 2020, Vol. 40
2020, Vol. 40文章信息
- 李涛, 彭丽萍, 师庆东, 麦尔哈巴·尼加提, 戴岳
- Li T, Peng L P, Shi Q D, Marhaba·Nijat, Dai Y
- 新疆准噶尔盆地不同径级梭梭和白梭梭的水分来源
- Water sources of Haloxylon ammodendron and H. persicum of different diameter classes in Junggar Basin, Xinjiang
- 生态学报. 2020, 40(6): 2099-2110
- Acta Ecologica Sinica. 2020, 40(6): 2099-2110
- http://dx.doi.org/10.5846/stxb201809071913
-
文章历史
- 收稿日期: 2018-09-07
- 修订日期: 2019-12-11





2. 新疆绿洲生态重点实验室, 乌鲁木齐 830046
2. Key Laboratory of Xinjiang Oasis Ecology, Xinjiang University, Urumqi 830046, China
在干旱区, 水是主要限制性因子之一, 水分的变化直接影响植物的生长状况和生境格局[1], 因此, 确定植物水分来源及其利用方式对干旱区植物的恢复及了解植物对干旱环境的适应性和耐受性有重要意义。传统的根系调查法存在费时耗力, 有一定的误差, 同时难以量化植物对水源的利用比例等问题, 而稳定同位素技术克服了这些难题, 其具有较高的准确性和灵敏度[2], 为了解植物水分来源及利用比例提供了新的方法。
梭梭(Haloxylon ammdendron)和白梭梭(H. persicum)均为藜科(Chenopodiaceae)梭梭属(Haloxylon)多年生小乔木或灌木, 主要分布在中亚干旱少雨的荒漠地区[3]。梭梭和白梭梭作为优良的生物固沙植物, 对沙漠高温、干旱和盐碱都具有很强的适应性和耐受性[2, 4], 因此成为生态修复中的首选植物。新疆准噶尔盆地的梭梭占全国荒漠植被总面积的68%[5-6]而白梭梭在我国仅分布于新疆[7]。前人对梭梭和白梭梭做了许多方面的研究[8-14], 但主要集中于对梭梭和白梭梭结构形态和生理生态方面的研究, 对不同径级梭梭和白梭梭的水分来源及利用方式的研究较少。在本研究区, 梭梭多分布于沙丘底部或沙丘中部及丘间低地, 白梭梭主要分布在沙丘顶部, 研究不同径级的梭梭和白梭梭的水分来源对于认识梭梭和白梭梭的分布具有一定的探索意义。
不同林龄的植物主要水分来源不同。刘树宝等[15]研究发现, 胡杨幼苗的主要吸水层位是地下30—50 cm, 成熟木与过熟木的主要吸水层位都接近地下水;赵西宁等[16]等研究发现不同树龄的枣树, 在开花坐果期, 随着树龄的增加, 枣树对浅层土壤水利用比例减少, 对深层土壤水的利用比例增加。植物的水分利用方式不仅与潜在水源有关还与植物种类与树龄有关[17], 因此, 本研究利用MixSIAR模型和植物平均吸水深度模型探究不同林龄梭梭和白梭梭的水分利用方式, 以揭示梭梭和白梭梭对干旱环境的适应性, 为荒漠区植被恢复与保护提供参考依据。
1 材料与方法 1.1 研究区概况研究区位于新疆准东木垒县乔仁乡辖区境内(44°12′42″N, 90°03′12.70″E, 海拔:686.8 m), 该地深处大陆腹地, 属大陆干旱荒漠气候, 年温差和昼夜温差大, 四季分明。5—8月为夏季, 高温炎热, 白天气温常在40℃以上, 11月至次年2月为冬季, 绝对最低气温为-49.8℃。年平均降水量106 mm, 年蒸发量1202—2382 mm, 5—8月偶有雷阵雨, 冬季积雪稀少。由于受多风、降雨量小和蒸发量大等气候因素影响, 研究区植物组成简单, 物种单一, 区内建群植物是由超旱生、旱生的半乔木、灌木、小半灌木以及旱生的一年生草本、多年生草本和中生的短命植物等组成, 主要组成植物有梭梭、白梭梭、盐生假木贼(Anabasis salsa)、小蓬(Nanophyton erinaceum)、短叶假木贼(Anabasis brevifolia)、无叶假木贼(Anabasis aphylla)、泡果白刺(Nitraria sphaerocarpa)、角果藜(Ceratocarpus arenarius)、盐生草(Halogeton glomeratus)、盐爪爪(Kalidium foliatum)、虎尾草(Chloris virgata)等。其中, 梭梭多分布于丘间低地或沙丘中部, 白梭梭主要分布在沙丘顶部。
1.2 研究方法 1.2.1 样品采集(1) 植物样品采集:实验于2017年8月开展, 在研究区内分别选取长势良好且健康的不同径级的梭梭和白梭梭, 每种径级选取5株形态特征相近的植株, 剪取非绿色的栓化小枝(直径0.1—0.3 cm, 长3—5 cm), 去皮后迅速装入特制的玻璃瓶中并用封口膜密封, 然后放入随身携带的冰盒中, 带回实验室冷冻保存(低于-20℃), 用于测定δ18Ο值。采样时间为2017年8月中下旬, 采样的前后几天内均无降雨产生也无凝结水生成, 且降水、地表径流等“初始”水源只有转化成土壤水后才能被植物吸收[15], 研究区发生降水事件较少, 因此本文不考虑降水的影响。由于梭梭和白梭梭在年龄划分的过程中, 存在多年轮且地上植株增长不规则等问题[18], 因此本文运用径级结构代替年龄结构[19], 分别将梭梭和白梭梭划分成四个径级:Ⅰ(d≤1 cm)、Ⅱ(1 cm < d≤5 cm)、Ⅲ(5 cm < d≤10 cm)、Ⅳ(10 cm < d)。各径级梭梭和白梭梭的基本特征分别见表 1、2。
| 径级 Diameter class |
株高 Height/cm |
冠幅 Canopy width/cm |
基径 Basal diameter/cm |
土壤质地 Soil texture |
生活型 Life form |
生境 Habitat |
| Ⅰ | 29.0±15.04 | 24.2×23 | 0.94±0.26 | 沙土 | 旱生小乔木 | 丘间低地 |
| Ⅱ | 118.2±20.29 | 109.2×107.4 | 3.18±1.07 | |||
| Ⅲ | 184.4±22.37 | 196.0×192.8 | 6.88±1.26 | |||
| Ⅳ | 271.2±38.48 | 257.0×263.8 | 11.92±0.91 | |||
| 数值为平均值±标准差, n=5;Ⅰ, 梭梭径级(d):d≤1 cm; Ⅱ, 梭梭径级(d):1 cm < d≤5 cm;Ⅲ, 梭梭径级(d):5 cm < d≤10 cm;Ⅳ, 梭梭径级(d):10 cm < d | ||||||
| 径级 Diameter class |
株高 Height/cm |
冠幅 Canopy width/cm |
基径 Basal diameter/cm |
土壤质地 Soil texture |
生活型 Life form |
生境 Habitat |
| Ⅰ | 29.40±10.55 | 17.60×15.4 | 0.680±0.13 | 沙土 | 旱生小乔木 | 沙丘顶部 |
| Ⅱ | 131.80±20.19 | 83.00×85.4 | 2.760±1.16 | |||
| Ⅲ | 288.25±25.06 | 203.80×195.8 | 7.675±1.40 | |||
| Ⅳ | 321.00±44.92 | 314.20×314.6 | 15.120±2.58 | |||
| 数值为平均值±标准差, n=5; Ⅰ, 白梭梭径级(d):d≤1 cm; Ⅱ, 白梭梭径级(d):1 cm < d≤5 cm;Ⅲ, 白梭梭径级(d):5 cm < d≤10 cm;Ⅳ, 白梭梭径级(d):10 cm < d | ||||||
(2) 土壤样品采集:在采集植物样品附近用土钻采集土壤样品。每20 cm土层取一个样, 每个土样取3个重复, 直至200 cm, 取样时间同植物的取样时间一致。将所取的每层土样分成两份, 一份迅速装入样品瓶中, 用封口膜封口后装入随身携带的冰盒中, 带回实验室用于测定δ18Ο值, 另一份装入铝盒, 带回实验室用烘干法测定土壤质量含水量。
(3) 地下水样品采集:地下水样品取自离研究区最近的林业管理站。由于地下水埋深较深(约18 m), 因此取林业管理站抽水泵抽出的井水作为地下水样品测定其δ18O值。
1.2.2 同位素分析植物和土壤样品中的水分在中国科学院新疆生态与地理研究所荒漠与绿洲生态国家重点实验室采用低温真空蒸馏抽提法进行抽提, 将抽提出的植物小枝木质部水(简称木质部水)和土壤水分别装入玻璃样品瓶中, 利用基于稳定同位素红外光谱技术的液态水同位素分析仪(LWIA, DLT-100, Los Gatos Research, Mountain View, USA)测量样品的氧稳定同位素比率。氧同位素比率(δ)值的计算公式如下:
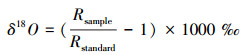
|
(1) |
式中, O代表需要测定的氧同位素值;Rsample代表样品中氧元素的重轻同位素丰度之比;Rstandard代表国际通用标准物中氧元素的重轻同位素丰度之比。由于利用低温真空蒸馏抽提技术获得的茎秆水中含有甲醇和乙醇类有机污染物, 易造成18Ο的同位素测量值超过仪器精度, 使其与实际值有一定的偏差。因此结合Los Gatos公司的光谱分析软件确定甲醇(narrow-band metric, NB)和乙醇(broad-band metric, BB)类物质污染程度的光谱度量值, 对同位素光谱污染进行校正[20], 以得到较准确的同位素值。
为了解不同径级梭梭和白梭梭的吸水层位, 将各径级梭梭和白梭梭木质部水, 各层土壤水以及地下水的δ18O值进行比较, 得出木质部水与各潜在水源的交叉点, 该垂直线的交叉点即是植物吸水的主要深度[21-22]。利用这种图形推断的方法可以对植物的吸水层位进行初步的判断。
1.2.3 MixSIAR模型MixSIAR模型是一个在R语言软件中运行的贝叶斯混合模型, 可以用来确定植物对不同潜在水源的利用比例, 具有可以同时处理任何数量的同位素值、同时分析多个潜在源、运行速度快、可以综合考虑各同位素值之间的差异等特性。
贝叶斯同位素混合模型MixSIAR估计了植物对潜在水源的利用比例, 其中包含了与多种来源相关的大量因子参数以及它们的不确定度, 使模型具有更高的准确性和严谨度[23]。模型运行时, 分别将不同径级的梭梭和白梭梭木质部同位素值作为混合数据输入MixSIAR模型中, 不同土壤层(0—20, 20—40, 40—60, 60—80, 80—100, 100—120, 120—140, 140—160, 160—180, 180—200 cm)及地下水同位素值的平均值和标准差作为源数据输入MixSIAR模型中, 由于植物在吸水过程中不发生同位素分馏现象[24], 因此将各潜在源的平均值和标准差均设为0, 作为差异数据输入MixSIAR模型中, 蒙特卡罗马尔科夫链运行长度设置为“长”, 误差项选择设置为“残差”[25], 然后运行模型。
为了方便对比与分析, 根据土壤水δ18O值的变化, 将潜在水源划分为四个:浅层土壤水:0—60 cm(土壤水δ18O值随土层深度的增加变化较大);中层土壤水:60—140 cm(土壤水δ18O值随土层深度的增加而减小);深层土壤水:140—200 cm(土壤水δ18O值随土层深度的增加趋于稳定)及地下水。
1.2.4 植物平均吸水深度模型不同径级的梭梭和白梭梭对土壤水的利用深度用植物平均吸水深度模型计算[26]。该模型是基于MATLAB软件, 以同位素质量守恒为前提条件, 运用插值计算出每厘米土壤的同位素值从而得出植物对土壤水的平均吸水深度。该模型运行前有三个假设:(1)在任何时间内, 植物都可以吸收0—50 cm处的水分; (2)在整个50 cm的部分, 植物的吸水服从正态分布(公式2);(3)植物不从两个不同的土壤剖面区域获取水分。

|
(2) |
式中, ni是植物在深度Y处所吸收水分的比例, μ是植物在土壤中吸水的平均深度, σ是标准偏差值为8.33。
模型的具体算法是:分别依次输入所采集的土壤层次, 每层土壤的同位素值, 植物木质部的同位素值, 然后输入植物吸收土壤水的标准偏差(8.33), 全部输入后, 模型开始由1计算, 土壤深度的计算间隔为1cm, 从而可以得出每厘米土壤水的同位素的贡献率。模型可以算出植物具体吸收哪一深度的土壤水分[27]。
1.2.5 数据分析由于梭梭和白梭梭是超旱生植物且梭梭属于聚盐性植物, 而旱生植物和盐生植物的根系在吸收水分的过程中会发生氢同位素分馏[28], 因此本研究采用稳定氧同位素进行分析。运用SPSS 19.0对数据进行分析, 使用单因素方差分析(one-way ANOVA)比较土壤剖面不同层次间土壤质量含水量之间的差异显著性及不同径级梭梭和白梭梭木质部水和土壤水δ18O值间的差异显著性, 运用MixSIAR模型分析潜在水源对植物的贡献比例, 植物平均吸水深度模型计算梭梭和白梭梭的平均吸水深度, 运用Origin8.0软件进行图形绘制。
研究区的气象数据来源于天气查询网站(网址:www.tianqi.911cha.com)。
2 结果分析 2.1 研究区降水和气温变化研究区在采样期间(2018年8月20—25日)未发生任何降水事件(图 1), 整个采样月研究区日平均气温呈波动性变化, 日平均气温最高31℃, 最低19℃, 总降水量为21.3 mm, 单次降水量均小于10 mm, 最大降水量为6.7 mm。

|
| 图 1 研究区采样期间的日均气温和日降水 Fig. 1 Daily mean temperature and daily precipitation in the research area during sampling period |
研究区位于荒漠地带, 不同径级的梭梭多生长于丘间低地, 白梭梭多生长于沙丘上。沙丘顶部和丘间低地土壤质量含水量总体较低, 且大致表现出随土层深度的增加而增加的趋势(图 2)。在丘间低地, 0—60 cm土壤质量含水量呈波动性变化, 变化范围在0.50%—0.66%, 土壤含水量在40 cm处最小, 为(0.26±0.03)%;60—140 cm土壤质量含水量变化不大;140—200 cm土壤质量含水量明显增加, 从(1.49±1.32)%增加到(2.38±1.63)%, 在200 cm处达到最大(图 2)。在沙丘顶部, 0—100 cm土壤质量含水量随土层深度的增加而增加, 从(0.14±0.17)%增加到(1.72±0.99)%, 在表层最小, 为(0.14±0.11)%;100—200 cm土壤质量含水量呈波动性变化, 变化范围为1.29%—2.14%, 在180 cm处达到最大, 为(2.14±0.59)%(图 2)。

|
| 图 2 丘间低地和沙丘顶部不同深度土层土壤质量含水量的变化(平均值±标准差, n=3) Fig. 2 Vertical profiles of gravimetricsoil water content at inter-dune and dune crest (Mean±SD, n=3) |
梭梭生长的丘间低地土壤水δ18O值总体变化范围在-0.66‰—-14.22‰之间, 在土壤表层表现出重同位素严重富集的现象, 且其值随土壤深度的增加而逐渐减小(图 3)。土壤水δ18O值在0—60 cm土层减小的幅度较大, 从(-0.66±1.96)‰减小到(-8.54±0.51)‰;在60—140 cm土层减小的幅度逐渐趋于平缓, 变化范围在-10.72‰—-13.36‰;在140—200 cm土层其变化趋于稳定, 其值在-14‰左右。白梭梭生长的沙丘顶部土壤水的δ18O值总体变化范围在0.55‰—-13.24‰之间, 在土壤表层表现出重同位素富集的现象, 整体呈随土壤深度增加而减小的趋势(图 3)。土壤水δ18O值在0—80 cm土层减小幅度较大, 从(0.55±1.55)‰减小到(-9.41±2.07)‰;在80—140 cm土层减小的幅度逐渐减小, 变化范围在-10.06‰—-11.92‰;在140—200 cm土层其值稳定在-12.79‰左右。

|
| 图 3 不同径级梭梭和白梭梭的木质部水、土壤水和地下水的δ18O值比较(平均值±标准差, n=3) Fig. 3 Comparison of δ18O values of xylem water, soil water and groundwaterbetween different diameter classes of Haloxylon ammodendron and H. persicum (Mean±SD, n=3) Ⅰ, 梭梭/白梭梭径级(d):d≤1 cm; Ⅱ, 梭梭/白梭梭径级(d):1 cm < d≤5 cm;Ⅲ, 梭梭/白梭梭径级(d):5 cm < d≤10 cm;Ⅳ, 梭梭/白梭梭径级(d):d>10 cm |
四个径级梭梭的δ18O值总体变化范围在-8.37‰—-13.95‰之间(图 3), 不同径级间梭梭木质部δ18O值差异显著(P < 0.01)。径级Ⅰ的梭梭木质部水δ18O值为(-8.37±0.64)‰, 与40—60 cm处土壤水的δ18O值(-8.54±0.51)‰接近(P=0.77), 因此可以推断径级Ⅰ的梭梭主要利用40—60 cm土层的土壤水;径级Ⅱ的梭梭木质部水δ18O值为(-13.95±1.19)‰, 与140—160 cm土层的土壤水的δ18O值(-14.42±0.087)‰接近(P=0.78), 因此初步推断径级Ⅱ的梭梭主要利用140—160 cm土层的土壤水;径级Ⅲ的梭梭木质部水δ18O值为(-13.77±1.20)‰, 与140—160 cm处土壤水的δ18O值接近(P=0.79), 因此推断径级Ⅲ的梭梭主要利用140—160 cm土层深度的土壤水。径级Ⅳ的梭梭木质部水δ18O值为(-13.07±1.67)‰, 与100—120 cm处土壤水的δ18O值(-13.48±1.93)‰接近(P=0.79), 因此推断径级Ⅳ的梭梭主要利用100—120 cm土层的土壤水。
四个径级的白梭梭δ18O值总体变化范围在-6.96‰—-13.18‰之间(图 3), 不同径级间白梭梭木质部δ18O值差异显著(P < 0.01)。径级Ⅰ的白梭梭木质部水δ18O值为(-6.96±1.46)‰, 与40—60 cm处土壤水的δ18O值(-6.37±0.18)‰接近(P=0.67), 因此可以推断径级Ⅰ的白梭梭主要利用40—60 cm土层的土壤水;径级Ⅱ的白梭梭木质部水δ18O值为(-10.70±0.75)‰, 与100—120 cm处土壤水的δ18O值(-11.81±0.88)‰接近(P=0.68), 因此可以推断径级Ⅱ的白梭梭主要利用100—120 cm处的土壤水;径级Ⅲ的白梭梭木质部水δ18O值为(-12.73±0.44)‰, 与160—180 cm处土壤水的δ18O值(-12.48±0.68)‰接近(P=0.58), 因此可以推断径级Ⅲ的白梭梭主要利用160—180 cm土层的土壤水;径级Ⅳ的白梭梭木质部水δ18O值为(-13.18±0.61)‰, 与180—200 cm处土壤水的δ18O值(-13.25±0.26)‰接近(P=0.84), 因此可以推断径级Ⅳ的白梭梭主要利用180—200 cm土层的土壤水。
2.4 梭梭和白梭梭对各潜在水源的利用比例 2.4.1 各径级梭梭和白梭梭对潜在水源的利用比例运用MixSIAR模型分析得出不同径级的梭梭对不同层次的土壤水和地下水的利用比例各不相同(图 4)。径级Ⅰ的梭梭对各潜在水源的利用比例大小为:20 cm>40 cm>60 cm>80 cm>100 cm>120 cm>140 cm>180 cm>200 cm>160 cm>地下水;径级Ⅱ的梭梭对各潜在水源的利用比例大小依次为:180 cm>160 cm>200 cm>120 cm>140 cm>100 cm>地下水>80 cm>60 cm>40 cm>20 cm;径级Ⅲ的梭梭对各潜在水源的利用比例大小依次为:180 cm>160 cm>200 cm>120 cm>140 cm>100 cm>地下水>80 cm>60 cm>40 cm>20 cm;径级Ⅳ的梭梭对各潜在水源的利用比例大小依次为:120 cm>180 cm>160 cm>200 cm>140 cm>100 cm>地下水>80 cm>60 cm>40 cm>20 cm。径级Ⅱ和径级Ⅲ的梭梭表现出一致的吸水模式, 径级Ⅰ和径级Ⅳ的梭梭对潜在水源的利用有较大差异。

|
| 图 4 不同径级的梭梭对潜在水源的利用比例(平均值±标准差, n=3) Fig. 4 Proportions of feasible water sources for Haloxylon ammodendron at different diameter classes (Mean±SD, n=3) |
运用MixSIAR模型分析得出不同径级的白梭梭对不同层次的土壤水和地下水利用比例各不相同(图 5)。径级Ⅰ的白梭梭对各潜在水源的利用比例大小依次为:20 cm>40 cm>60 cm>80 cm>100 cm>地下水>140 cm>180 cm>120 cm>160 cm>200 cm;径级Ⅱ的白梭梭对各潜在水源的利用比例大小依次为:160 cm>140 cm>180 cm>120 cm>200 cm>地下水>100 cm>80 cm>60 cm>40 cm>20 cm;径级Ⅲ的白梭梭对各潜在水源的利用比例大小依次为:160 cm>200 cm>180 cm>140 cm>120 cm>地下水>100 cm>80 cm>60 cm>40 cm>20 cm;径级Ⅳ的白梭梭对各潜在水源的利用比例大小依次为:160 cm>200 cm>180 cm>140 cm>120 cm>地下水>100 cm>80 cm>60 cm>40 cm>20 cm。径级Ⅰ的白梭梭对浅层土壤水的利用较多, 径级Ⅱ的白梭梭对中层土壤水和深层土壤水的利用较为平均, 径级Ⅲ和径级Ⅳ的白梭梭对深层土壤水利用较多。

|
| 图 5 不同径级的白梭梭对潜在水源的利用比例(平均值±标准差, n=3) Fig. 5 Proportions of feasible water sources for Haloxylon persicum at different diameter classes (Mean±SD, n=3) |
各径级梭梭都主要利用土壤水, 对地下水的利用较少, 但它们所利用的土壤水深度和比例有所不同(图 6)。径级Ⅰ的梭梭主要利用浅层土壤水, 利用比例为(45.4±10.9)%, 对其他各水源的利用大小依次为:中层土壤水(28.2±6.4)%>深层土壤水(18.2±5.5)%>地下水(8±7.2)%;径级Ⅱ的梭梭主要利用深层土壤水, 利用比例为(40.7±11.5)%, 对其他各水源的利用大小依次为:中层土壤水(39.3±9.3)%>浅层土壤水(12.5±4.5)%>地下水(7.5±7.0)%;径级Ⅲ的梭梭主要利用深层土壤水, 利用比例为(41.7±11.8)%, 对其他各水源的利用大小依次为:中层土壤水(39.4±9.5)%>浅层土壤水(11.9±4.3)%>地下水(7.1±6.6)%;径级Ⅳ的梭梭主要利用中层土壤水, 利用比例为(42±9.2)%, 对其他各水源的利用大小依次为:深层土壤水(36.6±10.3)%>浅层土壤水(15±4.9)%>地下水(8.4±7.6)%。随着梭梭径级的增加, 其吸水方式表现出由浅层到深层再到中层的变化。

|
| 图 6 不同径级的梭梭、白梭梭对潜在水源的利用比例对比(平均值±标准差, n=3) Fig. 6 Comparison of proportions of feasible water sources for Haloxylon ammodendron and H. persicum at different diameter classes(Mean±SD, n=3) |
各径级的白梭梭都主要利用土壤水, 对地下水利用的比例较小, 对不同层次的土壤水利用比例不同((图 6)。径级Ⅰ的白梭梭主要利用浅层土壤水, 利用比例为(40.5±10.0)%, 对其他各水源的利用大小依次为:中层土壤水(30.7±7.0)%>深层土壤水(21.2±6.3)%>地下水(7.7±7.0)%;径级Ⅱ的白梭梭主要利用中层土壤水, 利用比例为(40.2±9.08)%, 对其他各水源的利用大小依次为:深层土壤水(35.4±5.8)%>浅层土壤水(13.8.3±4.2)%>地下水(10.4±9.5)%;径级Ⅲ的白梭梭主要利用深层土壤水, 利用比例为(40.0±12.2)%, 对其他各水源的利用大小依次为:中层土壤水(36.9±8.8)%>浅层土壤水(14.0±5.12)%>地下水(9.1±8.3)%;径级Ⅳ的白梭梭主要利用深层土壤水, 利用比例为(52.4±15.0)%, 对其他各水源的利用大小依次为:中层土壤水(32.7±8.3)%>浅层土壤水(7.5±3.14)%>地下水(7.3±7.2)%。不同径级的白梭梭随着径级的增加, 表现出对深层土壤水的利用比例随之增加的现象。
2.5 不同径级梭梭和白梭梭的平均吸水深度不同径级的梭梭平均吸水深度略有不同, Ⅰ—Ⅲ径级的梭梭随径级的增加其平均吸水深度不断增加, 而Ⅳ径级的梭梭其平均吸水深度稍有减小(图 7)。径级Ⅰ的梭梭平均吸水深度为(59.72±4.36)cm, 径级Ⅱ的梭梭平均吸水深度为(144.94±37.23)cm, 径级Ⅲ的梭梭平均吸水深度为(156.97±46.37)cm, 径级Ⅳ的梭梭平均吸水深度为(132.84±46.65)cm。

|
| 图 7 不同径级的梭梭和白梭梭的平均吸水深度(平均值±标准差, n=5) Fig. 7 Mean depths of water uptake by Haloxylon ammodendron and H. persicum at different diameter classes (Mean±SD, n=5) |
不同径级的白梭梭平均吸水深度随径级的增加而增加, 其径级越大越趋向于利用深层土壤水(图 7)。径级Ⅰ的白梭梭平均吸水深度为(64.11±6.92)cm, 径级Ⅱ的白梭梭平均吸水深度为(105.28±12.88)cm, 径级Ⅲ的白梭梭平均吸水深度为(172.36±23.67)cm, 径级Ⅳ的白梭梭平均吸水深度为(194.45±0.89)cm。
3 讨论 3.1 土壤质量含水量及其同位素值变化土壤质量含水量无论是在沙丘顶部还是丘间低地都随土层深度的增加而增加, 但总体呈较低水平(图 2)。主要是因为当地的土壤受降水的补给较少, 且气候炎热, 表层土壤水分蒸发量大, 且沙土土质疏松, 透气性好但保水性差[29]。土壤含水量随土层深度的增加呈波动变化, 可能与植物根系分布与土壤物理结构有关。土壤水δ18O值在表层都达到最大值, 随土层深度的增加而减小(图 3)。这是由于该地蒸发量较大, 土壤水δ18O值易受到蒸发分馏作用的影响, 表层比下层土壤水更易富集重同位素[30-31]。
3.2 不同径级的梭梭和白梭梭的吸水深度植物对土壤水的利用主要取决于植物根系的分布。分布在表层土壤中的根系能够吸收降水补充的土壤水, 而分布在深层土壤中的根系主要吸收的是由冬春季节的降水或者地下水补充的土壤水[32]。由于梭梭根具有趋水性, 当干旱时, 其向下生长以吸取深层土壤水分[33]。对于本研究区, 由于极少发生降水且地下水位较深, 因此, 梭梭主要利用的水源是土壤水。径级Ⅰ的梭梭其主根系经挖掘实测长度范围在30—60 cm, 因此径级Ⅰ的梭梭主要利用0—60 cm的浅层土壤水, 这与傅思华等[34]的研究结果一致;径级Ⅱ—Ⅲ的梭梭, 由于其根系较为庞大, 垂直根系最大可以达到5 m左右[33], 因此, 推断其主要利用深层土壤水, 这与图形推断法、MixSIAR模型及平均吸水深度模型计算出的结果及与朱雅娟等[35]得出的研究结论一致。径级Ⅳ的梭梭根据图形推断法和MixSIAR模型及平均吸水深度模型计算出的结果推断其主要利用中层土壤水。在干旱环境中, 梭梭会趋向于利用较深层的土壤水, 但随着梭梭年龄的增加, 其水源利用更加灵活, 趋向于利用中层土壤水, 这与赵西宁[16]所研究的结果期不同树龄的枣树水分利用情况及朱雅娟等[36]所研究的秋季不同林龄梭梭水分利用状况是一致的, 这可能与梭梭根系的分布, 特别是细根的分布[36]及梭梭所处的生境有关。梭梭根系的特点使之较其他植物更容易获取地下水和广泛吸收土壤水不饱和区域的水分[33], 从而更好的适应干旱低矮小沙丘及丘间低地的生态环境。
不同林龄和大小的植物因所处的地表、地下以及微生物环境存在差异, 水分利用模式也会发生变化[37]。胡杨(Populus euphratica)随着树龄的增加其会利用更深层的土壤水[15], 辽西半干旱地区大扁杏(Prunus armeniaca)随着树龄的增加利用不同深度的土壤水[38]。本研究中, 白梭梭随年龄的增加, 趋向于利用更深层的土壤水(图 7)。径级Ⅰ的白梭梭经实测其主根系根长在20—40 cm, 因此, 主要利用0—60 cm的浅层土壤水, 这与图形推断法、MixSIAR模型及平均吸水深度模型推断出的结果一致;径级Ⅱ—Ⅳ的白梭梭, 根据本文的研究结果推断其为深根系植物, 且随着白梭梭年龄的增加, 其根系会不断向下生长, 以寻求更稳定的水源, 因此径级Ⅱ的白梭梭主要利用中层土壤水, 径级Ⅲ—Ⅳ的白梭梭主要利用深层土壤水。白梭梭在本研究区水分来源主要是土壤水, 这与李晖[39]和Wu等[40]的研究结果一致。由于当地的地下水埋深较深, 且白梭梭生长于沙丘顶部, 使之更难利用到地下水, 因此, 白梭梭以土壤水作为稳定的水源。
梭梭和白梭梭通过不同的水分利用方式, 合理的利用干旱区有限的水资源。傅思华等[41]研究表明梭梭对各潜在水源利用比例存在季节性变化, 在5月份主要利用融雪补给的水源和地下水, 在8月份主要利用深层土壤水及地下水。Dai等[42]研究表明当春季浅层土壤水丰富时梭梭主要利用浅层土壤水, 而白梭梭主要利用中层土壤水, 当夏季干旱时, 梭梭主要利用地下水而白梭梭主要利用深层土壤水。Wu等[40]研究表明在极端干旱的时期, 梭梭主要利用近地下水层的土壤水, 而白梭梭主要利用深层土壤水, 随着地下水位埋深的增加梭梭和白梭梭的吸水深度也随之增加。在本研究中, 地下水埋深较深(约18 m), 且采样时间在是新疆准东地区最干旱的时期(8月份), 不同径级的梭梭和白梭梭对土壤水的吸水层位和利用比例略有不同, 这可能与梭梭和白梭梭根系分布、土壤含水量差异及所处生境不同有关, 其分布更多是对丘间、丘顶环境的适应。不同径级的梭梭和白梭梭水分利用方式不同, 可能导致梭梭和白梭梭的生态位分化, 以降低它们对水分的竞争强度, 促进共存。
4 结论本研究中, 梭梭和白梭梭都将土壤水作为稳定水源, 不同径级的梭梭和白梭梭对土壤水的利用略有不同。生长在丘间低地的梭梭对水源的利用更加灵活, 随着径级的增长趋向于利用稍浅层的土壤水。生长在沙丘顶部的白梭梭随着径级的增长则趋向于利用更深层的土壤水。不同径级的梭梭和白梭梭选择利用不同深度的土壤水, 以降低它们对水分的竞争强度。
| [1] |
丁丹, 贾文雄, 马兴刚, 王洁. 祁连山亚高山灌丛优势植物水分来源. 生态学报, 2018, 38(4): 1348-1356. |
| [2] |
徐庆, 冀春雷, 王海英, 李旸. 氢氧碳稳定同位素在植物水分利用策略研究中的应用. 世界林业研究, 2009, 22(4): 41-46. |
| [3] |
沈亮, 陈君, 刘赛, 徐荣, 徐常青, 刘同宁. 脱水胁迫和光合日变化对梭梭和白梭梭叶绿素荧光参数的影响. 应用生态学报, 2015, 26(8): 2321-2328. |
| [4] |
王丽娟, 孙栋元, 赵成义, 李菊艳, 盛钰. 准噶尔盆地梭梭、白梭梭植物构型特征. 生态学报, 2011, 31(17): 4952-4960. |
| [5] |
王春玲, 郭泉水, 谭德远, 史作民, 马超. 准噶尔盆地东南缘不同生境条件下梭梭群落结构特征研究. 应用生态学报, 2005, 16(7): 1224-1229. DOI:10.3321/j.issn:1001-9332.2005.07.008 |
| [6] |
黄培祐, 李启剑, 袁勤芬. 准噶尔盆地南缘梭梭群落对气候变化的响应. 生态学报, 2008, 28(12): 6051-6059. DOI:10.3321/j.issn:1000-0933.2008.12.033 |
| [7] |
中国植被编辑委员会. 中国植被. 北京: 科学出版社, 1983: 589.
|
| [8] |
戴岳, 郑新军, 唐立松, 李彦. 古尔班通古特沙漠南缘梭梭水分利用动态. 植物生态学报, 2014, 38(11): 1214-1225. |
| [9] |
谢江波, 彭历芝, 李艳明, 李彦. 两种梭梭属幼苗在土壤N和P梯度下的竞争能力研究. 干旱区地理, 2018, 41(1): 83-91. |
| [10] |
宋于洋.古尔班通古特沙漠梭梭种群动态与持续发育[D].咸阳: 西北农林科技大学, 2011.
|
| [11] |
王戌梅.梭梭的遗传变异性研究[D].沈阳: 沈阳药科大学, 2009.
|
| [12] |
张维, 赵文勤, 谢双全, 张莉, 吕新华, 贺亚玲. 模拟降水和氮沉降对准噶尔盆地南缘梭梭光合生理的影响. 中国生态农业学报, 2018, 26(1): 106-115. |
| [13] |
李从娟, 马健, 李彦, 李惠. 梭梭和白梭梭主根周围土壤养分的梯度分布. 中国沙漠, 2011, 31(5): 1174-1180. |
| [14] |
陈虞超, 李苗, 吴明朝, 宋玉霞. 梭梭属两种植物的根结构和成分. 植物生理学报, 2013, 49(11): 1273-1276. |
| [15] |
刘树宝, 陈亚宁, 李卫红, 陈亚鹏, 任志国. 黑河下游不同林龄胡杨水分来源的D、18O同位素示踪. 干旱区地理, 2014, 37(5): 988-995. |
| [16] |
赵西宁, 李楠, 高晓东, 霍高鹏, 潘燕辉. 基于18O示踪的不同树龄枣树土壤水分利用特征分析. 农业工程学报, 2018, 34(3): 135-142. |
| [17] |
朱林, 许兴, 毛桂莲. 宁夏平原北部地下水埋深浅地区不同灌木的水分来源. 植物生态学报, 2012, 36(7): 618-628. |
| [18] |
李建贵, 宁虎森, 刘斌. 梭梭种群性状结构与空间分布格局的初步研究. 新疆农业大学学报, 2003, 26(3): 51-54. DOI:10.3969/j.issn.1007-8614.2003.03.013 |
| [19] |
梁士楚, 蒋潇潇, 李峰. 广西英罗港红树植物木榄种群年龄结构的研究. 海洋学研究, 2008, 26(4): 35-40. DOI:10.3969/j.issn.1001-909X.2008.04.005 |
| [20] |
孟宪菁, 温学发, 张心昱, 韩佳音, 孙晓敏, 李晓波. 有机物对红外光谱技术测定植物叶片和茎秆水δ18O和δD的影响. 中国生态农业学报, 2012, 20(10): 1359-1365. |
| [21] |
Brunel J P, Walker G R, Kennett-Smith A K. Field validation of isotopic procedures for determining sources of water used by plants in a semi-arid environment. Journal of Hydrology, 1995, 167(1/4): 351-368. |
| [22] |
Yang B, Wen X F, Sun X M. Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region. Agricultural andForest Meteorology, 2015, 201: 218-228. DOI:10.1016/j.agrformet.2014.11.020 |
| [23] |
Stock B C, Semmens B X.MixSIAR GUI User Manual.Version3.1.[2017-07-08]. https://github.com/brianstock/MixSIAR.
|
| [24] |
Brooks J R, Barnard H R, Coulombe R, McDonnell J J. Ecohydrologic separation of water between trees and streams in a Mediterranean climate. Nature Geoscience, 2010, 3(2): 100-104. DOI:10.1038/ngeo722 |
| [25] |
Wang J, Fu B J, Lu N, ZhangL. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Science of the Total Environment, 2017, 609: 27-37. DOI:10.1016/j.scitotenv.2017.07.133 |
| [26] |
Romero-Saltos H, da S. L. Sternberg L, Moreira M Z, NepstadD C. Rainfall exclusion in an eastern Amazonian forest alters soil water movement and depth of water uptake. American Journal of Botany, 2005, 92(3): 443-455. DOI:10.3732/ajb.92.3.443 |
| [27] |
刘树宝, 陈亚宁, 陈亚鹏, 邓海军, 方功焕. 基于稳定同位素技术的黑河下游不同林龄胡杨的吸水深度研究. 生态学报, 2016, 36(3): 729-739. |
| [28] |
Ellsworth P Z, Williams D G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant and Soil, 2007, 291(1/2): 93-107. |
| [29] |
伍光和, 王乃昂, 胡双熙, 田连恕, 张建明. 自然地理学(第四版). 北京: 高等教育出版社, 2008.
|
| [30] |
孙守家, 孟平, 张劲松, 万贤崇. 利用氘同位素研究太行山南麓枣树水分利用的季节性变化. 林业科学, 2011, 47(5): 46-53. |
| [31] |
Tang K L, Feng X H. The effect of soil hydrology on the oxygen and hydrogen isotopic compositions of plants'source water. Earth andPlanetary Science Letters, 2001, 185(3/4): 355-367. |
| [32] |
Chimner R A, Cooper D J. Using stable oxygen isotopes to quantify the water source used for transpiration by native shrubs in the San Luis Valley, Colorado U.S.A. Plant and Soil, 2004, 260(1/2): 225-236. DOI:10.1023/B:PLSO.0000030190.70085.e9 |
| [33] |
盛晋华, 乔永祥, 刘宏义, 翟志席, 郭玉海. 梭梭根系的研究. 草地学报, 2004, 12(2): 91-94. |
| [34] |
傅思华, 胡顺军, 杨涛, 朱海, 周天河. 古尔班通古特沙漠南缘地下水深埋区幼龄梭梭春夏季土壤水分利用动态. 水土保持学报, 2016, 30(5): 230-234. |
| [35] |
朱雅娟, 贾志清, 卢琦, 郝玉光, 张景波, 李磊, 綦艳林. 乌兰布和沙漠5种灌木的水分利用策略. 林业科学, 2010, 46(4): 15-21. DOI:10.3969/j.issn.1672-8246.2010.04.003 |
| [36] |
朱雅娟, 贾志清. 秋季巴丹吉林沙漠东南缘人工梭梭林水分来源. 林业科学, 2012, 48(8): 1-5. |
| [37] |
Kerhoulas L P, Kolb T E, Koch G W. Tree size, stand density, and the source of water used across seasons by ponderosa pine in northern Arizona. Forest Ecology and Management, 2013, 289: 425-433. DOI:10.1016/j.foreco.2012.10.036 |
| [38] |
刘丽颖, 殷有, 秦胜金, 周永斌, 卢国珍, 耿占礼. 辽西半干旱区不同林龄大扁杏人工林的水分利用特征. 水土保持学报, 2016, 30(6): 118-122. |
| [39] |
李晖.应用δD和δ18O确定古尔班通古特沙漠植物水分来源[D].乌鲁木齐: 中国科学院新疆生态与地理研究所, 2007.
|
| [40] |
Wu X, Zheng X J, Li Y, Xu G Q. Varying responses of two Haloxylon species to extreme drought and groundwater depth. Environmental and Experimental Botany, 2019, 158: 63-72. DOI:10.1016/j.envexpbot.2018.11.014 |
| [41] |
傅思华, 胡顺军, 李浩, 王泽锋. 古尔班通古特沙漠南缘梭梭(Haloxylonammodendron)群落优势植物水分来源. 中国沙漠, 2018, 38(5): 1024-1032. |
| [42] |
Dai Y, Zheng X J, Tang L S, Li Y. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylonspecies in the Gurbantonggut Desert. Plant and Soil, 2015, 389(1/2): 73-87. DOI:10.1007/s11104-014-2342-z |