 2020, Vol. 40
2020, Vol. 40文章信息
- 赵连春, 秦爱忠, 赵成章, 段凯祥, 王继伟, 文军
- ZHAO Lianchun, QIN Aizhong, ZHAO Chengzhang, DUAN Kaixiang, WANG Jiwei, WEN Jun
- 嘉峪关草湖湿地植物功能群组成及其性状对不同生境的响应
- Responses of composition and functional traits of plant functional groups to different habitats in Jiayuguan Caohu Wetland
- 生态学报. 2020, 40(3): 822-833
- Acta Ecologica Sinica. 2020, 40(3): 822-833
- http://dx.doi.org/10.5846/stxb201811052396
-
文章历史
- 收稿日期: 2018-11-05
- 网络出版日期: 2019-11-20





2. 甘肃省城乡规划设计研究院, 兰州 730000;
3. 西北师范大学地理与环境科学学院, 甘肃省湿地资源保护与产业发展工程研究中心, 兰州 730070
2. Institute of Urban and Rural Planning and Design of Gansu Province, Lanzhou 730000, China;
3. College of Geography and Environmental Science, Northwest Normal University, Research Center of wetland Resources Protection and Industrial Development Engineering of Gansu Province, Lanzhou 730070, China
植物功能群是对环境有相同响应和对主要生态系统过程有相似作用的物种的组合[1], 能够反映植被随环境的动态变化[2], 有效的解释生态系统的构建机制, 已成为研究生态系统的有效方法[3-4]。以功能群为基础的研究方法关注许多不同亲缘关系的种对同一生态环境压力的共同适应方式[5], 在一定程度上减少了群落复杂性, 突出了研究重点, 便于统计分析;而且可根据研究的背景、尺度以及拟解决的问题进行选择和调整, 因此适用对象广、灵活性高、针对性强;有利于从不同尺度上揭示出物种分布格局和物种对特定干扰的响应机制[6]。局域尺度上, 功能群主要受环境过滤和生态位的作用[7-9], 生境空间异质性对植物种群、功能群结构与动态等具有重要的影响[10]。因此, 从局域尺度上研究不同生境条件下植物功能群特征, 对于研究植物与环境关系、植物的环境适应机制、群落发育和群落构建机制等均具有重要意义。目前, 国内外学者利用功能群的研究方法, 在不同的尺度上从土地利用[11-12]、放牧[13-14]、火灾[15-16]、群落动态[17]、气候变化[1, 18]、生态系统服务功能[19]、谱系结构和演化[20-21]等方面对生态系统做了大量研究。尽管越来越多的研究集中到功能群, 但多是在区域或全球尺度上探讨气候变化对物种分布的影响, 从局域尺度探讨功能群分布与环境因子的关系报道不多[22], 尤其是从局域尺度上, 对生态脆弱区的生态系统如西北干旱区沙漠、戈壁、湿地等生态系统功能群的分布机制或功能群的环境适应机制进行的研究较少[23-24]。
嘉峪关草湖国家湿地公园位于嘉峪关市东北侧的荒漠区内, 地处荒漠绿洲过渡带, 属荒漠区较具特色的干旱区湿地之一, 拥有沼泽湿地、盐沼湿地、沙丘、戈壁等不同的景观, 是嘉峪关市与酒泉市两城市间的“城市之肾”, 对区域防风固沙、生态系统保护、生态环境净化等具有重要作用。近年来, 该区域因地处戈壁腹地, 干旱少雨, 区域生态水环境不断恶化, 土地沙化、荒漠化日益严重[25-26], 严重威胁着绿洲区域的生态安全。荒漠区植被如白刺(Nitraria tangutorum Bobr.)等, 可拦蓄和固定大量流沙, 形成灌丛沙堆, 固定绿洲外围的大量流沙, 对维护荒漠区生态环境有重要作用[27]。鉴于此, 本文基于植物功能性状的功能群分类方法, 对荒漠区不同生境条件下的功能群组成、功能性状对环境压力的响应等进行研究, 探讨植物功能群构建机制、分布机制和环境适应机制。试图明晰以下问题:(1)植物功能群组成是否受不同生境类型的影响?(2)植物功能性状是如何对生境做出响应的?本研究将为该区域群落构建机制的解释、干旱区植被变化的预测、生态保护、植被的修复与恢复等提供一定的理论依据。
1 材料和方法 1.1 研究区概况研究区位于甘肃省嘉峪关市新城镇草湖国家湿地公园内(图 1), 地理位置为39°54′10.55″─39°56′42.26″ N, 98°25′56.98″─98°29′25.23″ E, 海拔1426─1463 m, 年平均气温6.9 ℃, 年均降水量85.3 mm, 年均蒸发总量2148.8 mm, 年日照3000 h, 全年无霜期130 d, 昼夜温差变化较大, 年平均气温日较差16.67 ℃, 最大日较差29 ℃。风沙多, 冬冷夏热, 属荒漠型的中温带干旱大陆性气候。地处戈壁沙漠区, 自然环境恶劣, 生态环境脆弱, 有草本沼泽、盐沼、沙丘、戈壁4种地貌类型。生境不同, 土壤类型不同, 沼泽生境为沼泽土, 盐沼湿地为漠境盐碱土, 沙丘为风沙土, 戈壁为棕漠土。主要植物有红砂(Reaumuria songarica (Pall.) Maxim.)、白刺、黑果枸杞(Lycium ruthenicum Murray)、梭梭(Haloxylon ammodendron (C. A. Mey.) Bunge)、多枝柽柳(Tamarix ramosissima Ledeb.)、芦苇(Phragmites australis (Cav.) Trin. ex Steud.)、滨藜(Atriplex patens (Litv.) Iljin)、芨芨草(Achnatherum splendens (Trin.) Nevski)、碱蓬(Suaeda glauca (Bunge) Bunge)、嵩草(Kobresia myosuroides (Villars) Foiri)、圆穗苔草(Carex angarae Steud.)等。

|
| 图 1 研究区地理位置 Fig. 1 Location of study sites |
2018年8月植物生长茂盛期, 在嘉峪关草湖湿地开展野外调查取样工作。在草湖湿地上游水源地附近, 垂直河岸设置一条宽500 m, 长4000 m的样带(样带内包含沼泽、盐沼、沙丘、戈壁4种生境类型, 图 2), 在不同生境梯度内各选3个(5 m×5 m)典型样地, 每个样地内随机设3个(1 m×1 m)样方, 共27个样方, 记录各样方的地理位置及样方内植物名称、盖度、密度、高度等数据。通过文献查询、野外观察等方式, 获取该区域的植被功能性状数据。

|
| 图 2 研究样带的地形剖面图 Fig. 2 Topographic section of the transect |
在每个样方对角线各1/3处设3个点, 挖掘0.5 m深的土壤剖面, 用环刀间隔10 cm采取土样混合均匀, 土样编号、称鲜质量后带回实验室, 分析土壤指标。其中土壤水分采用烘干法;含盐量采用5:1水土比电导法[28]。
1.3 数据分析方法参照Mahdavi等人的研究方法, 确定调查物种的功能性状属性表(表 1), 然后对植物及其性状变量进行聚类分析(Ward′s method with relative Eucidean distance; PC-ORD)。为保证各功能群分布同所在生态系统相一致, 便于后面的分析研究, 手动确定分类水平, 使功能群分类结果同野外观察结果一致。
| 功能性状Functional traits | 属性Attributes |
| 植被性状Vegetative traits | |
| 生活型Life form | 一年生、地上芽、地面芽(包括地下芽)、高位芽 |
| 株高Plant height | 1=1—30 cm;2=31—60;3=61—100 cm;4= > 1 m |
| 克隆类型Clonality | 0=非克隆;1=克隆(0=no clonality; 1=clonal plant) |
| 植物策略类型plant strategy type | 竞争型(C);耐受型(S);杂草型(R);中间策略CS;CR;SR;CSR |
| 再生性状Regenerative traits | |
| 扩散方式Dispersal mode | 风播、动物传播、无明显扩散媒介(或自体扩散) |
| 授粉方式Pollination mode | 风媒、虫媒、动物媒、自我授粉 |
| 叶性状Leaf traits | |
| 光合途径Photosynthetic pathway | 0=C3;1=C4 |
| 叶或茎多肉Leaf or stem succulence | 0=非肉质;1=肉质(0=not succulent; 1= succulent) |
| 较少或较小的叶片Reduced leaves | 0=无明显叶退化;1=较少或较小的叶 |
| 具毛Hairiness | 0=无毛;1=有毛(0=glabrous; 1=hairy) |
| 多刺Thorniness | 0=无刺;1=有刺(0=no thorns; 1=thorny) |
| 被蜡Wax-coating | 0=无蜡;1=有蜡(0=no wax; 1= waxy) |
| 茎和根性状Stem and root traits | |
| 碳水化合物具增厚的根或增长的茎 Carbohydrate storage in thicked root and stem or long tap root |
0=无明确的贮藏器官;1=明确的贮藏器官 |
| 根茎型植物Rhizomatous plant | 0=非根茎型;1=根茎型(0=non-rhizomatous; 1=rhizomatous) |
对物种-样方矩阵进行DCA分析后, 将聚类分析得到的功能群作为变量投射到该散点图上, 得到功能群分布同物种分布的关系图。将样方-物种矩阵同物种-功能群矩阵相乘, 得到样方-功能群矩阵, 对该矩阵进行DCA分析, 得到功能群分布同生境的关系图;同理, 将性状-物种矩阵同物种-样方矩阵相乘, 得到性状-样地矩阵, 然后, 对性状-样地矩阵进行DCA分析, 得到功能性状分布同生境的关系图。用群落加权平均值法, 将各物种的相对盖度(百分比)归一化处理成相对数值后对性状值加权, 同分类性状进行比较, 得到各生境的综合性状[29]。
群落加权性状平均值公式(Community-weighted Mean Trait Value, CWM)[30]:
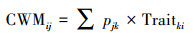
|
式中, pjk代表物种k在生境i中的相对盖度, Traitki代表物种k的性状i值的平均值。
相关矩阵、指数的计算在R语言中进行, DCA分析用CANOCO 4.5软件完成, 地理位置图、地形剖面图在ArcGIS 10.2软件中完成。
2 结果 2.1 不同生境的土壤特征随着生境由沼泽向盐沼、沙丘、戈壁变化, 土壤类型发生变化, 由沼泽土依次变为漠境盐碱土、风沙土、棕漠土。土壤湿度、容重、盐分、pH等会随之发生变化, 土壤湿度逐渐降低, 容重逐渐升高, 含盐量、pH值先升高后降低(表 2)。其中, 戈壁含水量比沼泽含水量降低了98.58%(P < 0.05), 沙丘和戈壁的土壤含水量差异不显著;戈壁容重比沼泽地增加了0.8倍(P < 0.05), 沙丘和戈壁的容重差异不显著;盐沼湿地含盐量最高, 草本沼泽含盐量最低, 两者相差15.05倍(P < 0.05);盐沼湿地pH值最高, 戈壁最低, 两者差0.1倍(P < 0.05), 盐沼湿地和沙丘土壤pH值差异不显著。
| 生境 Habitats |
土壤湿度 Soil moisture/% |
土壤容重 Soil bulk |
土壤电导率 Soil electrical conductivity/(ms/cm) |
土壤pH Soil pH |
| 草本沼泽Marsh | 48.7±1.99a | 0.89±0.01a | 0.65±0.14b | 8.14±0.06b |
| 盐沼湿地Salt marsh | 14.91±6.44b | 1.25±0.31c | 10.43±3.64a | 8.65±0.1a |
| 沙丘Dune | 4.23±5.29c | 1.58±0.04b | 1.05±0.91b | 8.47±0.39ab |
| 戈壁Gobi | 0.69±0.33c | 1.6±0.04b | 1.46±0.88b | 7.77±0.35c |
| 同一列数据后不同小写字母表示不同生境间差异显著(P < 0.05) | ||||
根据功能性状聚类分析的结果, 结合野外实地观测, 将研究区的优势植物分为9个功能群(图 3)。功能群FG 1主要由沼生或湿生植物构成, 植株的高度较低, 多具根茎, C4光合途径, 能克隆繁殖, 代表性物种有嵩草、苔草、具刚毛荸荠(Eleocharis valleculosa var. setosa)、海韭菜(Triglochin maritima)等。功能群FG 2主要由耐盐、耐旱植物组成, 植物根系发达、高度较高、能克隆繁殖, 主要物种有胀果甘草(Glycyrrhiza inflata)、沙蒿(Artemisia desertorum)等。功能群FG 3主要由耐盐、耐旱的虫媒植物组成, 植物根系也较发达, 可克隆繁殖。功能群FG 4主要由CS策略植物组成, 植物根系发达, 可克隆繁殖, 种子可风播, 根系侵占能力较强, 对缺水生境适应力较强, 主要物种有芦苇、芨芨草、罗布麻(Apocynum venetum)。功能群FG 5由耐旱植物组成, 植物低矮、叶小、肉质、根系发达, 主要植物有蓼子朴(Inula salsoloides)、珍珠猪毛菜(Salsola passerina)等。功能群FG 6以沙生植物为主, 根系较长、植物低矮, 适宜干旱环境, 轻度耐盐, 物种有蒲公英(Taraxacum mongolicum)、达乌里风毛菊(Saussurea davurica Adams)等。功能群FG 7以耐旱的戈壁灌木为主, 植物低矮、叶片减小或减少, 地下根系发达, 主要物种有红砂(Reaumuria soongarica)、骆驼刺(Alhagi sparsifolia)等。功能群FG 8以耐旱的沙生灌木为主, 植株较高, 根系发达, 叶片较小, 部分植物如柽柳为泌盐植物, 可生活在盐沼生境, 叶片较小, 虫媒, 主要由柽柳、梭梭、白刺等组成。功能群FG 9以一年生植物为主, SR策略, 小叶肉质, 耐盐耐旱, 环境适宜时可迅速生长, 主要由白茎盐生草(Halogeton arachnoideus)、刺沙蓬(Salsola tragus)等组成(表 3)。

|
| 图 3 物种功能群聚类分析结果 Fig. 3 Functional groups of species using hierarchical cluster analysis |
| 功能群(FG) Functional group |
描述 Description |
代表性物种 Characteristic species |
| FG 1 | 沼泽湿地生境, 物种以S策略为主, C 4光合途径为主、株高较低、地面或地下芽、能克隆繁殖、自体受粉、多具根茎型 | 嵩草(Kobresia myosuroides); 具刚毛荸荠(Eleocharis valleculosa var. setosa); 圆穗苔草(Carex orbicularis); 海韭菜(Triglochin maritima) |
| FG 2 | 盐沼湿地、沙丘生境, S策略, 地面芽或地上芽、株高较高、能克隆繁殖、根系发达 | 短芒大麦草(Hordeum brevisubulatum); 苦荬菜(Sonchus oleraceus Linn.); 胀果甘草(Glycyrrhiza inflata); 沙蒿(Artemisia desertorum); 羊草(Leymus chinensis (Trin.) Tzvel.); 驴喜草(Onobrychis viciifolia); 苦豆子(Sophora alopecuroides) |
| FG 3 | 沙丘生境, 地面芽或地下芽、能克隆繁殖、虫媒、具增厚或增长的根 | 蕨麻(Potentilla anserina); 马蔺(Iris lactea); 砂引草(Tournefortia sibirica) |
| FG 4 | 沙丘生境, CS策略, 株高较高、能克隆繁殖、根系较长 | 罗布麻(Apocynum venetum); 芦苇(Phragmites australis); 芨芨草(Achnatherum splendens) |
| FG 5 | 沙丘生境, 地面芽或地下芽、能克隆繁殖、S策略、风播、叶茎肉质、小叶具毛、具增厚或增长的根 | 蓼子朴(Inula salsoloides); 珍珠猪毛菜(Salsola passerina) |
| FG 6 | 沙丘生境, S策略, 地面芽植物、株高较低、虫媒、具增厚或增长的根 | 蒲公英(Taraxacum mongolicum); 鹅绒藤(Cynanchum chinense R. Br.); 海乳草(Glaux maritima); 达乌里风毛菊(Saussurea davurica Adams); 驼蹄瓣(Zygophyllum fabago Linn.) |
| FG 7 | 戈壁生境, S策略, 地上芽、能克隆繁殖、植株低矮、叶片减小或减少、增厚或增长的根 | 黑果枸杞(Lycium ruthenicum); 骆驼刺(Alhagi sparsifolia); 红砂(Reaumuria soongarica) |
| FG 8 | 盐沼湿地、沙丘生境, 地上芽、能克隆繁殖、植株较高、S策略、虫媒、叶片较少或较小、增厚或增长的根 | 柽柳(Tamarix austromongolica); 白刺(Nitraria tangutorum); 梭梭(Haloxylon ammodendron) |
| FG 9 | 沙丘或砂质盐碱地生境, SR策略, 一年生植物、C4光合途径、小叶肉质 | 白茎盐生草(Halogeton arachnoideus); 碱蓬(Suaeda glauca); 雾冰藜(Bassia dasyphylla); 刺沙蓬(Salsola tragus); 西伯利亚滨藜(Atriplex sibirica Linn.) |
对物种-样方矩阵进行DCA分析后, 将功能群变量投射到物种-样方排序图中(图 4), 该排序图从左到右代表了水分逐渐减少梯度, 从下向上代表了盐分增加梯度。结果显示, 功能群FG 1和FG 2、FG 4和FG 5、FG 3和FG 6之间正相关且相关性较大;与之相反, 功能群FG 1、FG 2和功能群FG 3、FG 6, 功能群FG 4、FG 5和功能群FG 7、FG 8之间呈较强的负相关关系。功能群FG 1和FG 2同湿生、沼生植物相关性较大, 功能群FG 7、FG 8同旱生植物相关、功能群FG 4和FG 5同盐生植物的相关性较大, 而功能区FG 3、FG 6同盐生、旱生生境都相关。功能群FG 7同戈壁生境相关。

|
| 图 4 功能群同物种关系 Fig. 4 Relation of functional groups with species FG 1, 功能群FG 1; FG 2, 功能群FG 2; FG 3, 功能群FG 3; FG 4, 功能群FG 4; FG 5, 功能群FG 5; FG 6, 功能群FG 6; FG 7, 功能群FG 7; FG 8, 功能群FG 8; FG 9, 功能群FG 9 |
对物种-样方矩阵进行DCA分析后, 将环境变量投射到物种-样方排序图中(图 5), 排序图从左往右代表了土壤水分逐渐减少或土壤容重逐渐增加梯度。结果表明, 盐沼和沙丘环境变量之间正相关, 其他环境变量之间则呈负相关关系。盐沼、沙丘生境内的物种重叠分布较多, 主要物种有柽柳、苦豆子、芦苇、达乌里风毛菊、芨芨草等。草本沼泽、戈壁生境内的物种分布差异明显, 草本沼泽主要物种有嵩草、圆穗苔草、具刚毛荸荠等, 戈壁主要物种有红砂、白刺、珍珠猪毛菜等。物种排序结果同野外观测结果基本一致。

|
| 图 5 物种同生境关系 Fig. 5 Relation of species with habitats Marsh, 沼泽湿地; gobi, 戈壁; salt-mar, 盐沼湿地; dune, 沙丘 |
根据样方-功能群矩阵DCA分析结果(图 6), 不同生境中植物功能群组成不一样。沼泽湿地生境以功能群FG 1为主, 盐沼湿地生境以功能群FG 2为主, 沙丘生境功能群以FG 3、FG 4、FG 5、FG 6、FG 8、FG 9为主, 戈壁生境以功能群FG 7为主。相邻或环境条件相似的生境中, 有部分功能群重叠分布, 如功能群FG 2在沙丘生境中、功能群FG 9在盐沼湿地中、功能群FG 6、FG 8、FG 9在戈壁生境中亦有少量分布(表 3)。

|
| 图 6 功能群同生境关系 Fig. 6 Relation of functional groups with habitats |
用群落加权平均值法对各生境内的功能性状进行分析, 得到各生境的功能性状(表 4)。结果表明, 沼泽湿地植被功能类型多以克隆、地面芽和根茎型草本植物为主, 多为中生或湿生;盐沼湿地植物以叶片被蜡、叶茎肉质、CS策略为主, 株高较高;沙丘植被多以增厚或增长的根、根茎型、风播、CR策略、叶片具毛、一年生、株高较高性状为主;荒漠戈壁植被以虫媒、叶片较少、增厚的根或茎、地上芽等性状为主。表明克隆性状对沼泽湿地环境条件较为适应, 地面芽、根茎等植物性状对沼泽湿地多湿、高盐环境条件较适应, 叶茎肉质、被蜡等植被性对对盐沼湿地环境条件较为适应, 风播、根茎、增厚的根、一年生、叶片具毛等植物性状对固定沙丘环境条件较为适应, 较少叶片、地上芽、虫媒等植被性状对荒漠戈壁环境条件较为适应。
| 生境 Habitats |
克隆类型 Clonality |
增厚的茎或增长的根 Specialized organs |
光合途径 Photosynthetic pathway |
地面(地下)芽植物 Hemicryptophyte (incl. geophyte) |
根茎型 Rhizomatous |
被蜡 Waxy |
风播 Anemochory |
虫媒 Entomophilous |
叶茎肉质 Succulent |
叶片减少 Reduced leaves |
地上芽植物 Chamaephyte |
株高 Plant height |
CS策略 CS strategy |
CR策略 CR strategy |
具毛 Hairy |
一年生 Annual |
| 沼泽湿地Marsh | 1.6 | 0.53 | 1.61 | 1.64 | 0.69 | 0.30 | 0.5 | 0.03 | 0 | 0.03 | 0 | 3.4 | 0.52 | 0 | 0.01 | 0 |
| 盐沼湿地Salt marsh | 0.91 | 0.93 | 0.16 | 0.89 | 0.45 | 0.45 | 0.7 | 0.06 | 0.41 | 0.02 | 0.01 | 3.2 | 0.59 | 0.21 | 0.30 | 0.04 |
| 沙丘Dune | 0.85 | 1.01 | 0.57 | 0.73 | 0.51 | 0.16 | 0.89 | 0.39 | 0.29 | 0.37 | 0.11 | 4.1 | 0.56 | 1.04 | 0.36 | 0.14 |
| 荒漠戈壁Gobi | 0.84 | 0.89 | 0.14 | 0.17 | 0.26 | 0 | 0.06 | 0.41 | 0.14 | 0.95 | 0.66 | 2.2 | 0.01 | 0.3 | 0.02 | 0.1 |
功能性状和环境的排序分析结果与此基本一致(图 7), 克隆、根茎、地面芽、C4植物光合途径、增厚的根或增长的茎、耐受型等功能性状靠近沼泽湿地环境分布, 被蜡、根茎、地面芽等功能性状靠近盐沼湿地环境分布, 风播、耐受、一年生、较高株高等功能性状靠近固定沙丘环境分布, 虫媒、叶茎肉质、叶片减少、地上芽植物、耐受策略等功能性状靠近于戈壁荒漠环境分布。

|
| 图 7 植物功能性状同生境关系 Fig. 7 Relation of functional traits with habitats Anemophi, 风媒; anemocho, 风播; waxy, 叶片被蜡; hemmicry, 地面芽; CS, 竞争耐受策略型; rhizomat, 根茎型; hairy, 叶片具毛; H3, 高度61—100 cm; thorny, 多刺; H4, 高度=>1 m; speciali, 具贮藏器官; entomoph, 虫媒; H2, 高度31—60 cm; phanerop, 高位芽; clonal, 克隆; chamaeph, 地上芽; zoochory, 动物传播; reduced, 较小或较少的叶; leaf/stem, 茎叶肉质; SR, 耐受杂草策略型; annual, 一年生; selp-pol, 自我授粉; no obvio, 无明显传播媒介; photosyn, C4光合途径; H1, 高度1—30 cm |
异质性在自然系统中具有普遍性[31], 植物生长和繁殖所必需的资源(如水分、养分和光照等)以及所处的环境条件(如温度、湿度和干扰等)均呈现异质性分布格局[32], 局域尺度上, 土壤性质的多样化及异质性是影响植物群落组成差异及多样性的一个重要环境因子, 土壤含水率等性质的差异导致植物群落类型发生转换[33-34]。本研究结果与此基本一致。随着生境由沼泽向盐沼、沙丘、戈壁变化, 环境条件发生变化, 海拔逐渐升高, 土壤含水量降低, 土壤容重升高, 土壤盐分和pH值也发生显著变化(表 2);植物物种组成(图 5)、功能群组成随之发生变化(图 6)。这可能是由于:(1)不同的物种对环境具有不同的生态适应性, 物种组成及其功能性状随着区域的环境梯度的变化而发生改变[35-36]。随着生境变化, 研究区域的环境条件呈现出明显的湿生、盐生、沙生、旱生等特征, 不同的环境条件影响了植物的适生分布范围, 导致物种组成及分布发生变化, 形成了该区域植被异质性的空间分布(图 5), 这也符合环境过滤影响着物种和功能性状的分布的环境过滤理论[7, 37]。(2)研究区域内气候条件较为一致, 随着生境条件的改变, 土壤类型、含水量、容重等土壤性质发生显著变化(表 2), 成为影响该区域植物群落组成、结构及多样性的一个重要环境因子, 在土壤条件的作用下, 该区域的功能群的组成和分布随之发生变化。这与钱亦兵等人提出的相似气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异的研究结果一致[38]。
3.2 功能性状对生境的响应植物功能性状是植物长期处于一定的生态条件下, 在不断适应环境的过程中, 通过进化而产生的结果[39], 在一定程度上反映了植物应对环境变化的策略[40]。随着环境条件的变化, 植物对环境做出一定的适应策略, 植物功能性状会发生可塑性变化, 功能性状通过各种性状的调整组合, 形成适应环境的功能性状组[41]。本研究中, 随着生境的变化, 生境中植物的主要功能性状特征随之发生变化, 沼泽湿地生境植被功能性状以克隆、根茎型等功能特征为主, 盐沼湿地以叶片被蜡、叶茎肉质等功能特征为主, 沙丘生境以增厚或增长的根、叶片具毛等功能特征为主, 荒漠戈壁以叶片少、增厚或增长的根等功能特征为主(表 4), 说明生境条件的改变, 导致功能群的功能性状发生适应性的改变。这主要是因为:(1)沼泽湿地生境中, 地势低洼, 土壤含水量最高(表 2), 形成湿生环境条件。该环境中水分充足, 植株的含水量、水分的疏导效率较高、木质化程度较低。为适应该环境的低光、缺氧、养分匮乏不良影响, 获取更高的CO2固定效率, 提高繁殖能力和存活率, 植物光合途径以C4循环为主, 繁殖方式也适应性的以克隆生长和无性繁殖为主[42]。(2)盐沼湿地生境中, 土壤含水量较高、盐渍化程度较大(表 2), 易对植物形成盐胁迫。盐胁迫会造成植物的生理性干旱, 影响着植物的生长发育。盐生植物则通过改变外部形态, 调节生理机制, 应对不良生境条件的影响。通过减少叶片、具表皮毛或表皮被蜡等策略, 最大限度的减少水分蒸腾面积, 通过茎、叶中发达的储水组织以保持水分, 稀释细胞内的盐浓度, 使细胞不致因盐度过高而受到伤害, 表现出了适应旱生和盐生生境的特征。(3)沙丘生境中, 土壤含水量较低, 土质松软、沙化, 沙埋干扰严重, 形成干旱少雨环境。为适应该环境, 沙丘植物通过风播占据有利生境进行繁殖;增强的根系获取地下水资源;叶片具毛、被蜡以减少蒸腾作用;在一个生育期内完成生命周期, 并以种子状态度过干旱少雨不良环境等策略, 最大限度的利用周边环境资源, 完成生活史。克隆植物可通过形态结构的可塑性调节(形态可塑性)、克隆分株选择性放置(觅食行为)、分株间物质传输与共享(克隆整合或生理整合)、相连分株形态和生理等功能的特化(克隆分工)等策略[43-45], 在异质性生境中占据有利斑块或逃离不利生境, 对沙丘生境较为适应, 因此该生境中克隆植物分布较多, 例如芦苇。(4)戈壁生境中, 土壤含水量最低, 土壤容重最大, 土壤矿质营养贫乏。此环境条件下的植物以矮小的灌木为主, 具有明显的旱生植物特点。为适应该旱生环境, 植物多采用纵深发达的地下根系、较低的高度、较小的叶片等性状策略, 来增强对地下水分的吸收、减少水分消耗、降低水分蒸腾, 维持株体内部较高的含水量和水势, 保障植物的正常的生长发育。
4 结论基于对功能性状进行系统聚类分析的功能群分类法, 考虑了物种的综合性状, 可以区分不同生境下的功能群差异, 比提前定义功能群的分类方法更客观、有效, 可适用于多种尺度的功能群研究。局域尺度上, 生境的异质性, 尤其是土壤条件的异质性是影响植物群落组成及主要功能性状的重要环境因子。生境条件不同, 功能群组成不同。随着生境条件的改变, 植物功能性状会发生适应性的变化, 沼泽湿地生境内植被的功能性状以克隆、根茎型等特征为主, 盐沼湿地以叶片被蜡、叶茎肉质等特征为主, 沙丘生境以增厚或增长的根、叶片具毛等特征为主, 荒漠戈壁以叶片少、增厚或增长的根等特征为主。本研究分析了植物功能群对不同生境的适应策略, 可为当地生态管理、生态恢复以及物种更替的预测等提供一定的理论依据。文中仅对不同生境下的功能群组成及其性状特征进行了分析, 而未对同一生境条件下功能群异质性分布的机制进行深入研究, 还需进一步实验验证。
| [1] |
Díaz S, Cabido M. Plant functional types and ecosystem function in relation to global change. Journal of Vegetation Science, 1997, 8(4): 463-474. DOI:10.2307/3237198 |
| [2] |
唐海萍, 蒋高明. 植物功能型及其生态学意义. 应用生态学报, 2000, 11(3): 461-464. DOI:10.3321/j.issn:1001-9332.2000.03.033 |
| [3] |
Díaz S, Cabido M, Zak M, Carretero E M, Araníbar J. Plant functional traits, ecosystem structure and land-use history along a climatic gradient in central-western Argentina. Journal of Vegetation Science, 1999, 10(5): 651-660. DOI:10.2307/3237080 |
| [4] |
Petchey O L. On the statistical significance of functional diversity effects. Functional Ecology, 2004, 18(3): 297-303. DOI:10.1111/j.0269-8463.2004.00852.x |
| [5] |
Diaz S, Cabido M. Vive la différence:plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655. |
| [6] |
Wright S J. Tropical forests in a changing environment. Trends in Ecology & Evolution, 2005, 20(10): 553-560. |
| [7] |
Cornwell W K, Schwilk D W, Ackerly D D. A trait-based test for habitat filtering:convex hull volume. Ecology, 2006, 87(6): 1465-1471. DOI:10.1890/0012-9658(2006)87[1465:ATTFHF]2.0.CO;2 |
| [8] |
Lebrija-Trejos E, Pérez-García E A, Meave J A, Bongers F, Poorter L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology, 2010, 91(2): 386-398. DOI:10.1890/08-1449.1 |
| [9] |
Maire V, Gross N, Börger L, Proulx R, Wirth C, Da Silveira Pontes L, Soussana J F, Louault F. Habitat filtering and niche differentiation jointly explain species relative abundance within grassland communities along fertility and disturbance gradients. New Phytologist, 2012, 196(2): 497-509. DOI:10.1111/j.1469-8137.2012.04287.x |
| [10] |
陈玉福, 于飞海, 张称意, 董鸣. 根茎禾草沙鞭的克隆生长在毛乌素沙地斑块动态中的作用. 生态学报, 2001, 21(11): 1745-1750. DOI:10.3321/j.issn:1000-0933.2001.11.001 |
| [11] |
Verheyen K, Honnay O, Motzkin G, Hermy M, Foster D R. Response of forest plant species to land-use change:a life-history trait-based approach. Journal of Ecology, 2003, 91(4): 563-577. DOI:10.1046/j.1365-2745.2003.00789.x |
| [12] |
Quétier F, Lavorel S, Thuiller W, Davies I. Plant-trait-based modeling assessment of ecosystem-service sensitivity to land-use change. Ecological Applications, 2007, 17(8): 2377-2386. DOI:10.1890/06-0750.1 |
| [13] |
Klimesova J, Latzel V, De Bello F, Van Groenendael J M. Plant functional traits in studies of vegetation changes in response to grazing and mowing:towards a use of more specific traits. Preslia, 2008, 80(3): 245-253. |
| [14] |
Díaz S, Lavorel S, McIntyre S, Falczuk V, Casanoves F, Milchunas D G, Skarpe C, Rusch G, Sternberg M, Noy-Meir I, Landsberg J, Zhang W, Clark H, Campbell B D. Plant trait responses to grazing-a global synthesis. Glob Change Biology, 2007, 13(2): 313-341. DOI:10.1111/j.1365-2486.2006.01288.x |
| [15] |
Lloret F, Vilá M. Diversity patterns of plant functional types in relation to fire regime and previous land use in mediterranean woodlands. Journal of Vegetation Science, 2003, 14(3): 387-398. DOI:10.1111/j.1654-1103.2003.tb02164.x |
| [16] |
Keeley J E, Pausas J G, Rundel P W, Bond W J, Bradstock R A. Fire as an evolutionary pressure shaping plant traits. Trends in Plant Science, 2011, 16(8): 406-411. DOI:10.1016/j.tplants.2011.04.002 |
| [17] |
Pillar V D, Blanco C C, Müller S C, Sosinski E E, Joner F, Duarte L D S. Functional redundancy and stability in plant communities. Journal of Vegetation Science, 2013, 24(5): 963-974. DOI:10.1111/jvs.12047 |
| [18] |
Box E O. Plant functional types and climate at the global scale. Journal of Vegetation Science, 1996, 7(3): 309-320. DOI:10.2307/3236274 |
| [19] |
De Bello F, Lavorel S, Díaz S, Harrington R, Cornelissen J H C, Bardgett R D, Berg M P, Cipriotti P, Feld C K, Hering D, Da Silva P M, Potts S G, Sandin L, Sousa J P, Storkey J, Wardle D A, Harrison P A. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodiversity and Conservation, 2010, 19(10): 2873-2893. DOI:10.1007/s10531-010-9850-9 |
| [20] |
Kraft N J B, Ackerly D D. Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest. Ecological Monographs, 2010, 80(3): 401-422. DOI:10.1890/09-1672.1 |
| [21] |
Pillar V D, Duarte L D S. A framework for metacommunity analysis of phylogenetic structure. Ecology Letters, 2010, 13(5): 587-596. DOI:10.1111/j.1461-0248.2010.01456.x |
| [22] |
张志东, 臧润国, 丁易. 海南岛霸王岭热带天然林景观中木本植物功能群划分及其潜在分布. 林业科学, 2009, 45(10): 1-8. |
| [23] |
García-Mora M R, Gallego-Fernández J B, García-Novo F. Plant functional types in coastal foredunes in relation to environmental stress and disturbance. Journal of Vegetation Science, 1999, 10(1): 27-34. |
| [24] |
Gallego-Fernández J B, Martínez M L. Environmental filtering and plant functional types on Mexican foredunes along the gulf of mexico. Écoscience, 2011, 18(1): 52-62. DOI:10.2980/18-1-3376 |
| [25] |
吕子君, 卢欣石. 甘肃河西走廊草地沙质荒漠化监测与治理对策. 江西农业大学学报, 2005, 27(3): 365-370. DOI:10.3969/j.issn.1000-2286.2005.03.011 |
| [26] |
马仲武, 王新源, 王小军, 丁乾平, 曲浩. 甘肃省酒泉市土地沙漠化现状及动态分析. 中国农业资源与区划, 2018, 39(3): 141-147. |
| [27] |
Yang H T, Li X R, Liu L C, Gao Y H, Li G, Jia R L. Soil water repellency and influencing factors of nitraria tangutorun nebkhas at different succession stages. Journal of Arid Land, 2014, 6(3): 300-310. DOI:10.1007/s40333-013-0199-2 |
| [28] |
张雅琼, 梁存柱, 王炜, 王立新, 彭江涛, 闫建成, 贾成朕. 芨芨草群落土壤盐分特征. 生态学杂志, 2010, 29(12): 2438-2443. |
| [29] |
Mahdavi P, Bergmeier E. Plant functional traits and diversity in sand dune ecosystems across different biogeographic regions. Acta Oecologica, 2016, 74: 37-45. DOI:10.1016/j.actao.2016.06.003 |
| [30] |
余晶晶.千岛湖常见木本植物功能性状、功能群组成及其与岛屿空间特征的关系[D].杭州: 浙江大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10335-1016280053.htm
|
| [31] |
Kotliar N B, Wiens J A. Multiple scales of patchiness and patch structure:a hierarchical framework for the study of heterogeneity. Oikos, 1990, 59(2): 253-260. DOI:10.2307/3545542 |
| [32] |
Jackson R B, Caldwell M M. The scale of nutrient heterogeneity around individual plants and its quantification with geostatistics. Ecology, 1993, 74(2): 612-614. DOI:10.2307/1939320 |
| [33] |
Gregory S V, Swanson F J, McKee W A, Cummins K W. An ecosystem perspective of riparian zones. BioScience, 1991, 41(8): 540-551. DOI:10.2307/1311607 |
| [34] |
Naiman R J, Décamps H. The ecology of interfaces:riparian zones. Annual Review of Ecology and Systematics, 1997, 28: 621-658. DOI:10.1146/annurev.ecolsys.28.1.621 |
| [35] |
卜文圣, 臧润国, 丁易, 张俊艳, 阮云泽. 海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化. 生物多样性, 2013, 21(3): 278-287. |
| [36] |
Diaz S, Cabido M, Casanoves F. Plant functional traits and environmental filters at a regional scale. Journal of Vegetation Science, 1998, 9(1): 113-122. DOI:10.2307/3237229 |
| [37] |
Cornwell W K, Ackerly D D. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal california. Ecological Monographs, 2009, 79(1): 109-126. DOI:10.1890/07-1134.1 |
| [38] |
钱亦兵, 蒋进, 吴兆宁. 艾比湖地区土壤异质性及其对植物群落生态分布的影响. 干旱区地理, 2003, 26(3): 217-222. DOI:10.3321/j.issn:1000-6060.2003.03.004 |
| [39] |
Carmona D, Lajeunesse M J, Johnson M T J. Plant traits that predict resistance to herbivores. Functional Ecology, 2011, 25(2): 358-367. DOI:10.1111/j.1365-2435.2010.01794.x |
| [40] |
孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150-165. DOI:10.3321/j.issn:1005-264X.2007.01.019 |
| [41] |
Soliveres S, Maestre F T, Bowker M A, Torices R, Quero J L, García-Gómez M, Cabrera O, Cea A P, Coaguila D, Eldridge D J, Espinosa C I, Hemmings F, Monerris J J, Tighe M, Delgado-Baquerizo M, Escolar C, García-Palacios P, Gozalo B, Ochoa V, Blones J, Derak M, Ghiloufi W, Gutiérrez J R, Hernández R M, Noumi Z. Functional traits determine plant co-occurrence more than environment or evolutionary relatedness in global drylands. Perspectives in Plant Ecology, Evolution and Systematics, 2014, 16(4): 164-173. DOI:10.1016/j.ppees.2014.05.001 |
| [42] |
Grace J B. The adaptive significance of clonal reproduction in angiosperms:an aquatic perspective. Aquatic Botany, 1993, 44(2/3): 159-180. |
| [43] |
Ikegami M, Van Hal S, Van Rheenen J W A, Whigham D F, Werger M J A. Spatial division of labour of schoenoplectus americanus. Plant Ecology, 2008, 199(1): 55-64. |
| [44] |
Dong M, Yu F H, Alpert P. Ecological consequences of plant Clonality. Annals of Botany, 2014, 114(2): 367-367. DOI:10.1093/aob/mcu137 |
| [45] |
Wang Z W, Li Y H, During H J, Li L H. Do clonal plants show greater division of Labour morphologically and physiologically at higher patch contrasts?. PLoS One, 2011, 6(9): e25401. DOI:10.1371/journal.pone.0025401 |