 2020, Vol. 40
2020, Vol. 40文章信息
- 王婷, 杨思维, 花蕊, 楚彬, 叶国辉, 牛钰杰, 唐庄生, 花立民
- WANG Ting, YANG Siwei, HUA Rui, CHU Bin, YE Guohui, NIU Yujie, TANG Zhuangsheng, HUA Limin
- 高寒草原植物功能群组成对退化程度的响应
- Response characteristics of composition of plant functional groups to various grassland degradation conditions in alpine steppe on the Tibetan Plateau, China
- 生态学报. 2020, 40(7): 2225-2233
- Acta Ecologica Sinica. 2020, 40(7): 2225-2233
- http://dx.doi.org/10.5846/stxb201811082430
-
文章历史
- 收稿日期: 2018-11-08
- 网络出版日期: 2019-12-26





2. 毕节市畜牧兽医科学研究所, 毕节 551700
2. Institute of Animal and Veterinary Science, Bijie 551700, China
青藏高原是陆地生态系统中重要的组成部分[1-2], 而高寒草原作为青藏高原区主要的草地类型, 其面积占全国草地总面积的10.6%[3], 在全球碳、氮循环、生物多样性维护、水土保持、畜牧业发展等方面发挥着重要的作用[4-8]。高寒草原生态系统是一个集土壤-微生物-植物-动物-人类活动于一体的多界面复合体, 由于其自身的脆弱性, 加之人类不合理利用以及干扰造成该生态系统出现不同程度的退化[9]。植物群落特征同草地退化具有协同性, 植物对退化应对策略的分异使得植物群落组成发生改变, 进而影响草地植物群落的分布格局[10]。植物功能群是研究植物随环境因子变化的基本单元, 植物通过功能类群重组来适应复杂多变的环境[11]。且植被与土壤的相互关系也一直是生态学研究的重点领域[12-13]。退化不仅影响着植物群落物种组成及其演替, 而且对土壤理化属性有重要影响[9, 14], 退化过程中植物群落通过功能群的重新调整可以对土壤因子变化进行选择性适应[15], 使功能群落与土壤属性间达到一种反馈平衡。目前, 关于土壤特性和植被特征已开展了大量研究。杜际增等[16]在长江黄河源区对近45年高寒草地退化特征及成因分析的研究表明, 高寒草地呈现以覆盖度降低、破碎化与干旱化加剧为主的退化趋势, 气温升高引起的暖干化是导致高寒草地退化的主因, 过度放牧和人类不合理利用是导致草地退化加剧的重要因素;周华坤等[17]对紫花针茅高寒草原植被和土壤退化特征的研究结果表明, 随着高寒草原退化加剧, 植被盖度、草地质量指数和地上生物量比例下降, 草地间的相似性指数减小, 土壤湿度、土壤紧实度、土壤有机质、速效磷、硝态氮、速效钾的含量都减少;韩立辉等[18]对青藏高原典型“黑土滩”区秃斑块及周边的土壤和植被特征以及变化规律的研究结果表明, 秃斑块是草地养分流失的通道, 且秃斑面积大小与其周边景观异质性有关。上述研究从不同的角度研究草地的结构、功能及其对退化的响应, 对理解不同退化过程对土壤特性、植被结构与功能及其影响因素具有积极作用, 但研究多集中于植物群落单一特征(如高度、盖度或丰富度等), 有关高寒草原功能群组成对草地退化响应的研究仍然匮乏[19]。
因此, 本研究以黄河源区高寒草地中最具代表性的紫花针茅(Stipa purpurea)高寒草原为研究对象, 在青海省果洛州玛多县选择不同退化程度的高寒草原, 研究其土壤理化性状和植物功能群变化特征, 采用多元排序的方法, 以期阐明高寒草原退化过程中的植被群落与土壤理化属性之间的关系, 为高寒草原植被恢复重建提供科学依据。
1 材料与方法 1.1 研究区概况研究区位于青海省玛多县(33°50′ — 35°40′ N、96°50′ — 99°20′ E), 行政上隶属青海省果洛藏族自治州, 平均海拔在4200 m以上, 空气稀薄, 气候寒冷。玛多县属于高原大陆性半湿润气候, 年均温-5.3—-2.4 ℃, 最冷月1月的平均温度为-12.6 ℃, 最热月7月的平均气温为9.7 ℃, 无绝对的无霜期, 年均降水量247.8 — 484.8 mm。植被类型以高寒草原为主, 优势物种为紫花针茅(Stipa purpurea)、二裂委陵菜(Potentilla bifurca)和火绒草(Leontopodium nanum)为主[17]。
1.2 试验设计本试验以空间代替时间的方法, 在试验区域选取5块处于不同退化程度的样地, 按退化程度从轻到重依次标记为样地1到样地5, 样地面积均为100 m × 100 m。在上述5块样地中随机设置5个样方(50 cm × 50 cm)。于2017年8月初进行植物群落调查与土壤取样。各退化样地内植物群落高度、盖度和组成结构有明显分异, 随退化程度加剧群落盖度递减(表 1)。
| 样地号 Plot No. |
经度 Longitude |
纬度 Latitude |
海拔 Altitude/m |
盖度 Coverage/% |
群落高度 Community height/cm |
群落物种组成 Community species Composition |
退化程度 Degradation gradient |
| 1 | 97°54′42″E | 34°54′42″N | 4320 | 94.00±1.14a | 9.75±0.54a | 紫花针茅Stipa purpurea 矮嵩草Kobresia humilis 早熟禾Poa crymophila |
未退化 |
| 2 | 97°40′10″E | 35°02′27″N | 4305 | 76.00±3.27b | 3.12±0.18b | 紫花针茅Stipa purpurea 早熟禾Poa crymophila 多茎委陵菜Potentilla multicaulis |
轻度退化 |
| 3 | 97°35′33″E | 35°04′09″N | 4375 | 70.00±3.94b | 3.54±0.77b | 二裂委陵菜Potentilla bifurca 紫花针茅Stipa purpurea 独活Heracleumhemsleyanum |
中度退化 |
| 4 | 97°39′35″E | 35°07′00″N | 4369 | 44.40±6.92c | 1.64±0.11c | 西伯利亚蓼Polygonum sibiricum 柔软紫菀Aster flaccidus 赖草Leymus secalinus (Georgi) Tzvel. |
重度退化 |
| 5 | 97°39′17″E | 35°06′00″N | 4390 | 16.80±3.51d | 4.09±0.32b | 沙生凤毛菊Saussurea arenaria 矮丛风毛菊Saussurea eopygmaea |
极度退化 |
| 同一列中不同的小写字母代表不同处理间的差异显著性(P < 0.05) | |||||||
在每个50 cm × 50 cm样方中, 用针刺法测定每一物种的盖度, 每一物种随机选取10株, 分别测量其高度;在每块样地内随机抛样圆30次, 记录每一物种出现的频度。用最大值标准化法计算物种高度、盖度和频度的相对值。根据通用的植物功能性划分方法将物种划分为3个功能群(禾本科、莎草科、杂类草)[20]。
1.3.2 土壤取样土壤取样在样方附近, 土壤取样在样方附近用直径为7 cm的土钻分别取0—10、10—20、20—30 cm土层的土样, 每一层3个重复, 装入自封袋带回实验室进行风干, 之后过2 mm土壤筛去除根系和砾石, 然后进行土壤化学指标分析。土壤容重和质量含水量用环刀(100 cm3)分别取0—10、10—20、20—30 cm土层的土样, 每一层3个重复, 之后装入铝盒带回实验室称其鲜重, 再在烘箱105 ℃下烘干至恒重称其干重。
1.3.3 主要测定指标及方法土壤有机质采用重铬酸钾氧化-稀释热法测定;土壤样品经浓硫酸和混合催化剂(硫酸钠-硫酸铜-硒)消解样品后, 全氮用AA3型连续流动分析仪测定, 全磷用钼锑抗比色法测定, 全钾用火焰光度计法测定;土壤pH测定采用pHSJ-4A型pH计[21]。
1.4 数据分析 1.4.1 功能群内物种丰富度、盖度和重要值功能群的物种丰富度用每个样方各功能群内物种数的多少表示。功能群盖度用各功能群内物种的分盖度之和表示。物种重要值计算公式为:
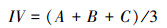
|
式中, A 为相对盖度;B为相对频度;C为相对高度。功能群重要值用各功能群植物的重要值之和表示。
相对重要值计算公式为:
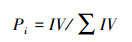
|
式中, IV为重要值。
1.4.2 土壤通气孔隙度土壤总孔隙度计算公式为:
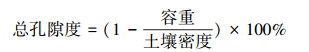
|
土壤通气孔隙度计算公式为:

|
数据利用Excel 2010进行初步处理, 采用Spass 19.0进行方差分析, 用LSD法进行多重比较, Canoco 4.5软件进行多元排序。多元排序分析中使用功能群重要值作为每种功能群基础数据, 排序之前做除趋势对应分析[22]。采用RDA冗余分析综合分析各功能群组成与土壤退化作用下土壤因子的关系。为评估环境因子对功能群组成影响的相对重要性, 分别计算每个环境因子的总效应和净效应[23]。所有排序的显著性均由MonteCarlo随机置换(499次)检验进行检验, 采用CanocoDraw绘制RDA排序图。
2 结果与分析 2.1 不同退化程度下植物功能群特征本研究共调查到20科36属48种植物。随退化程度的加剧群落结构由紫花针茅和矮嵩草群落向杂类草类群转变(表 1)。对于功能群物种丰富度, 禾本科和杂类草的物种丰富度均在中度退化时达到最大值(样地3)。对于功能群盖度, 禾本科、莎草科均随退化程度的加剧而呈显著降低(P < 0.05)的趋势, 而杂类草盖度随退化程度的加剧呈先增加后降低的趋势(表 2)。随退化程度加剧, 群落总重要值呈先增后降的趋势, 禾本科和莎草科的重要值呈降低的趋势, 而杂类草的重要值呈增加的趋势(图 1)。对于群落中不同功能群的重要值比例, 禾本科和莎草科的相对重要值随退化程度呈降低的趋势, 而杂类草的相对重要值随退化程度加剧而增加(图 1)。
| 样地号 Plot |
科 Family |
属 Genera |
种 Species |
功能群丰富度 Functional group richness |
功能群盖度 Functional group coverage/% |
地上生物量 Above-ground Biomass/ (g/m2) |
|||||
| 禾本科 Gramineae |
莎草科 Sedge |
杂类草 Forb |
禾本科 Gramineae |
莎草科 Sedge |
杂类草 Forb |
||||||
| 1 | 6 | 15 | 17 | 3.00±0.00b | 1.40±0.24b | 12.00±0.00b | 51.93±4.43a | 39.16±3.15a | 30.00±4.15ab | 33.00±1.86ab | |
| 2 | 7 | 12 | 15 | 2.00±0.00c | 2.00±0.00a | 8.80±0.58c | 46.39±3.01a | 4.37±1.49b | 48.20±7.44a | 40.10±3.38a | |
| 3 | 12 | 20 | 23 | 4.20±0.20a | 1.20±0.20b | 15.60±0.40a | 39.50±4.96a | 6.22±2.09b | 35.80±8.10ab | 29.44±3.90bc | |
| 4 | 12 | 16 | 22 | 4.00±0.00a | 2.00±0.00a | 15.20±0.37a | 7.80±5.64b | 2.60±1.94b | 38.40±3.14ab | 22.58±3.03cd | |
| 5 | 8 | 8 | 9 | 0.00±0.00d | 0.00±0.00c | 8.20±0.20c | 0.00±0.00b | 0.00±0.00b | 24.20±3.97b | 19.61±2.73d | |
| 同一列中不同的小写字母代表不同处理间的差异显著性(P <0.05) | |||||||||||

|
| 图 1 不同退化程度下植物功能群重要值变化 Fig. 1 The variation of functional group important value in the plots under different degree of degradation |
土壤容重均随土层深度增加而增加, 而土壤含水量、土壤通气孔隙度均随土层深度增加而显著降低(P < 0.05)。不同土层深度, 0—30 cm土层的土壤容重随退化程度加剧而显著增加(P < 0.05);0—30 cm土层的土壤通气孔隙度随退化程度加剧而显著降低的趋势(P < 0.05);而0—30 cm土层的土壤含水量均随退化程度加剧呈先降低后增加再降低的趋势(图 2)。

|
| 图 2 不同退化程度下土壤理化性状比较 Fig. 2 The comparison of soil physical and chemistry properties in the plots under different degree of degradation 图中不同的小写字母代表不同处理间的差异显著性(P < 0.05) |
在不同退化程度, 土壤全氮、全磷、全钾含量和pH随土层深度增加变化规律不同。土壤有机碳含量均随土层深度增加而显著降低(P < 0.05)。而不同土层深度, 0—30 cm土层土壤有机碳和全氮随退化程度加剧呈显著降低的趋势(P < 0.05), 但样地4有所增加;全磷、全钾、pH随退化程度加剧未表现一致的变化规律。
2.3 不同退化程度下功能群组成与土壤因子的关系对试验样地的25个样方植物功能群重要值和土壤因子数据进行多元分析可知(图 3), 各功能群重要值与土壤通气孔隙度呈正相关关系, 各功能群重要值均随土壤通气孔隙度的增加而增加;各功能群重要值与土壤容重呈负相关关系, 各功能群重要值均随土壤容重的增加而降低。各功能群重要值与土壤全氮和土壤有机质呈正相关关系。

|
| 图 3 不同退化程度样地功能群随土壤因子变化 Fig. 3 The relationships between plant functional group and soil factor |
经Monte Carlo随机置换检验(表 3), 土壤通气孔隙度、容重、有机质、pH和全氮含量对禾本科功能群重要值方差解释量达到极显著水平(P < 0.01), 土壤含水量、土壤全磷和土壤全钾对禾本科功能群重要值方差解释量达到显著水平(P < 0.05);除去协同变量影响, 环境解释变量中土壤有机质、土壤pH、土壤全磷和土壤含水量对禾本科功能群重要值方差解释量达到显著水平(P < 0.05)。对于莎草科来说, 土壤有机质对其功能群重要值方差解释量最大, 达到极显著水平(P < 0.01);除去协同变量影响, 土壤全磷和土壤含水量对莎草科功能群重要值方差解释量达到显著水平(P < 0.05)。对于杂类草来说, 水分对其功能群重要值方差解释量最大, 达到极显著水平(P < 0.01);除去协同变量影响, 土壤全磷对莎草科功能群重要值方差解释量达到极显著水平(P < 0.01), 土壤pH和土壤含水量对莎草科功能群重要值方差解释量达到显著水平(P < 0.05)。
| 环境因子 Environmental factors |
禾本科Gramineae | 莎草科Sedge | 杂类草Forb | |||||||||||
| 总效应 | F | 净效应 | F | 总效应 | F | 净效应 | F | 总效应 | F | 净效应 | F | |||
| 土壤含水量 Soil water content |
0.183 | 5.135* | 0.015 | 3.092* | 0.210 | 6.111* | 0.016 | 3.626* | 0.259 | 8.031** | 0.045 | 3.307* | ||
| 土壤容重 Soil bulk density |
0.554 | 28.524** | 0.002 | 0.568 | 0.646 | 41.930** | 0.002 | 0.556 | 0.224 | 6.650** | 0.004 | 0.302 | ||
| 土壤通气孔隙度 Soil ventilation porosity |
0.560 | 29.214** | 0.003 | 0.426 | 0.633 | 39.635** | 0.001 | 0.329 | 0.219 | 6.452** | 0.008 | 0.568 | ||
| 土壤有机质 Soil organic matter |
0.423 | 16.855** | 0.027 | 5.664* | 0.736 | 64.077** | 0.005 | 1.237 | 0.201 | 5.771** | 0.030 | 2.228 | ||
| 土壤全氮 Soil total nitrogen |
0.344 | 12.050** | 0.009 | 1.923 | 0.697 | 52.879** | 0.003 | 0.694 | 0.201 | 5.790** | 0.048 | 3.536* | ||
| 土壤全磷 Soil total phosphorus |
0.202 | 5.805* | 0.019 | 3.962* | 0.387 | 14.547** | 0.016 | 3.650* | 0.176 | 4.899** | 0.092 | 6.792** | ||
| 土壤全钾 Soil total potassium |
0.152 | 4.118* | 0.008 | 1.732 | 0.163 | 4.466* | 0.001 | 0.242 | 0.162 | 4.441** | 0.010 | 0.775 | ||
| 土壤pH Soil pH | 0.355 | 12.651** | 0.02 | 4.218* | 0.067 | 1.650 | 0.001 | 0.340 | 0.209 | 6.062** | 0.061 | 4.526* | ||
| *表示在0.05水平(双侧)上显著相关, **表示在0.01水平(双侧)上显著相关; | ||||||||||||||
植物功能群是研究植物随环境因子变化的基本单元, 植物通过功能类群重组来适应复杂多变的环境[15]。本研究发现, 随退化程度加剧, 禾本科、杂类草功能群的物种丰富度和重要值均呈先增后降的趋势, 而前者盖度呈降低趋势, 后者盖度先增后降且其相对重要值逐渐增加。因为草地退化是多因素主导的结果, 放牧干扰就是其中重要因素之一。放牧强度直接作用于植物本身, 使植物功能群组成特征发生改变, 导致物种的生态位改变, 进而形成退化梯度下的不同生态适应对策[14]。主要表现为:(1)优良牧草(禾本科和莎草科)的生态位首先收缩;(2)阔叶类杂草生态位趁机扩张造成杂类草植物比例的增加;(3)禾本科和莎草科等直立型植物的生长受限最终退出生态系统, 杂类草植物最终也慢慢退化, 其中部分一年生植物和沙生植物增加, 如灰绿藜(Chenopodium glaucum L.)和沙生凤毛菊(Saussurea arenaria)等。这与周华坤[17]、马世震[24]以及牛钰杰[25]等在高寒草地的研究结果一致。
土壤化学属性和物理属性是评价土壤系统质量优劣的重要指标, 也是评价草地生态系统可否可持续发展的基础[14]。本研究发现, 随退化程度加剧, 土壤有机质、全氮、pH以及全磷等土壤养分特征随退化程度增加基本表现为降低的趋势。这可能是由于家畜的践踏造成土壤机械组成发生变化, 风蚀、水蚀作用下土壤中富含营养物质的土壤黏粉粒含量降低[26];其次家畜践踏使得枯落物增多以及家畜粪尿回归等对土壤养分具有正反馈效应[14], 因此造成部分土壤养分特征随退化程度加剧降低趋势并非十分明显。而全钾含量无明显的变化则可能是我国北方为富钾地区的原因。本研究中土壤物理属性随退化程度表现出一致的变化规律。各退化区样地, 随土层深度增加, 土壤容重增加, 而土壤含水量和通气孔隙度降低。随退化程度加剧, 0—30 cm各土层深度的容重均增加、土壤通气孔隙度降低、含水量先降低后增加再降低的趋势, 这是由于三江源区高寒草原的退化使植被盖度、生物量等降低导致土壤裸露, 水分蒸发加快, 且物种的减少也会导致植物对土壤水分利用的降低[26];另一方面草地沙化的加剧, 加之放牧的作用对土壤物理属性的影响, 使土壤容重增大, 土壤孔隙度变小, 土壤蓄水能力变差[24]。因此, 退化对土壤物理属性的影响具有确定性, 且土壤结构在退化过程中不易反弹, 且随时间累积的作用越来越明显[27]。
3.2 高寒草原退化过程中土壤因子对植物功能群组成的影响植物功能群是具有确定植物功能特征的一系列植物的组合, 是研究植物群落随环境因子动态变化的基本单元。高寒草原植物功能群组成特征是气候、土壤、水分、地形及植物本身的生物学特性等多种因素长期共同影响的结果[28], 而土壤的异质性可以对植物提供更多生态位资源, 提高群落的物种多样性, 同时植物对土壤养分条件的需求必然影响到个体或群体在群落中的分布。不同植物功能群应对退化的变化不一致, 说明不同功能类型的植物对环境因子变异的适应性, 形成不同环境因子梯度下的生态对策[29]。
草地植物多样性与土壤环境因子间具有一定的相关性[30], 这是草地生态系统维持相对稳定的基础。本研究以功能群为基本单元对植物群落进行RDA排序得出:各功能群重要值与土壤通气孔隙度的正相关性最大, 与土壤容重的负相关性最大;进一步对各功能群重要值进行冗余分析, 发现土壤物理属性对每一种功能群重要值都有重要影响。表明草地退化过程中土壤物理属性是影响高寒草原植物功能群组成的主导因素, 高寒草原通过功能群重新组合来适应土壤物理属性变异对群落所带来的影响, 形成土壤物理属性梯度下的功能群适应性变化[31]。有研究表明, 草地功能群构建过程中因环境因子限制导致物种生态位分化, 致使群落中一些物种共存而另一些物种丧失[32], 在此过程中环境因子首先决定包含哪些性状的功能群可以聚集在局域群落中[33]。除去协同变量影响, 环境解释变量中土壤全磷对杂类草功能群重要值方差解释量达到极显著水平(P < 0.01), 土壤全氮、土壤pH以及土壤含水量对其方差解释量达到显著水平(P < 0.05), 而有研究发现磷是限制草地植物生长的因子[34], 这表明除去综合因素对杂类草草地植物差异分布的影响, 退化过程中杂类草草地受土壤肥力和土壤水分的显著影响, 但在草地退化过程中部分土壤养分特征尤其是土壤全磷(图 2)随退化程度加剧降低趋势并非十分明显, 因此杂类草植物特别是沙生植物可以占据急剧退化的草地土壤资源生长和分布。
4 结论高寒草原退化对植物功能群组成特征和土壤物理属性均有一定影响。随着草地退化程度的加剧, 各功能群盖度、物种丰富度以及重要值都发生显著变化, 导致草地结构由复杂趋于简单化。草地退化使土壤容重增加, 导致土壤通气孔隙度随着草地退化程度的加剧而减小。通过冗余分析, 草地退化首先影响土壤物理属性进而影响草地功能群组成, 最终导致草地生产力下降。
| [1] |
章志龙, 刘旭东, 贾鹏, 杜国祯. 氮添加对高寒草甸植物花期物候和群落结构的影响. 草业科学, 2013, 30(5): 728-735. |
| [2] |
崔庆虎, 蒋志刚, 刘季科, 苏建平. 青藏高原草地退化原因述评. 草业科学, 2007, 24(5): 20-26. DOI:10.3969/j.issn.1001-0629.2007.05.004 |
| [3] |
段敏杰, 高清竹, 万运帆, 李玉娥, 郭亚奇, 旦久罗布, 洛桑加措. 放牧对藏北紫花针茅高寒草原植物群落特征的影响. 生态学报, 2010, 30(14): 3892-3900. |
| [4] |
McGill W B, Hunt H W, Woodmansee R G, Reuss J O, Paustian K H. Formulation, process controls, parameters and performance of PHOENIX: a model of carbon and nitrogen dynamics in grassland soils. Simulation of nitrogen behaviour of soil-plant systems: papers of a workshop, Models for the behaviour of nitrogen in soil and uptake by plant, comparison between different approaches, Wageningen, the Netherlands, January 28-February 1, 1980/editors M.J. Frissel and J.A. van Veen.
|
| [5] |
Brodie E, Edwards S, Clipson N. Soil fungal community structure in a temperate upland grassland soil. FEMS Microbiology Ecology, 2003, 45(2): 105-114. |
| [6] |
Plantureux S, Peeters A, McCracken D. Biodiversity in intensive grasslands: effect of management, improvement and challenges//Proceedings of the 13th International Occasional Symposium of the European Grassland Federation. Tartu, Estonia: EGS, 2005, 3(2): 153-164.
|
| [7] |
Osterkamp W R, Hupp C R, Stoffel M. The interactions between vegetation and erosion:new directions for research at the interface of ecology and geomorphology. Earth Surface Processes and Landforms, 2012, 37(1): 23-36. DOI:10.1002/esp.2173 |
| [8] |
Burylo M, Hudek C, Rey F. Soil reinforcement by the roots of six dominant species on eroded mountainous marly slopes (Southern Alps, France). CATENA, 2011, 84(1/2): 70-78. |
| [9] |
周艳松, 王立群, 张鹏, 梁金华, 王旭峰. 大针茅根系构型对草地退化的响应. 草业科学, 2011, 28(11): 1962-1966. |
| [10] |
林丽, 张德罡, 曹广民, 欧阳经政, 柯浔, 刘淑丽, 张法伟, 李以康, 郭小伟. 放牧强度对高寒嵩草草甸土壤养分特性的影响. 生态学报, 2016, 36(15): 4664-4671. |
| [11] |
李西良, 刘志英, 侯向阳, 吴新宏, 王珍, 胡静, 武自念. 放牧对草原植物功能性状及其权衡关系的调控. 植物学报, 2015, 50(2): 159-170. |
| [12] |
沈泽昊, 杨明正, 冯建孟, 李新辉, 彭培好, 郑智. 中国高山植物区系地理格局与环境和空间因素的关系. 生物多样性, 2017, 25(2): 182-194. |
| [13] |
唐庄生, 安慧, 邓蕾, 上官周平. 荒漠草原沙漠化植物群落及土壤物理变化. 生态学报, 2016, 36(4): 991-1000. |
| [14] |
龙瑞军, 董世魁, 胡自治. 西部草地退化的原因分析与生态恢复措施探讨. 草原与草坪, 2005, 25(6): 3-7. DOI:10.3969/j.issn.1009-5500.2005.06.001 |
| [15] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学:生命科学, 2015, 45(4): 325-339. |
| [16] |
杜际增, 王根绪, 李元寿. 近45年长江黄河源区高寒草地退化特征及成因分析. 草业学报, 2015, 24(6): 5-15. |
| [17] |
周华坤, 赵新全, 温军, 陈哲, 姚步青, 杨元武, 徐维新, 段吉闯. 黄河源区高寒草原的植被退化与土壤退化特征. 草业学报, 2012, 21(5): 1-11. |
| [18] |
韩立辉, 尚占环, 任国华, 王彦龙, 马玉寿, 李希来, 龙瑞军. 青藏高原"黑土滩"退化草地植物和土壤对秃斑面积变化的响应. 草业学报, 2011, 20(1): 1-6. |
| [19] |
刘晔, 许玥, 石松林, 彭培好, 沈泽昊. 金沙江干旱河谷植物群落的数量分类及其结构分异的环境解释. 生物多样性, 2016, 24(4): 407-420. |
| [20] |
高贤明, 陈灵芝. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析. 植物学报, 1998, 40(6): 553-559. DOI:10.3321/j.issn:1672-9072.1998.06.012 |
| [21] |
中国科学院南京土壤研究所. 土壤理化分析. 上海: 上海科学技术出版社, 1978.
|
| [22] |
ter Braak C J F, Smilauer P. CANOCO Reference Manual and CanoDraw for Windows User's Guide: Software for Canonical Community Ordination (Version 4.5). Microcomputer Power, Ithaca, NY, 2002.
|
| [23] |
Lososová Z, Chytrý M, Cimalová S, KropáčZ, Otýpková Z, Pyšek P, Tichý L. Weed vegetation of arable land in central Europe:gradients of diversity and species composition. Journal of Vegetation Science, 2004, 15(3): 415-422. DOI:10.1111/j.1654-1103.2004.tb02279.x |
| [24] |
牛钰杰, 杨思维, 王贵珍, 刘丽, 杜国祯, 花立民. 放牧强度对高寒草甸土壤理化性状和植物功能群的影响. 生态学报, 2018, 38(14): 5006-5016. |
| [25] |
马世震, 彭敏, 陈桂琛, 周国英, 孙箐. 黄河源头高寒草原植被退化特征分析. 草业科学, 2004, 21(10): 19-23. DOI:10.3969/j.issn.1001-0629.2004.10.005 |
| [26] |
柴雯.高寒草甸覆盖变化下土壤水分动态变化研究[D].兰州: 兰州大学, 2008. http://cdmd.cnki.com.cn/Article/CDMD-10730-2008162141.htm
|
| [27] |
伍星, 李辉霞, 傅伯杰, 靳甜甜, 刘国华. 三江源地区高寒草地不同退化程度土壤特征研究. 中国草地学报, 2013, 35(3): 77-84. DOI:10.3969/j.issn.1673-5021.2013.03.014 |
| [28] |
董全民, 赵新全, 李世雄, 王彦龙, 郑伟, 侯宪宽, 宋磊. 草地放牧系统中土壤-植被系统各因子对放牧响应的研究进展. 生态学杂志, 2014, 33(8): 2255-2265. |
| [29] |
李禄军, 曾德慧, 于占源. 物种/功能群去除实验及其在生态学中的应用. 生态学报, 2010, 30(1): 197-204. |
| [30] |
Janssens F, Peeters A, Tallowin J R B, Bakker J P, Bekker R M, Fillat F, Oomes M J M. Relationship between soil chemical factors and grassland diversity. Plant and Soil, 1998, 202(1): 69-78. |
| [31] |
张爱宁, 安沙舟, 张蕊思, 夏小伟, 卡斯达尔·努尔旦别克, 董磊. 不同退化梯度下大尤尔都斯高寒草原植物种群生态位特征. 草业科学, 2017, 34(2): 302-309. |
| [32] |
Tanentzap A J, Burrows L E, Lee W G, Nugent G, Maxwell J M, Coomes D A. Landscape-level vegetation recovery from herbivory:progress after four decades of invasive red deer control. Journal of Applied Ecology, 2009, 46(5): 1064-1072. DOI:10.1111/j.1365-2664.2009.01683.x |
| [33] |
Kraft N J B, Ackerly D D. Assembly of plant communities//Monson R K, ed. Ecology and the Environment. New York, NY: Springer, 2014.
|
| [34] |
Fridley J D. Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities. Oecologia, 2002, 132(2): 271-277. DOI:10.1007/s00442-002-0965-x |