃岛”, 使得景观格局破碎化程度降低, 蔓延度提高, 这可能有利于通过种内聚集而降低种间的竞争强度, 从而有利于恢复灌丛在群落中的地位[48]。
5 结论围封10年之后, 典型草原的生产力逐渐恢复, 但灌木的扩张也指示着草原的退化;在干旱的时期, 降水的缺乏会加剧草灌之间的竞争, 灌木生长状况优于草本, 而在水资源相对充足时, 灌木对草本生长的促进作用更为明显;景观格局的破碎化程度与灌木的扩张并非线性关系, 受到灌木的繁殖方式及其与草本之间竞争的影响。灌木的扩张过程逐渐影响草原生态系统的景观格局及其功能, 促使生态系统得以自我恢复和重建。同时对于草原生态系统的管理, 不能简单地采用完全禁牧围封的方式, 要综合考虑区域气候、养分条件、植被间相互作用等因素, 在干旱年份注意对灌木扩张的防范, 从而保证草场的可持续利用。
| [1] |
江红蕾, 徐霞, 成亚薇, 李霞, 田晓宇. 基于生态可持续的土地利用空间格局分配. 中国农学通报, 2017, 33(33): 67-73. |
| [2] |
李新荣.干旱沙区土壤空间异质性变化对植被恢复的影响//中国生态学会2006学术年会论文荟萃.沈阳: 中国生态学学会, 2006: 361-370.
|
| [3] |
Brown J H, Valone T J, Curtin C G. Reorganization of an arid ecosystem in response to recent climate change. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(18): 9729-9733. |
| [4] |
Yahdjian L, Sala O E. Size of precipitation pulses controls nitrogen transformation and losses in an arid Patagonian ecosystem. Ecosystems, 2010, 13(4): 575-585. |
| [5] |
Gao Q, Reynolds J F. Historical shrub-grass transitions in the northern Chihuahuan Desert:modeling the effects of shifting rainfall seasonality and event size over a landscape gradient. Global Change Biology, 2003, 9(10): 1475-1493. |
| [6] |
Duniway M C, Snyder K A, Herrick J E. Spatial and temporal patterns of water availability in a grass-shrub ecotone and implications for grassland recovery in arid environments. Ecohydrology, 2010, 3(1): 55-67. |
| [7] |
Li X Y, Yang Z P, Li Y T, Lin H. Connecting ecohydrology and hydropedology in desert shrubs:stemflow as a source of preferential flow in soils. Hydrology and Earth System Sciences, 2009, 13(7): 1133-1144. |
| [8] |
刘峻杉, 徐霞, 张勇, 田玉强, 高琼. 长期降雨波动对半干旱灌木群落生物量和土壤水分动态的效应. 中国科学:生命科学, 2010, 40(2): 166-174. |
| [9] |
李巧燕, 来利明, 周继华, 姜联合, 杨柳, 易三桂, 孙清琳, 郑元润. 鄂尔多斯高原草地灌丛化不同阶段主要植物水分利用特征. 生态学杂志, 2019, 38(1): 89-96. |
| [10] |
Scheiter S, Higgins S I. Impacts of climate change on the vegetation of Africa:an adaptive dynamic vegetation modelling approach. Global Change Biology, 2009, 15(9): 2224-2246. |
| [11] |
韩砚君, 牛建明, 张庆, 董建军, 张雪峰, 康萨如拉. 锡林河流域近30年草原植被格局动态及驱动力分析. 中国草地学报, 2014, 36(2): 70-77. |
| [12] |
彭海英, 李小雁, 童绍玉. 内蒙古典型草原灌丛化对生物量和生物多样性的影响. 生态学报, 2013, 33(22): 7221-7229. |
| [13] |
宋彦涛, 乌云娜, 张靖, 霍光伟, 张凤杰. 放牧强度对克氏针茅(Stipa krylovii)草原植被景观格局的影响. 中国沙漠, 2016, 36(3): 674-680. |
| [14] |
王永利, 云文丽, 梁存柱, 王炜. 内蒙古典型草原区植被格局变化及退化导因探讨. 干旱区资源与环境, 2007, 21(10): 144-149. |
| [15] |
郑肖然, 李小雁, 李柳, 彭海英, 张思毅. 干旱半干旱区灌丛斑块与降水量响应关系的熵模型模拟. 生态学报, 2015, 35(23): 7803-7811. |
| [16] |
熊小刚, 韩兴国. 内蒙古退化草原中与小叶锦鸡儿相关的小尺度土壤碳、氮资源异质性动态. 生态学报, 2006, 26(2): 483-488. |
| [17] |
熊小刚, 韩兴国. 资源岛在草原灌丛化和灌丛化草原中的作用. 草业学报, 2006, 15(1): 9-14. |
| [18] |
Donohue I, Styles D, Coxon C, Irvine K. Importance of spatial and temporal patterns for assessment of risk of diffuse nutrient emissions to surface waters. Journal of Hydrology, 2005, 304(1/4): 183-192. |
| [19] |
田晓宇, 徐霞, 江红蕾, 龚容, 关梦茜. 退耕还林(草)政策下土地利用结构优化研究——以内蒙古太仆寺旗为例. 中国人口·资源与环境, 2018, 28(S2): 25-30. |
| [20] |
张勇.基于温带半干旱草原围封坡面样地连续观测的植被与土壤水分关系.北京: 北京师范大学, 2011.
|
| [21] |
邬建国. 景观生态学:格局、过程、尺度与等级(第二版). 北京: 高等教育出版社, 2007.
|
| [22] |
张军红, 吴波. 干旱、半干旱地区土壤水分研究进展. 中国水土保持, 2012(2): 40-43. |
| [23] |
Knapp A K, Fay P A, Blair J M, Collins S L, Smith M D, Carlisle J D, Harper C W, Danner B T, Lett M S, McCarron J K. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science, 2002, 298(5601): 2202-2205. |
| [24] |
Amani M, Salehi B, Mahdavi S, Masjedi A, Dehnavi S. Temperature-vegetation-soil moisture dryness index (TVMDI). Remote Sensing of Environment, 2017, 197: 1-14. |
| [25] |
Chen Z J, Mohanty B P, Rodriguez-Iturbe I. Space-time modeling of soil moisture. Advances in Water Resources, 2017, 109: 343-354. |
| [26] |
Qiao L, Zou C B, Stebler E, Will R E. Woody plant encroachment reduces annual runoff and shifts runoff mechanisms in the tallgrass prairie, USA. Water Resources Research, 2017, 53(6): 4838-4849. |
| [27] |
彭海英, 童绍玉, 李小雁. 内蒙古典型草原土壤及其水文过程对灌丛化的响应. 自然资源学报, 2017, 32(4): 642-653. |
| [28] |
Ludwig J A, Wilcox B P, Breshears D D, Tongway D J, Imeson A C. Vegetation patches and runoff-erosion as interacting ecohydrological processes in semiarid landscapes. Ecology, 2005, 86(2): 288-297. |
| [29] |
Richards J H, Caldwell M M. Hydraulic lift:substantial nocturnal water transport between soil layers by Artemisia tridentata roots. Oecologia, 1987, 73(4): 486-489. |
| [30] |
龚容, 徐霞, 田晓宇, 江红蕾, 李霞, 关梦茜. 三种锦鸡儿属植物水力结构特征及其干旱适应策略. 生态学报, 2018, 38(14): 4984-4993. |
| [31] |
龚容, 徐霞, 江红蕾, 田晓宇, 李霞, 关梦茜. 干旱半干旱区几种典型灌木半灌木茎叶水分传导系统的结构特征. 北京师范大学学报(自然科学版), 2018, 54(04): 534-542. |
| [32] |
Neilson R P. High-resolution climatic analysis and southwest biogeography. Science, 1986, 232(4746): 27-34. |
| [33] |
Adler P B, Milchunas D G, Sala O E, Burke I C, Lauenroth W K. Plant traits and ecosystem grazing effects:comparison of U.S. sagebrush steppe and Patagonian steppe.. Ecological Applications, 2005, 15(2): 774-792. |
| [34] |
Cipriotti P A, Aguiar M R. Direct and indirect effects of grazing constrain shrub encroachment in semi-arid Patagonian steppes. Applied Vegetation Science, 2012, 15(1): 35-47. |
| [35] |
高琼, 刘婷. 干旱半干旱区草原灌丛化的原因及影响-争议与进展. 干旱区地理, 2015, 38(6): 1202-1212. |
| [36] |
Westoby M, Walker B, Noy-Meir I. Opportunistic management for rangelands not at equilibrium. Journal of Range Management, 1989, 42(4): 266-274. |
| [37] |
Walter H. Ecology of Tropical and Subtropical Vegetation. Edinburgh: Oliver and Boyd, 1971.
|
| [38] |
Ogle K, Reynolds J F. Plant responses to precipitation in desert ecosystems:integrating functional types, pulses, thresholds, and delays. Oecologia, 2004, 141(2): 282-294. |
| [39] |
田晓艳.内蒙古羊草草原暗栗钙土水分动态变化研究[D].呼和浩特: 内蒙古农业大学, 2007. http://cdmd.cnki.com.cn/article/cdmd-10129-2007219468.htm
|
| [40] |
白日军, 杨治平, 张强, 张训忠. 晋西北不同年限小叶锦鸡儿灌丛土壤氮矿化和硝化作用. 生态学报, 2016, 36(24): 8008-8014. |
| [41] |
李香真, 张淑敏, 邢雪荣. 小叶锦鸡儿灌丛引起的植物生物量和土壤化学元素含量的空间变异. 草业学报, 2002, 11(1): 24-30. |
| [42] |
Barger N N, Archer S R, Campbell J L, Huang C Y, Morton J A, Knapp A K. Woody plant proliferation in North American drylands:a synthesis of impacts on ecosystem carbon balance. Journal of Geophysical Research, 2011, 116(G4): G00K07. |
| [43] |
邵新庆, 王堃, 王赟文, 刘贵河. 典型草原自然恢复演替过程中植物群落动态变化. 生态学报, 2008, 28(2): 855-861. |
| [44] |
Boutton T W, Liao J D. Changes in soil nitrogen storage and δ15N with woody plant encroachment in a subtropical savanna parkland landscape. Journal of Geophysical Research, 2010, 115(G3): G03019. |
| [45] |
苏永中, 赵哈林, 张铜会. 几种灌木、半灌木对沙地土壤肥力影响机制的研究. 应用生态学报, 2002, 13(7): 802-806. |
| [46] |
左小安, 赵学勇, 赵哈林, 郭轶瑞, 李玉霖, 刘任涛, 毛伟. 沙地退化植被恢复过程中灌木发育对草本植物和土壤的影响. 生态环境学报, 2009, 18(2): 643-647. |
| [47] |
刘振国, 李镇清. 植物群落中物种小尺度空间结构研究. 植物生态学报, 2005, 29(6): 1020-1028. |
| [48] |
晁增国, 汪诗平, 徐广平, 胡宜刚, 张振华. 围封对退化矮嵩草草甸群落结构和主要种群空间分布格局的影响. 西北植物学报, 2008, 28(11): 2320-2326. |