 2020, Vol. 40
2020, Vol. 40文章信息
- 杨颖, 刘鹏霞, 周红宏, 夏利花
- YANG Ying, LIU Pengxia, ZHOU Honghong, XIA Lihua
- 近15年长江口海域海洋生物变化趋势及健康状况评价
- Evaluation of the biodiversity variation and ecosystem health assessment in Changjiang estuary during the past 15 years
- 生态学报. 2020, 40(24): 8892-8904
- Acta Ecologica Sinica. 2020, 40(24): 8892-8904
- http://dx.doi.org/10.5846/stxb201912272811
-
文章历史
- 收稿日期: 2019-12-27






2. 自然资源部海洋生态监测与修复技术重点实验室, 上海 201206
2. Key Laboratory of Marine Ecological Monitoring and Restoration Technologies, MNR, Shanghai 201206, China
长江口是我国最大河口, 受河流系统、气候变化以及人类活动的深刻影响, 其生态系统具有典型河口区域性特征。由于长江向河口区输入丰富的营养物质, 长江口海域历史上是著名渔场。但随着开发活动加剧, 长江口海域面临严峻的生态环境问题, 多年来水体严重富营养化, 导致赤潮频发[1-2], 低氧现象突出[3];海域生境破碎化[4], 海洋生物种类和数量都处于较低水平, 鱼类资源减少[5], 近30a长江口生态系统健康状况逐渐下降[6], 生态系统总体处于亚健康状态[7-8]。长江口由于其河口流系复杂性、生态系统典型性, 吸引了众多学者的关注, 本世纪以来, 开展了大量关于长江口海域生态评价研究工作[9-13]。王金辉等开展了长江口及邻近水域的生物多样性变化趋势分析[9], 王江涛等开展了长江口海域近50a来营养盐的变化及其对浮游植物群落演替的影响研究[10], 黄海燕等开展了2004—2016年夏季长江口浮游植物及其影响研究[11], 沈新强等基于长江口生态恶化的现状, 提出建立以长江口生态系统为基础的管理机制[13]。以上成果均基于部分年份或部分监测指标开展研究, 本文系统梳理了2004年长江口生态监控区建立以来的监测资料, 基于15a连续监测数据, 对长江口海域海洋生物变化趋势及健康状况进行评价, 以期为长江经济带保护提供基础支撑。
1 材料与方法 1.1 资料来源 1.1.1 评价区域与站位评价区域为长江口及邻近海域(图 1), 范围为徐六泾以东至123°, 启东嘴以南至杭州湾上海海域以北的海域, 面积为13668km2。

|
| 图 1 长江口海域监测站位示意图 Fig. 1 The monitoring sites in Changjiang River estuary |
数据引用了国家海洋局东海环境监测中心2004—2018年每年8月的长江口生态监控区监测水质数据(DIN、DIP、重金属、石油类等)和海洋生物(浮游植物、浮游动物和底栖生物)数据。文中除标明的文献数据外, 其他历史数据均来源于国家海洋局东海环境监测中心历年在长江口海域的业务化监测, 每年的站位数量和位置与图 1或有不同。
采样及监测方法按照《海洋监测规范》 [14]中标准方法。浮游植物采用浅水Ⅲ型浮游生物网垂直拖网。浮游动物采用浅水Ⅰ型和Ⅱ型浮游生物网, 种类组成(包括优势种和常见种等)结合浅水Ⅰ型和Ⅱ型浮游生物网样品分析;生物密度、生物量采用浅水Ⅰ型浮游生物网采集的样品数据。底栖生物定量样品用采泥器采集底泥, 每站采样面积不少于0.2m2, 选取20%站位采集阿氏拖网样品。
1.2 评价方法与标准 1.2.1 生物多样性评价采用种类多样性指数H′ (Shannon-Weaver index)、种类丰富度指数d (Margalef′S index)、均匀度指数J′ (Pielou index)和优势度指数Y 对海洋生物多样性进行评价[14], 计算公式如式(1)—(4)。式中:S—种类数;N—所有种的个体总数;Pi—样品中第i种个体数占总个体数的比例;fi—第i种在各样品中的出现频率;ni—样品中第i种生物个体数。取Yi≥0.02的种类为优势种类[15]。
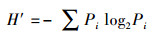
|
(1) |
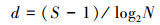
|
(2) |
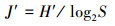
|
(3) |
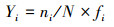
|
(4) |
引用《近岸海洋生态健康评价指南》 [16]中河口生态系统健康状况评价方法, 对长江口海洋生物健康状况进行评价, 评价指标及分级标准如表 1, 其中Ⅰ级赋值50, Ⅱ级赋值30, Ⅲ级赋值10, 生物健康指数位于[35, 50]为健康, [20, 35)为亚健康, [10, 20)为不健康。
| 指标Parameters | Ⅰ | Ⅱ | Ⅲ |
| 浮游植物密度 Phytoplankton density(胞/m3) |
>50%A—≤150%A | >10%A—≤50%A或 >150%A—≤200%A |
≤10%A或>200%A |
| 浮游动物密度 Zooplankton density(个/m3) |
>75%B—≤125%B | >50%B—≤75%B或 >125%B—≤150%B |
≤50%B或>150%B |
| 浮游动物生物量 Zooplankton biomass(mg/m3) |
>75%C—≤125%C | >50%C—≤75%C或 >125%C—≤150%C |
≤50%C或>150%C |
| 底栖动物密度 Benthos density(个/m2) |
>75%D—≤125%D | >50%D—≤75%D或 >125%D—≤150%D |
≤50%D或>150%D |
| 底栖动物生物量 Benthos biomass(g/m2) |
>75%E—≤125%E | >50%E—≤75%E或 >125%E—≤150%E |
≤50%E或>150%E |
| A、B、C、D和E表示长江口8月份浮游生物和大型底栖生物评价依据。其中A为30×105个/m3;B为5×103个/m3;C为480.0mg/m3;D为20个/m2;E为10.0g/m2 | |||
对近15年长江口海域浮游植物、浮游动物、底栖生物的种类组成、密度变化、生物多样性情况进行分析评价。
2.1 浮游植物 2.1.1 种类组成(1) 种类数变化
近15年长江口海域夏季浮游植物共鉴定出8门373种(定种367种), 其中硅藻(Diatomophyceae) 237种(定种232种), 占65.3%;甲藻(Dinophyceae) 94种, 占25.2%;绿藻(Chlorophyceae)25种, 占6.7%;蓝藻(Cyanophyceae)11种, 占2.9%;金藻(Chrysophyceae)3种, 裸藻(Euglenophyta)2种(定种1种), 动鞭藻(Zooflagellates)1种, 隐藻(Cryptophyta)1种。王金辉等[9]报道1997年10月—2002年5月长江口及邻近水域浮游植物138属402种, 与之相比, 近15年鉴定的浮游植物总种类数有所减少, 其中硅藻数量减少22种, 绿藻、蓝藻分别减少11种, 其余门类也有所减少, 但甲藻总数量增加41种。
各年份鉴定出浮游植物4—6门类、60—149种, 平均107种, 2009年最低, 2013年最高。平均种类数与20世纪80年代相比略低, 与90年代相比持平[9]。各年份种类数分布趋势如图 2所示, 浮游植物种类数变化趋势以2009年为分界点, 之前种类数震荡降低, 之后呈现震荡升高趋势。

|
| 图 2 2004—2018年长江口海域浮游植物种类变化 Fig. 2 Variation of net-phytoplankton species in the Changjiang River Estuary from 2004 to 2018 |
(2) 种类组成变化
各年份种类组成上硅藻均占绝对优势, 种类数占浮游植物总种类数范围57.6%—76.7%, 甲藻次之, 占13.3%—33.3%。年际分布上, 硅藻与甲藻占比高低基本互补。各年份硅藻、甲藻比例变化情况如图 3所示, 从20世纪80年代以来, 长江口海域硅藻种类占比呈现震荡下降的趋势, 甲藻种类占比有所升高[9]。按照年代划分, 20世纪80年代硅藻占比在80%以上;1990年—2009年的20年间, 硅藻的平均占比约在70%;2010年—2108年硅藻平均占比63%, 且相对稳定, 变化幅度不大。甲藻占比从1990年—2007年震荡升高, 从5%大幅升至33.3%;2008—2018年总体变化不大, 平均占比23.3%。硅藻与甲藻比例的变化, 与硅藻赤潮与甲藻赤潮的演变趋势相一致, 在针对长江口邻近海域的研究中发现, 有害藻华优势类群近期已由硅藻逐渐转变为甲藻[1]。据统计, 20世纪80—90年代骨条藻(Skeletonema spp.)在赤潮优势种中所占比例超过30%, 到2000年其比例下降至24%;而原甲藻(Prorocentrum)在20世纪80—90年代所占比例仅有10%左右, 到2000年前后增加到36%[1]。关注长江口海域硅藻、甲藻演替的研究较多[1, 9, 10], 显示20世纪80年代以来本海域浮游植物群落结构发生巨大变化。从本文的趋势变化分析, 2010年以来, 硅藻、甲藻群落结构进入了一个新的平衡状态。

|
| 图 3 长江口海域浮游植物中硅藻、甲藻种类占比变化 Fig. 3 Variation of Diatomophyceae and Dinophyceaein ratio in the Changjiang River Estuary |
近15年长江口海域浮游植物的年度平均细胞密度范围为(8.37—7480)×105个/m3, 平均值为1.07×108个/m3, 2014年最低, 2012年最高。长江口海域为赤潮高发区域[1], 每年5—8月为赤潮高发时段, 2011、2012年监测期间, 部分站位正在发生中肋骨条藻赤潮, 异于正常时期水体状态, 扣除该2年份数据, 其余年份浮游植物密度均值为3.82×107个/m3。与1997—2002年长江口海域浮游植物丰水期平均密度(2.96×107个/m3)[9]相比有所上升。总体分布上, 长江口海域夏季浮游植物平均细胞丰度呈上升趋势(图 4), 与黄海燕[11]等研究结果一致。

|
| 图 4 2004—2018年长江口夏季浮游植物丰度变化 Fig. 4 Variation of net-phytoplankton density in summer in the Changjiang River Estuary from 2004 to 2018 |
近15年长江口海域浮游植物的种类多样性变化情况如图 5所示。生物多样性指数范围为0.81—2.18, 2011年最低, 2015年最高, 2004—2008年浮游植物多样性指数波动较小, 2009—2018年震荡幅度变大, 但总体上无明显变化规律。丰富度指数范围0.45—1.28, 2004、2014年最低, 2018年最高, 各年份有一定波动, 2014—2018年逐年升高。均匀度指数范围0.21 —1.63, 2016年最低, 2014年最高, 除了2014、2015年两个年份异偏高, 其他年份物种均匀度指数较为平稳且缓慢降低。

|
| 图 5 2004—2018年长江口海域浮游植物多样性变化趋势 Fig. 5 Variation of net-phytoplankton biodiversity in the Changjiang River Estuary from 2004 to 2018 |
近15年夏季长江口海域浮游植物优势种如表 2所示。以下藻种在不同年份成为优势藻种:中肋骨条藻(Skeletonemacostatum)、佛氏海毛藻(Thalassiothrixfrauenfeldii)、假弯角毛藻(Chaetocerospseudocurviser)、尖刺菱形藻(Nitzschiapungens)、洛氏角毛藻(Chaetoceroslorenzianus)、菱形海线藻(Thalassionemanitzschioides)、尖刺拟菱形藻(Pseudo-nitzschiapungens)、旋链角毛藻(Chaetoceroscurvisetus)、梭角藻(Ceratiumfusus)、虹彩圆筛藻(Coscinodiscus oculus-iridis)、笔尖形根管藻(Rhizosoleniastyliformis)等。2004—2013年, 中肋骨条藻连续10a为本海域第一优势种;2014年梭角藻变为第一优势种;2015年尖刺拟菱形藻为第一优势种, 中肋骨条藻不是优势藻种;2016、2018年中肋骨条藻又成为第一优势种, 且优势度达到历史最高。
| 年份Year | 优势种Dominant species |
| 2004 | 中肋骨条藻、佛氏海毛藻、假弯角毛藻 |
| 2005 | 中肋骨条藻、尖刺菱形藻、洛氏角毛藻、 |
| 2006 | 中肋骨条藻、菱形海线藻、尖刺拟菱形藻 |
| 2007 | 中肋骨条藻 |
| 2008 | 中肋骨条藻 |
| 2009 | 中肋骨条藻 |
| 2010 | 中肋骨条藻、旋链角毛藻 |
| 2011 | 中肋骨条藻 |
| 2012 | 中肋骨条藻、尖刺拟菱形藻 |
| 2013 | 中肋骨条藻 |
| 2014 | 梭角藻、中肋骨条藻、尖刺拟菱形藻、虹彩圆筛藻、笔尖形根管藻 |
| 2015 | 尖刺拟菱形藻、洛氏角毛藻、旋链角毛藻 |
| 2016 | 中肋骨条藻、尖刺拟菱形藻 |
| 2017 | 尖刺拟菱形藻、中肋骨条藻、洛氏角毛藻 |
| 2018 | 中肋骨条藻 |
各年份第一优势种平均优势度变化情况如图 6所示, 历年第一优势种平均优势度变化范围0.28—0.88, 其中2014年最低, 2018年最高。长江口海域优势种群浮游植物的爆发性生长引起优势度的剧烈变化, 使群落结构处于不稳定状态。

|
| 图 6 2004—2018年长江口浮游植物第一优势种优势度变化 Fig. 6 Variation of net-phytoplankton first dominant species′ dominance index in the Changjiang River Estuary from 2004 to 2018 |
近15年长江口海域夏季共鉴定浮游动物(I型网和II型网)11门472种(不含浮游幼虫), 其中节肢动物(Arthropoda)298种, 是最大类群;腔肠动物(Coelenterata)68种, 环节动物(Annelida)30种, 软体动物(Mollusca)22种, 尾索动物(Tunicata)22种, 毛颚动物(Chaetognatha)20种, 栉水母动物(Ctenophora)5种, 原生动物(Protozoa)3种, 脊索动物(Chordata)2种, 棘皮动物(Echinodermata)1种, 轮虫动物(Rotifera)1种。王金辉[9]等报道1997年10月—2002年5月长江口及邻近水域浮游动物19个类群480种(含浮游幼虫63种), 与之相比, 近15年夏季鉴定的浮游动物种类数有所增加。
各年份鉴定出浮游动物75—186种, 平均99种, 2005年最低, 2008年最高。各年份种类数分布趋势如图 7所示, 2007—2009年鉴定的种类数较高, 其他年份有一定波动, 但差异不大。主要类群为节肢动物, 各年份平均占比63.6%, 其中2004—2008年节肢动物占比持续减少, 主要因素是节肢动物中最大类群桡足类种类减少;腔肠动物各年份平均占比14.3%%, 其中2007、2008年有较大波动, 2007年仅鉴定出1种, 2008年鉴定出26种, 其余年份变化不大。2009—2018年, 节肢动物与腔肠动物占比均处于动态平衡状态, 比例互补。

|
| 图 7 2004—2018年长江口海域浮游动物种类变化 Fig. 7 Variation of zooplankton species in the Changjiang River Estuary from 2004 to 2018 |
近15年夏季长江口海域浮游动物I型网的年度平均密度范围为203—2942个/m3, 平均值为829个/m3, 2006年最低, 2017年最高。总体分布上, 夏季浮游动物的平均密度呈上升趋势(图 8), 其中2004—2015年在一定范围内波动, 总体平稳;2016、2017年平均密度异常升高, 2018年大幅回落。

|
| 图 8 2004—2018年长江口海域浮游动物密度和生物量变化情况 Fig. 8 Variation of zooplankton density and biomass in the Changjiang River Estuary from 2004 to 2018 |
浮游动物I型网的年度平均生物量变化范围为406—1729mg/m3, 平均值为750 mg/m3, 2013年最低, 2018年最高。总体分布上, 夏季浮游动物的平均生物量呈上升趋势(图 8)。与1997—2002年长江口海域浮游动物丰水期平均生物量(292.7 mg/m3)[9]相比, 近15年浮游动物生物量年均值有明显上升。2004—2014年浮游动物密度和生物量变化趋势基本一致, 2015—2018年, 变化趋势相反, 可能预示着浮游动物群落结构发生较大变化。
2.2.3 生物多样性长江口海域浮游动物(I型网)种类多样性变化情况如图 9所示。年度平均生物多样性指数范围为2.16—3.24, 2016年最低, 2006年最高, 浮游动物多样性指数总体波动较小, 呈现小幅震荡下降的趋势。平均丰富度指数范围0.68—3.79, 2014年最低, 2008年最高, 各年份波动幅度较大, 2008—2014年持续6年大幅下降, 2015年后有所回升。均匀度指数范围0.60 —2.38, 除了2014、2015年两个年份异常偏高, 其他年份均匀度指数平稳。

|
| 图 9 2004—2018年长江口海域浮游动物多样性变化情况 Fig. 9 Variation of zooplankton biodiversity in the Changjiang River Estuary from 2004 to 2018 |
近15年长江口海域浮游动物(I型网)优势种如表 3所示。各年份出现的优势种共有17种, 其中背针胸刺水蚤(Centropagesdorsispinatus)在13个年份中作为优势种出现, 其中2008、2012、2014—2018年共7年为第一优势种;太平洋纺锤水蚤(Acartiapacifica)在11个年份中作为优势种出现, 2007年为第一优势种;真刺唇角水蚤(Labidoceraeuchaeta)在7个年份中作为优势种出现, 2011年为第一优势种;肥胖箭虫(Sagitta enflata)在8个年份中作为优势种出现;双生水母(Diphyeschamissonis)在7个年份中作为优势种出现;其他优势种类均为个别年份出现。
| 年份Year | 优势种Dominant species |
| 2004 | 五角水母/Muggiaeaatlantica、针刺拟哲水蚤/Paracalanusaculeatus、真刺唇角水蚤、双生水母、背针胸刺水蚤、太平洋纺锤水蚤 |
| 2005 | 真刺唇角水蚤、针刺拟哲水蚤、小拟哲水蚤/Paracalanusparvus |
| 2006 | 真刺唇角水蚤、背针胸刺水蚤、百陶箭虫/ Sagitta bedoti、肥胖箭虫、海龙箭虫/Sagitta nagae |
| 2007 | 太平洋纺锤水蚤、中华哲水蚤/Calanussinicus、背针胸刺水蚤、肥胖箭虫 |
| 2008 | 背针胸刺水蚤、太平洋纺锤水蚤、双生水母、肥胖箭虫、肥胖三角溞/Evadne tergestina |
| 2009 | 双生水母、太平洋纺锤水蚤、肥胖箭虫、中华哲水蚤、真刺拟哲水蚤、肥胖三角溞、真刺唇角水蚤 |
| 2010 | 太平洋纺缍水蚤、背针胸刺水蚤、肥胖箭虫、真刺唇角水蚤、双生水母、中型莹虾/ Lucifer intermedius |
| 2011 | 真刺唇角水蚤、太平洋纺锤水蚤、火腿许水蚤/Schmackeriapoplesia、背针胸刺水蚤 |
| 2012 | 背针胸刺水蚤、太平洋纺锤水蚤、虫肢歪水蚤/Tortanusvermiculus、双生水母 |
| 2013 | 太平洋纺锤水蚤、背针胸刺水蚤、火腿许水蚤、真刺唇角水蚤、虫肢歪水蚤 |
| 2014 | 背针胸刺水蚤、真刺唇角水蚤、糠虾幼体、火腿许水蚤、太平洋纺锤水蚤 |
| 2015 | 背针胸刺水蚤、双生水母、肥胖箭虫、肥胖三角溞、针刺拟哲水蚤 |
| 2016 | 背针胸刺水蚤、太平洋纺锤水蚤、真刺唇角水蚤 |
| 2017 | 背针胸刺水蚤、肥胖箭虫 |
| 2018 | 背针胸刺水蚤、肥胖箭虫、中华哲水蚤、精致真刺水蚤/Enchaetaconcinna /、亚强次真哲水蚤/Eucalanussubcrassus、太平洋纺锤水蚤、双生水母 |
各年份第一优势种优势度范围为0.06—0.33, 2004—2018年平均值为0.18(图 10)。2004—2014年的10年中第一优势种优势度震荡上升, 近5年第一优势种的优势度波动较大。

|
| 图 10 2004—2018年浮游动物第一优势种优势度变化 Fig. 10 Variation of zooplankton first dominant species′ dominance index in the Changjiang River Estuary from 2004 to 2018 |
近15年长江口海域夏季共鉴定底栖生物9门435种, 其中环节动物(Annelida)182种, 是底栖生物中的最大类群。软体动物(Mollusca) 90种, 节肢动物(Arthropoda)89种, 脊索动物(Chordata)41种, 棘皮动物(Echinodermata)23种, 腔肠动物(Coelenterata) 4种, 星虫动物(Sipuncula) 3种, 螠虫动物(Echiura) 2种, 纽形动物(Nemertea)1种。与1997年10月—2002年5月长江口及邻近水域底栖生物种类数(4个类群97种)[9]相比, 近15年鉴定的底栖生物种类数大幅增加。
各年份鉴定出底栖生物43—124种, 平均81种, 其中2004年最低, 2017年最高(图 11), 近15年底栖生物种类数有明显升高的趋势, 2015—2018年均超过110种。环节动物为主要类群, 各年份平均占比51.6%, 2004—2009年环节动物占比逐年升高, 2009—2018年则逐年下降。节肢动物各年份平均占比17.7%, 2007—2014年占比较其他年份略低。总体上环节动物与节肢动物占比比例互补。

|
| 图 11 2004—2018年长江口海域底栖生物种类变化 Fig. 11 Variation of benthos species in the Changjiang River Estuary from 2004 to 2018 |
近15年夏季长江口海域底栖生物的平均密度范围为13.5—174个/m2, 各年份总平均值为98个/m2, 2014年最低, 2018年最高。各年份平均栖息密度变化剧烈, 2010—2014年大幅下降, 2014—2018年反弹, 由最低位上升到最高位(图 12)。年平均生物量变化范围为0.26—13.6g/m2, 均值5.52 g/m2, 2014年最低, 2018年最高。与1997—2002年长江口海域底栖生物平均生物量(8.93 g/m2)[9]相比, 近15年生物量有所下降。各年份中生物量的变化趋势总体与生物密度一致, 震荡幅度大, 2005—2009年呈明显下降趋势, 2010年异常升高后即迅速下降, 2014年触底, 2018年反弹至最高位。

|
| 图 12 2004—2018年长江口海域底栖生物密度和生物量变化 Fig. 12 Variation of benthos density and biomass in the Changjiang River Estuary from 2004 to 2018 |
近15年长江口海域底栖生物多样性变化情况如图 13所示。生物多样性指数范围为0.99—2.45, 其中2012年最低, 2014年最高。2004—2013年底栖生物多样性指数总体呈下降趋势, 2014年反弹至最高点, 近5年有所波动。丰富度指数范围0.52—1.46, 2004年最低, 2016年最高, 各年份有一定波动。均匀度指数范围0.47 —2.45, 2011年最低, 2014年最高, 总体变化趋势与生物多样性指数基本一致。

|
| 图 13 2004—2018年长江口海域底栖生物多样性变化情况 Fig. 13 Variation of benthos biodiversity in the Changjiang River Estuary from 2004 to 2018 |
近15年夏季长江口海域底栖生物优势种如表 4所示, 第一优势种平均优势度范围0.017—0.096, 2015年最低, 2012年最高。以下13种底栖生物在不同年份成为过长江口海域优势种:背蚓虫(Notomastuslatericeus)、不倒翁虫(Sternaspissculata)、彩虹明樱蛤(Moerellairidescens)、短吻铲荚螠(Listriolobusbrevirostris)、短叶索沙蚕(Lumbrinerislatreilli)、钩虾(Gammarussp)、尖叶长手沙蚕(Magelonacincta)、欧文虫(Oweniafusformis)、奇异稚齿虫(Paraprionospiopinnata)、日本角吻沙蚕(Goniada japonica)、扇栉虫(Amphicteisgunneri)、双形拟单指虫(Cossurelladimorpha)、丝异须虫(Heteromastusfiliformis)和圆筒原盒螺(Eocylichnacylindrella)等。其中背蚓虫分别为2004、2005、2007年的第一优势种, 其他年份不再是优势种;2009—2018年丝异须虫连续10a都是优势种, 其中有8a为第一优势种, 且2012年优势度达到了15a中最高的0.096。底栖生物第一优势种优势度变化情况如图 14所示, 2004—2008年优势度总体平稳, 2009—2015年第一优势种优势度震荡幅度变大, 总体上略呈下降的趋势。
| 年份 Year |
优势种 Dominant species |
| 2004 | 背蚓虫、奇异稚齿虫、日本角吻沙蚕、不倒翁虫、圆筒原盒螺 |
| 2005 | 背蚓虫、短叶索沙蚕、双形拟单指虫 |
| 2006 | 双形拟单指虫、尖叶长手沙蚕/、背蚓虫 |
| 2007 | 背蚓虫、双形拟单指虫、钩虾SP.、尖叶长手沙蚕 |
| 2008 | 尖叶长手沙蚕 |
| 2009 | 丝异须虫 |
| 2010 | 彩虹明樱蛤/、双形拟单指虫、丝异须虫、欧文虫 |
| 2011 | 丝异须虫 |
| 2012 | 丝异须虫 |
| 2013 | 丝异须虫 |
| 2014 | 钩虾SP.、丝异须虫 |
| 2015 | 丝异须虫 |
| 2016 | 丝异须虫、扇栉虫 |
| 2017 | 丝异须虫 |
| 2018 | 丝异须虫、短吻铲荚螠 |

|
| 图 14 2004—2018年长江口底栖生物第一优势种优势度变化 Fig. 14 Variation of benthos first dominant species′ dominance index in the Changjiang River Estuary from 2004 to 2018 |
对长江口海域海洋生物(浮游植物、浮游动物、底栖生物)健康状态进行评价。近15年长江口海域生物健康指数情况如图 15所示, 各年份健康指数震荡幅度较大, 指数范围为11.0—33.3, 各年份平均值为21.9, 接近亚健康指数下限。年际分布来看, 2004、2005、2010—2014年共7年份海洋生物处于“亚健康”状态, 2006—2009、2015—2018年共8年份处于“不健康”状态。海洋生物健康指数在河口生态系统健康指数中权重占50%[16], 根据东海区海洋生态环境状况公报[8], 长江口生态系统长期处于亚健康状态, 其中海洋生物为亚健康或不健康状态应是影响河口生态系统健康的重要因素之一。

|
| 图 15 2004—2018年长江口海域海洋生物健康指数变化 Fig. 15 Variation of Marine biological health index in the Changjiang River Estuary from 2004 to 2018 |
按照河口生物健康指数评价标准[16], 分别评价生物健康指数的5个指标。夏季长江口海域浮游植物密度总体偏高, 年平均密度仅2004、2005年符合I类赋值要求, 2014年符合II类赋值要求, 其他年份均为III类, 高于评价标准上限。浮游动物密度总体偏低, 年平均密度仅2017年赋值为II类, 其余年份均为III类, 低于评价标准下限;与之相反, 浮游动物生物量总体偏高, 2004、2005、2009、2010、2012、2013共6个年份年平均生物量符合I类赋值要求, 2006、2011和2014年符合II类赋值要求, 其余6年均为III类, 且高于评价标准上限。底栖动物密度总体偏高, 仅2014年符合II类赋值要求, 其余14年均为III类, 且高于评价标准上限;底栖动物生物量总体偏低, 2004、2005和2010年符合I类赋值要求, 2007、2016、2018年符合II类赋值要求, 其余9年均为III类, 且低于评价标准下限。
3.2 对主要污染物的压力响应及相关性分析长江径流携带流经地区的工、农业废水和城市污水, 将大量生源要素(C、N、P等)、有机污染物、重金属等带入长江口海域[17], 通过长江径流进入河口的污染物总量对上海长江口污染贡献率约占90%以上[18]。本文分别对长江主要污染物入海总量、海域严重富营养化(富营养化统计方法按文献[19])面积比例、营养盐结构等与各生物指标(生物健康指数, 硅藻甲藻种类数比值, 浮游生物和底栖生物的密度、生物量、优势度、均匀度、丰富度、多样性指数等)进行相关性(Pearson相关性, 双尾, N=15, P < 0.05)分析, 探讨环境因子对海洋生物的影响。
3.2.1 污染物通量与生物指标的相关性选取了长江口徐六泾断面监测的DIN、DIP、石油类和重金属(Cu、Pb、Cd、Cr、Zn、Hg)等主要污染指标, 统计了各指标2004—2018年历年1—8月份的累积等标污染负荷(等标污染负荷统计方法按照文献[20])与各生物指标的相关性。结果表明, DIN、石油烃类入海通量与生物健康指数呈显著负相关关系, 相关系数r分别为-0.592和-0.661;DIP与底栖生物生物量呈显著负相关关系(r = -0.597), 与浮游动物生物量和丰富度呈显著正相关关系, 相关系数r分别为0.611和0.629;重金属与底栖生物优势度呈显著正相关关系(r=0.661)。
3.2.2 营养盐结构变化与生物指标的相关性统计各站位的N/P和Si/N均值与各生物指标的相关性。结果表明, N/P与浮游植物丰度呈显著负相关(r = -0.589), 但与浮游植物均匀度和多样性指数呈显著正相关, 相关系数r分别为0.605和0.631;Si/N与浮游植物多样性指数呈显著负相关(r = -0.573)。
3.2.3 富营养化面积与生物指标的相关性统计历年长江口海域严重富营养化面积比例, 与各生物指标进行相关性分析。结果表明, 严重富营养化面积比例与硅藻甲藻种类数比值、底栖生物优势度均呈显著正相关关系, 相关系数r分别为0.625和0.751。
3.2.4 相关性结果讨论从宏观上, 营养盐(N、P、Si)水平是生态系统的物质基础, 对营养盐与浮游植物(初级生产力)的相互关系的研究较多[10-11, 21-26], 但由于其影响机制复杂, 大部分研究都是基于其统计数据的相关性分析。本文结合前人的研究基础, 试图部分给出上述指标的相关性分析。
DIN、石油烃类入海通量与生物健康指数呈负相关关系, 可能与作为生源要素促进浮游生物过渡生长有关。20世纪80年代以来, 长江口海域水体中DIN和DIP快速增加[21], 导致水体富营养化严重, 促进了海区浮游植物生长, 生物量增加, 近15年浮游植物密度有所上升, 2004—2018年中大部分年限浮游植物密度超过海洋生物健康评价标准[16]。有研究表明[27-28], 海水中石油烃溶解组分, 特别是低浓度条件下, 由于石油烃可以为海洋浮游植物生长提供所需的部分碳源, 从而对海洋浮游植物的生长产生一定促进作用。因此, 营养盐与石油烃类的输入在促进浮游植物生长方面可能存在叠加效应。营养要素与浮游生物、底栖生物的相关关系还需要进一步探讨。
N/P和Si/N以及严重富营养化面积与各生物指标的相关性可能与研究海域的水文、水质、生物分布特性有关。长江口海域水质分布特点为口内、口门及近河口区域盐度低、悬浮物含量高, 水质浑浊, DIN与DIP浓度高(N/P也高), 富营养化严重;随离岸方向盐度逐渐升高, DIN与DIP浓度迅速降低, 富营养化现象逐渐消失[8]。浮游植物的分布也具有河口区特征, 主要硅藻种类高生物量区域分布在低盐高营养盐的近河口羽状锋区, 主要甲藻种类高生物量区域主要分布在海域相对高盐低浊度的羽状锋区和层化水体上部[29]。据此推断, 近河口区域N/P升高, Si/N下降[21], 可能抑制了低盐区部分硅藻的生长, 促进了高盐区域甲藻种类和生物量的增长, 导致了N/P与浮游植物丰度呈负相关, 与浮游植物均匀度和多样性指数呈正相关, Si/N则与浮游植物多样性指数呈负相关。这与王江涛等[10]研究的近30a来长江口海域过量的DIN和持续升高的N/P可能是造成这硅藻-甲藻藻种长期演变现象的主要原因的结论相一致。同理, 严重富营养化区域处于近河口区域, 以硅藻分布为主, 非富营养化区域的甲藻种类增多, 因此严重富营养化面积比例与硅藻甲藻种类数比值呈正相关关系。
重金属与底栖生物的相关性可能与其毒性效应有关。重金属作为一类持久性和生物累积性污染物, 是近海环境中最主要的污染物之一, 进入海洋的重金属污染物绝大部分富集在沉积物中[30], 部分种类底栖生物对重金属含量表现出较高的耐受能力, 有些耐受能力则较低[30], 最终耐受力高的生物种类以优势度表现出来, 即重金属与底栖生物优势度呈正相关。
4 小结本文对近15年长江口海域夏季海洋生物变化趋势及健康状况进行评价, 结果表明:
(1) 群落结构组成发生一定变化。近15年与20世纪90年代末相比, 浮游植物种类数有所减少, 但细胞密度有所上升;浮游动物种类数有所增加, 生物量明显增加;底栖生物种类数大幅增加, 但生物量有所下降。浮游植物以硅藻为主, 但甲藻占比有所增加, 2010年以来, 硅藻、甲藻群落结构进入了一个新的平衡状态。浮游动物中以节肢动物为主, 节肢动物中的主要类群桡足类占比有所下降, 2009—2018年, 节肢动物与腔肠动物占比均处于动态平衡状态。近15年底栖生物种类数有明显升高的趋势, 2015—2018年均超过110种。
(2) 生物多样性总体水平一般。浮游植物多样性指数总体较低, 丰富度和均匀度较差, 第一优势种的优势度较高。浮游动物多样性指数和丰富度指数总体优于浮游植物, 处于较好水平, 但多年呈现下降的趋势。底栖生物多样性水平一般, 丰富度和均匀度较差, 优势种渐趋单一。
(3) 海洋生物总体处于不健康状态。各年份平均健康指数为21.9, 接近亚健康指数下限, 其中7年份处于“亚健康”状态, 8年份处于“不健康”状态。影响生物健康的主要指标为浮游植物密度总体偏高, 浮游动物密度总体偏低、生物量总体偏高, 底栖动物密度总体偏高、生物量总体偏低。
(4) 生态系统变化与陆源主要污染物排放、营养结构变化和水体富营养化均具有一定的相关性。DIN、石油类入海通量与生物健康指数呈显著负相关关系(P < 0.05), DIP与底栖生物生物量呈显著负相关关系。N/P与浮游植物丰度呈显著负相关, 但与浮游植物均匀度和多样性指数呈显著正相关;Si/N与浮游植物多样性指数呈显著副相关。海域严重富营养化面积比例与硅藻甲藻种类数比值呈显著正相关关系。
致谢: 感谢上海海洋大学水产与生命学院方淑波老师给予的帮助。| [1] |
于仁成, 张清春, 孔凡洲, 周正熙, 陈振帆, 赵越, 耿慧霞, 代丽, 颜天, 周名江. 长江口及其邻近海域有害藻华的发生情况、危害效应与演变趋势. 海洋与湖沼, 2017, 48(6): 1178-1186. |
| [2] |
王孝程, 解鹏飞, 李晴, 张金勇, 李宏俊. 长江口海域生态环境状况及保护对策. 环境科学研究, 2020, 33(5): 1197-1205. |
| [3] |
池连宝.长江口及邻近海域低氧区的时空变化特征与关键过程研究[D].青岛: 中国科学院大学(中国科学院海洋研究所), 2019.
|
| [4] |
叶属峰.大型工程对长江河口近岸海域生态系统的影响及机理研究[D].上海: 上海交通大学, 2005.
|
| [5] |
刘守海, 张昊飞, 何彦龙, 彭小家, 秦玉涛, 季晓, 杨志. 基于河口生物完整性指数评价上海周边海域健康状况的初步研究. 生态环境学报, 2018, 27(8): 1494-1501. |
| [6] |
陈耀辉, 刘守海, 何彦龙, 秦玉涛, 季晓, 张昊飞, 徐韧. 近30年长江口海域生态系统健康状况及变化趋势研究. 海洋学报, 2020, 42(4): 55-65. |
| [7] |
周晓蔚, 王丽萍, 郑丙辉. 长江口及毗邻海域生态系统健康评价研究. 水利学报, 2011, 42(10): 1201-1208. |
| [8] |
国家海洋局东海分局. 2017年东海区海洋环境公报.[2019-12-26]. http://ecs.mnr.gov.cn/xxgk_166/xxgkml/hytj/202009/t20200918_17772.shtml
|
| [9] |
王金辉, 黄秀清, 刘阿成, 张有份. 长江口及邻近水域的生物多样性变化趋势分析. 海洋通报, 2004, 23(1): 32-39. |
| [10] |
王江涛, 曹婧. 长江口海域近50a来营养盐的变化及其对浮游植物群落演替的影响. 海洋环境科学, 2012, 31(3): 310-315. DOI:10.3969/j.issn.1007-6336.2012.03.002 |
| [11] |
黄海燕, 王秋璐, 许艳, 刘捷, 曲艳敏. 2004-2016年夏季长江口网采浮游植物及其影响因素. 海洋与湖沼, 2018, 49(2): 319-330. |
| [12] |
张正龙, 徐韧, 范海梅. 近20 a以来长江口生态监控区边滩湿地变化的研究. 海洋环境科学, 2008, 27(S1): 5-8. |
| [13] |
沈新强, 晁敏, 全为民, 王云龙. 长江河口生态系现状及修复研究. 中国水产科学, 2006, 13(4): 624-630. DOI:10.3321/j.issn:1005-8737.2006.04.018 |
| [14] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378-2007海洋监测规范.北京: 中国标准出版社, 2008.
|
| [15] |
徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13-15. |
| [16] |
中华人民共和国国家海洋局. HY/T 087-2005近岸海洋生态健康评价指南.北京: 中国标准出版社, 2005.
|
| [17] |
许秋寒, 钱佳欢, 陈钊英, 张海平, 陈玲. 长江口及毗邻海域水环境现状与污染防治对策. 中国发展, 2015, 15(3): 10-14. |
| [18] |
陈慧敏, 孙承兴, 仵彦卿. 上海海域污染源分析及控制对策. 水资源保护, 2011, 27(2): 70-74, 79-79. |
| [19] |
邹景忠, 董丽萍, 秦保平. 渤海湾富营养化和赤潮问题的初步探讨. 海洋环境科学, 1983, 2(2): 41-54. |
| [20] |
陈剑虹, 杨保华, 邓子云. 环境统计应用. 北京:化学工业出版社, 2005, 94-95. |
| [21] |
杨颖, 徐韧. 近30a来长江口海域生态环境状况变化趋势分析. 海洋科学, 2015, 39(10): 101-107. |
| [22] |
韩瑞, 陈求稳, 王丽, 汤新武, 沈新强. 基于生态通道模型的长江口水域生态系统结构与能量流动分析. 生态学报, 2016, 36(15): 4907-4918. |
| [23] |
尹艳娥, 沈新强, 蒋玫, 袁琪, 平仙隐, 徐亚岩, 韩金娣, 王云龙. 长江口及邻近海域富营养化趋势分析及与环境因子关系. 生态环境学报, 2014, 23(4): 622-629. |
| [24] |
杨东方, 高振会, 王培刚, 于子江, 石强. 营养盐Si和水温影响浮游植物的机制. 海洋环境科学, 2006, 25(1): 1-6. |
| [25] |
杨东方, 王凡, 高振会, 孙培艳, 石强. 长江口理化因子影响初级生产力的探索Ⅰ.营养盐限制的判断方法和法则在长江口水域应用. 海洋科学进展, 2005, 23(3): 368-373. |
| [26] |
杨东方, 王凡, 高振会, 陈永利, 谢利. 长江口理化因子影响初级生产力的探索Ⅱ.磷不是长江口浮游植物生长的限制因子. 海洋科学进展, 2006, 24(1): 97-107. |
| [27] |
张传松, 王修林, 石晓勇, 韩秀荣, 孙霞, 祝陈坚, 陆茸. 东海赤潮高发区COD和石油烃分布特征及其与赤潮发生关系的初步研究. 应用生态学报, 2003, 14(7): 1093-1096. |
| [28] |
Shi X Y, Wang X L, Han X R, Jiang Y, Zhu M Y, Chen S, Koshikawa H. Relationship between petroleum hydrocarbon and plankton in a mesocosm experiment. Acta Oceanologica Sinica, 2001, 20(2): 231-240. |
| [29] |
孔凡洲.长江口赤潮区浮游植物粒级结构、种类组成和色素分析[D].北京: 中国科学院研究生院, 2011.
|
| [30] |
贾海波, 胡颢琰, 唐静亮, 王益鸣, 柴小平. 长江口及其邻近海域表层沉积物中重金属含量对大型底栖生物的影响. 海洋环境科学, 2011, 30(6): 809-813. |