 2020, Vol. 40
2020, Vol. 40文章信息
- 封传红, 张梅, 马利, 白马吞珠, 黄冲, 沈丽, 李庆, 郭聪
- FENG Chuanhong, ZHANG Mei, MA Li, BAIMA Tunzhu, HUANG Chong, SHEN Li, LI Qing, GUO Cong
- 西藏飞蝗蝗蝻群集迁移特性及其群集效应
- Effects of migration and aggregation of Locusta migratoria tibetnsis nymphs
- 生态学报. 2020, 40(20): 7534-7542
- Acta Ecologica Sinica. 2020, 40(20): 7534-7542
- http://dx.doi.org/10.5846/stxb201908021634
-
文章历史
- 收稿日期: 2019-08-02
- 网络出版日期: 2020-08-27





2. 甘孜州植保植检站, 康定 626000;
3. 全国农业技术推广服务中心, 北京 100026;
4. 四川省种子站, 成都 610041;
5. 四川农业大学, 温江 611130;
6. 四川大学, 成都 610065
2. The Plant Protection Station of Ganzi Prefecture, Kangding 626000, China;
3. The National Agro-Tech Extension and Service Center, Beijing 100026, China;
4. The Seed Management Station of Sichuan Province, Chengdu 610041, China;
5. Sichuan Agriculture University, Wenjiang 611130, China;
6. Sichuan University, Chengdu 610065, China
西藏飞蝗Locusta migratoria tibetnsis Chen是我国青藏高原特有种, 主要危害青稞、冬小麦、牧草等[1], 近年来受全球气候变暖的影响, 西藏飞蝗潜在分布区域扩大, 高密度蝗群面积增加, 引起农作物和牧草绝产绝收, 生态环境恶化[2-3]。作为我国三大飞蝗之一, 西藏飞蝗蝗蝻具有群集迁移习性, 能将分散的为害集中变为灾害[4-5], 加之, 蝗蝻群集迁移路线难以预测, 常常造成监测不准, 防治被动情况。蝗蝻群体运动涉及到形态、生理生化多个方面。飞蝗种存在群居型、散居型2种生态型, 群居型和散居型在形态特征、生理机能、行为及体色等方面存在明显差异[6], 群居型总是寻求同类聚集而居, 而散居型则躲避同类, 行孤单生活; 群居型经常在白天进行迁飞扩散, 而散居型则要到夜晚才偶尔飞行[7]。田间飞蝗在行为、内分泌、聚集信息、食物、资源分布等因素的作用下, 种群中群居型比例增加, 大量蝗虫聚集在一起[8-9], 但西藏飞蝗蝗蝻在聚集阶段没有明确的迁移方向和稳定的迁移速度。聚集后各个蝗蝻相互影响, 整个蝗蝻群行动的方向、速度趋向一致, 产生了群集迁移[10]。即便遇到人为、地貌等因素的干扰, 其个体速度和方向仍然保持与群体一致。这种一致性产生是西藏飞蝗灾害形成的前提。
群集运动是动物社会行为之一, 蚂蚁、蜜蜂、鱼群等均有社会行为。蚂蚁群体在低密度下呈现近随机分布状态, 高密度下为显著聚集状态[11]。在实验室环形通道中运动的沙漠蝗蝻随着通道中蝗蝻数量的增加, 表现出高度一致运动特性[10]。模拟显示, 鱼群只要其个体遵守3条运动规则, 就可以形成有序的群体[12]。复杂理论表明, 由单个成员组成的群体, 可以通过自组织, 显示出在个体层次上不能出现的多种复杂行为[13], 正是这些行为可以增加物种适应能力[14]。昆虫存在有18种不同类型的群集形式, 其形成功能可分为防御、牵引结构、温度调节、恶劣条件下的群体生存和穿越障碍物5种, 对于某个群集来说这些功能并非互斥的, 可能同时具备[15]。红火蚁群集体具有防水、粘弹性等物理特性, 这种性质是由个体活动相互连接产生的[16]。蚂蚁群体总能发现一条最短的路找到食物, 蜂群对优质蜜源的选择, 都是自组织昆虫群集形成的功能[17]。沙漠蝗群集迁移时会从水底爬过, 迁飞蝗群能够避免飞越水面, 这减少了蝗群落入海水中的危险[18-19]。自然条件下西藏飞蝗蝗蝻群集迁移可以看作是一个从无序到有序的自组织过程。对西藏飞蝗乃至飞蝗群集迁移的研究, 大多采取观察方法, 缺乏利用自组织原理分析蝗虫群集形成机制, 深入到飞蝗群集效应方面的研究。另一方面阐明西藏飞蝗蝗蝻群集迁移特性、形成机制及其效应, 为利用预防措施扰乱其群集迁移、将蝗虫灾害控制在群集迁移之前提供了可能性。
本文利用川西高原暴发西藏飞蝗蝗蝻群视频资料, 通过轨迹跟踪等方法, 获取蝗蝻群集运动参数, 分析蝗蝻群集运动规律、迁移特性和群集效应, 旨在从系统学角度发现西藏飞蝗灾变行为机制, 评估西藏飞蝗蝗蝻群迁移能力及成灾潜力, 为西藏飞蝗的监测预警和防治提供科学依据。
1 材料与方法 1.1 蝗蝻群集迁移视频采集采集环境。标准视频采集:2009年7月在四川省甘孜州炉霍县宜木乡进行了群居型蝗蝻群拍摄, 蝗蝻密度平均为50—220头/m2, 经度100°77′E, 纬度31°30′N, 海拔3100—3200 m。选取3类型场景:已经翻耕农田, 前茬为青稞, 植被覆盖率在5%以下; 草地, 植被覆盖率50%—60%;沙滩, 植被覆盖率在5%以下。其他视频:2006年7月四川省甘孜州甘孜县卡功乡发生高密度群居型西藏飞蝗, 经度99°73′E, 纬度31°67′N, 海拔3300—3500 m, 蝗蝻从周围草场迁入农田, 农田已经翻耕, 前茬为青稞, 密度为30—400头/m2。
标准视频采集方法:用支架将摄像机垂直竖起, 摄像机镜头距地面1.4—1.8 m, 地面水平放置1根直尺, 设置摄像机镜头的取景大小, 荧屏高度为0.5 m; 拍摄不同的环境下蝗蝻群, 每个点拍摄5—10 min, 拍摄时摄像机保持静止; 拍摄时人员尽量远离蝗蝻群, 避免干扰; 摄像机采用数码摄像机, 下载速度每秒25帧。其他视频:选择手持拍摄的抖动相对较轻的视频序列分析蝗蝻群集运动规律。
1.2 蝗蝻群运动一致性参数采用计算群体平均矢量速度的方法来衡量蝗蝻运动的一致性程度[20], 建立序参数公式(do)。序参数越接近1, 蝗蝻运动一致性越高。
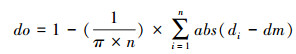
|
式中, n为视野的蝗蝻数, di为第i个蝗蝻的瞬时方向, dm为n个蝗蝻同时刻平均方向, 方向范围[0, 2π)。
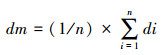
|
采用自推进粒子模型(Self-propelled particles model, SPP)来模拟蝗蝻群的运动[20]。SPP模型规则:迁移蝗蝻运动方向由其邻近蝗蝻决定。
(1) 假定在开始时期(t=1)蝗蝻速度(V)恒定, 方向(θ)随机。
(2) 任意时刻, 蝗蝻i的位置Xi(t)为:
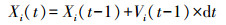
|
(3) 任意时刻, 蝗蝻运动方向θi(t)为影响距离(R)内n个蝗蝻运动方向的平均值。
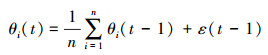
|
式中, ε为随机变量, 相当于地形、植被等环境因子对蝗蝻运动的干扰。
1.4 数据分析和处理采用OpenCV进行蝗蝻视频轨迹分析。平均数、标准差等统计值的计算、SPP模型设计和图形的制作均采用MathWorks公司Matlab R2014a软件进行。
2 结果和分析 2.1 自然环境中西藏飞蝗蝗蝻运动 2.1.1 蝗蝻群集迁移速度在高密度蝗蝻群集迁移时, 单个蝗蝻以爬行和短距离跳跃为主, 一般爬行7—13帧再短距离跳跃3—5帧, 也有连续跳跃的, 长距离跳跃主要是应急时发生。蝗蝻群集迁移平均速度为0.1256 ±0.0784 m/s(数值为平均值±标准误差)。
2.1.2 蝗蝻群集迁移方向选取稳定迁移的蝗蝻群的位移资料, 将视野中蝗蝻两两距离与移动方向差作散点密度图。图 1表明, 坐标原点(0, 0)密度最大, 群集中个体之间距离越近, 个体间方向趋向越一致。在0.2 m的范围内, 各蝗蝻移动方向差在1.3963 rad内。

|
| 图 1 群集迁移蝗蝻群中个体之间运动方向差与距离散点密度图 Fig. 1 Density map of normalizing movement direction difference and distance among migrating locust nymph individuals in the fields 蝗蝻方向差为绝对值, 范围为[0, π)。个体之间距离和方向差按(x-min)/(max-min)公式进行标准化处理 |
(1)蝗蝻群运动轨迹
草地、农田和沙滩3种环境西藏飞蝗蝗蝻群运动轨迹表明, 蝗蝻群沿着一定路线向前迁移, 相邻蝗蝻运动轨迹接近平行(图 2)。草地运动轨迹相对分散。

|
| 图 2 农田、草地和沙滩西藏飞蝗蝗蝻移动轨迹 Fig. 2 Movement tracks of Locusta migratoria tibetnsis Chen nymphs in farmland, grassland and sandbeach |
(2) 蝗蝻群运动一致度
蝗蝻群在农田、草地、沙滩运动一致性较强, 但不同环境一致性程度不同, 其序参数按照沙滩、农田、草地依次降低(图 3)。农田蝗蝻群序参数为0.7870±0.0829(数值为平均值±标准误差), 草地蝗蝻群序参数0.6987±0.1384, 沙滩蝗蝻群序参数为0.8502±0.0940。

|
| 图 3 农田、草地、沙滩西藏飞蝗蝗蝻运动序参数 Fig. 3 Normalized alignment of migrating movement of Locusta migratoria tibetnsis Chen nymphs in farmland, grassland and sandbeach |
根据前述田间观测, 西藏飞蝗蝗蝻群集迁移SPP模型参数设置见表 1。
| 参数Parameters | 值Value | 备注Remark |
| 速度(V)Speed | 0.1 m/s | 实测速度 |
| 影响距离(R)Interaction radius | 0.2 m | 实测 |
| 噪音(ε)Noise | 0.2 | 影响距离内蝗蝻平均方向差/(2π) |
| 边界条件Boundary condition | 周期边界 | 当蝗蝻从某边跑出模拟场景, 立即从对边相同位置以相同速度进入场景 |
| SPP:自推进粒子模型(Self-propelled particles model) | ||
模拟表明, 密度较高的蝗蝻群(100头/m2)在初始时方向随机, 经过逐步局部的调整, 使整体运动趋向一致(图 4)。但是密度较低的蝗蝻群不会出现运动一致的情况。

|
| 图 4 蝗蝻群运动由初始无序状态转变为有序状态 Fig. 4 The transition from the beginning disorder to finally order of locust nymph group 模拟蝗蝻群密度为100头/m2, 初始时期t=1时, 方向随机, 模拟结束时间为t=2000 |
不同密度蝗蝻群序参数有明显的变化, 密度越高蝗蝻运动序参数越高, 当密度超过12—15头/m2, 序参数曲线平稳, 可以稳定在0.8以上, 接近或者超过农田、沙滩或草地观测值, 此时方差在0.05左右, 表明系统进入稳定的有序状态(图 5)。

|
| 图 5 密度为1—20头/m2蝗蝻群序参数的变化 Fig. 5 Normalized alignment of movement of locust nymph group of density from 1—20 locusts/m2 |
(1) 群集对迁移距离的影响
在低密度时, 蝗蝻群序参数较低, 蝗蝻个体没有明确的定向机制, 随机游走。高密度蝗蝻群运行方向比较稳定, 群集迁移的距离明显远于随机游走的蝗蝻。图 6中1头/m2蝗蝻群移动距离平均为9.1896 m, 100头/m2蝗蝻群移动距离平均为26.7909 m。随着模拟时间的加长, 未群集的蝗蝻始终围绕原点附近扩散, 而群集的蝗蝻群位置距原点越来越远。

|
| 图 6 密度为1头/m2、100头/m2蝗蝻群的轨迹图 Fig. 6 Movement tracks of locust nymph group of 1 and 100 locusts/m2 模拟蝗蝻数均为100头, 时间步长为300 |
(2) 蝗蝻群对食物或有益资源的定向
轨迹图说明蝗蝻群开始未趋向食物, 但通过群集运动, 所有个体最后找到食物位置(图 7)。蝗蝻群运动方向的频次分布说明蝗蝻群在最初1—70个时间步里整个群体一直在调整方向, 而后群体内所有个体均趋向食物位置运动, 运动方向不再变化, 至120时间步左右, 找到食物(图 7)。随机运动蝗蝻群模拟表明, 序参数在0.2—0.3以下, 寻食轨迹混乱, 只有部分蝗蝻能够找到食物。

|
| 图 7 蝗蝻群寻找食物过程中轨迹和方向分布 Fig. 7 Movement tracks and direction distribution of locust nymphs group looking for food |
方向分布是将360度分为40个区间, 统计各个区间运动方向的蝗蝻占百分比。蝗蝻群密度为100头/m2, 初始蝗蝻随机分布在x∈[0, 1]和y∈[0, 1]的区间, 任意设定点状食物坐标为(8, 8)。假设食物信息随距离的增加而衰减, 符合高斯分布模型:f(r)=a1×exp(-|r-b1|/c1)2), r为某个蝗蝻的坐标, b1为食物坐标(8, 8), c1为标准差(c1=3), a1为给定常数, a1=20
(3) 先验蝗蝻对群体作用
模拟表明先验个体可引导蝗蝻群迁移(图 8), 先验蝗蝻引导整个蝗蝻群集迁移成功率与先验蝗蝻的比例有关, 比例越高, 成功引导蝗蝻群运动的几率越大。先验蝗蝻比例达到5%, 引导成功率达80%以上。

|
| 图 8 1%—9%先验蝗蝻对蝗蝻群集迁移引导效果 Fig. 8 The guidance of experienced nymphs of 1%—9% the populationnumber to migration of locust nymph group 先验蝗蝻指方向固定以pi/4的角度直线运动的蝗蝻, 9个蝗蝻群密度均为100头/m2 |
田间观察西藏飞蝗蝗蝻在温度过低时不发生迁移, 此外降雨和大风天气也不迁移。为了减少温度、光照等差异对群集迁移的影响, 本文于7月上旬采集视频, 蝗蝻为3—5龄, 晴天或多云、无风或微风天气进行观测, 拍摄时间为10:00—13:00, 气温为17—22℃, 此时间段为蝗蝻群稳定迁移期。2个视频采集地点均位于雅砻江流域河谷, 环境条件一致, 沙滩土质为沙土至沙壤土, 是西藏飞蝗发生源头; 草地为退化草地, 植被稀矮, 覆盖率较低, 土质为砂砾; 农田为青稞收割后翻耕的轮休地。本文中沙滩、草地、农田相互交错, 为同一气候条件。
3.2 西藏飞蝗蝗蝻群集效应西藏飞蝗蝗蝻群集迁移一般发生在9:00—16:00, 按照本文计算, 1 d西藏飞蝗蝗蝻群集迁移的最大距离约为2.5 km, 而随机扩散距离约为70—80 m。雅砻江河谷地带为狭长地带, 蝗蝻群集迁移15 d可以从河滩等虫源地覆盖河谷大部分地区。群集迁移减少了个体的随意漫游, 加之群居型飞蝗的运动速度高于散居型[21], 相对于散居型, 这进一步增加了群居型群集迁移距离。蝗蝻群对食物的定向作用, 在对远距离食物定向中更为突出, 图 7食物源在坐标(8, 8)时, 随机游走蝗蝻群也有70%左右能发现食物, 当将食物源移到坐标(30, 30), 实验100次, 随机游走蝗蝻只有1.8%的个体能发现食物, 而蝗蝻群集迁移找到食物的成功率是13%, 为随机游走蝗蝻的7.2倍。西藏飞蝗发生于青藏高原, 在高原海拔、温度、寄主植物、氧气含量、紫外线等环境因子的影响下, 西藏飞蝗获取食物等生命活动更为困难[22-26]。西藏飞蝗蝗蝻通过群集迁移能够取得分散个体不能获取的在迁移距离和寻找食物上的效应, 补偿了西藏飞蝗作为昆虫在语言、视觉、嗅觉等通讯能力的不足[27], 有利于其适应高原恶劣环境, 实现种群繁衍[5, 28]。
3.3 西藏飞蝗的防治指标本文从行为角度得到蝗蝻群集迁移形成的临界密度为12—15头/m2, 在此密度之下, 蝗蝻是局限性的分散危害, 难以形成集中灾害。从经济阈值的角度, 西藏飞蝗在青稞、牧草、小麦等上的防治指标均在5头/m2以内[1, 29]。由于青藏高原缺氧, 地形复杂, 行动不便, 用工费用高上, 防治指标严, 对防治成本消耗过大。再者, 用传统的经济阈值计算西藏飞蝗的防治指标没有考虑到生态成本, 在青藏高原更应该考虑到尽量少使用农药等。因此本文建议适当放宽西藏飞蝗防治指标。
3.4 群集迁移的群体决策对于成群结队觅食或迁移的动物, 运动决策通常取决于群体成员之间的互动[30], 美洲鳊鱼群体中少数有经验的可以引导整个鱼群找到食物[31]。目前还没有观察到自然条件下蝗蝻群领头者, 似乎民主决策更为合适[32]。本文模拟显示, 蝗蝻群的先验者的存在可以极大地影响群体运动, 而且先验者只需占极少的比例即可, 这与蟑螂群体寻找食物的决策过程类似[33]。近来研究表明, 蝗虫可以记忆群集效应长达24 h[34], 如果蝗蝻能够记忆其他的有益信息, 那么对于整个蝗蝻群来说将会获得巨大利益。
| [1] |
杨群芳, 廖志昌, 李庆, 杨刚, 封传红, 蒋春先. 西藏飞蝗食性及其防治指标. 植物保护学报, 2008, 39(5): 399-404. |
| [2] |
Feng C H, Guo C, Luo L M, Qin Z. Response of Locusta migratoria tibetensis Chen (Orthoptera:Acrididae) to climate warming over the Tibetan plateau. Journal of Applied Entomology, 2012, 136(4): 313-320. DOI:10.1111/j.1439-0418.2011.01659.x |
| [3] |
苏红田, 白松, 姚勇. 近几年西藏飞蝗的发生与分布. 草业科学, 2007, 24(1): 78-80. |
| [4] |
唐昭华, 王保海, 王成明, 袁维红. 西藏飞蝗蝗蝻的习性. 西藏农业科技, 1992(1): 37-40. |
| [5] |
封传红, 陈淑群, 郭聪, 张跃进, 刘万才, 罗林明. 蝗虫的聚集行为机制研究进展. 植物保护, 2014, 40(1): 16-20. |
| [6] |
朱道弘, 阳柏苏. 飞蝗变型及体色多型的内分泌控制机理. 动物学研究, 2004, 25(5): 460-464. |
| [7] |
Hoste B, Sas F, Vandersmissen T, De Loof A, Breuer M, Huybrechts J. A miniaturized assay to quantify effects of chemicals or physical stimuli upon locust activity. Insect Science, 2006, 13(5): 401-407. DOI:10.1111/j.1744-7917.2006.00109.x |
| [8] |
Hassanali A, Njagi P G N, Bashir M O. Chemical ecology of locusts and related acridids. Annual Review of Entomology, 2005, 50: 223-245. DOI:10.1146/annurev.ento.50.071803.130345 |
| [9] |
Despland E, Simpson S J. Resource distribution mediates synchronization of physiological rhythms in locust groups. Proceedings of the Royal Society B, 2006, 273(1593): 1517-1522. DOI:10.1098/rspb.2006.3471 |
| [10] |
Buhl J, Sumpter D J T, Couzin I D, Hale J J, Despland E, Miller E R, Simpson S J. From disorder to order in marching locusts. Science, 2006, 312(5778): 1402-1406. DOI:10.1126/science.1125142 |
| [11] |
段青玲, 李耕, 姜立, 狄增如, 韩战钢. 密度对蚁群自组织聚集的影响. 北京师范大学学报:自然科学版, 2017, 53(3): 281-287. |
| [12] |
Hemelrijk C K, Kunz H. Density distribution and size sorting in fishschools:an individual-based model. Behavioral Ecology, 2005, 16(1): 178-187. |
| [13] |
Parrish J K, Edelstein-Keshet L. Complexity, pattern, and evolutionary trade-offs in animal aggregation. Science, 1999, 284(5411): 99-101. DOI:10.1126/science.284.5411.99 |
| [14] |
Sumpter D J T. The principles of collective animal behaviour. Philosophical Transactions of the Royal Society B, 2006, 361(1465): 5-22. DOI:10.1098/rstb.2005.1733 |
| [15] |
Anderson C, Theraulaz G, Deneubourg JL. Self-assemblages in insect societies. Insectes Sociaux, 2002, 49(2): 99-110. DOI:10.1007/s00040-002-8286-y |
| [16] |
Tennenbaum M, Liu Z Y, Hu D, Fernandez-Nieves A. Mechanics of fire ant aggregations. Nature Materials, 2016, 15: 54-59. DOI:10.1038/nmat4450 |
| [17] |
Bonabeau E, Theraulaz G, Deneubourg J L, Aron S, Camazine S. Self-organization in social insects. Trends in Ecology & Evolution, 1997, 12(5): 188-193. |
| [18] |
Heitler W J, Mitchell J L, Dinwiddie L. Underwater locomotion in the desert locust:behavioural choice when confronted with an aquatic barrier. Journal of Insect Behavior, 2005, 18(5): 669-683. DOI:10.1007/s10905-005-7018-3 |
| [19] |
Shashar N, Sabbah S, Aharoni N. Migrating locusts can detect polarized reflections to avoid flying over the sea. Biology Letters, 2005, 1(4): 472-475. DOI:10.1098/rsbl.2005.0334 |
| [20] |
Czirók A, Stanle H E, Vicsek T. Spontaneously ordered motion of self-propelled particles. Journal of Physics A:Mathematical and General, 1997, 30(5): 1375-1385. DOI:10.1088/0305-4470/30/5/009 |
| [21] |
郭志永, 石旺鹏, 张龙, 王贵龙. 东亚飞蝗行为和形态型变的判定指标. 应用生态学报, 2004, 15(5): 859-862. |
| [22] |
朱昱翰, 李庆, 杨刚, 白马吞珠, 匡健康, 蒋春先, 王海建, 杨群芳. 低温和光周期对西藏飞蝗体内物质的影响. 应用生态学报, 2016, 27(2): 629-633. |
| [23] |
李庆, 吴蕾, 杨刚, 匡健康, 封传红, 罗怀海, 杨群芳, 蒋春先, 王海建. 温度和紫外辐射胁迫对西藏飞蝗抗氧化系统的影响. 生态学报, 2012, 32(10): 3189-3197. |
| [24] |
李庆, 廖志昌, 杨刚, 封传红, 杨群芳, 罗怀海, 蒋春先, 王海建. 植被及土壤对西藏飞蝗产卵的影响. 中国农业科学, 2012, 45(4): 656-665. |
| [25] |
李庆, 王思忠, 封传红, 张敏, 蒋凡, 杨刚, 罗林明. 西藏飞蝗(Locusta migratoria tibetensis Chen)耐寒性理化指标. 生态学报, 2008, 28(3): 1314-1320. |
| [26] |
封传红, 单绪南, 郭聪, 罗林明. 1961-2005年西藏飞蝗潜在分布的变化. 昆虫学报, 2011, 54(6): 694-700. |
| [27] |
尚玉昌. 昆虫的感觉和通讯. 生物学杂志, 1986(3): 43-46. |
| [28] |
Chen Y L, Zhang D E. Historical evidence for population dynamics of Tibetan migratory locust and the forecast of its outbreak. Insect Science, 1999, 6(2): 135-145. DOI:10.1111/j.1744-7917.1999.tb00160.x |
| [29] |
唐昭华, 王保海, 王成明, 代万安. 西藏飞蝗为害麦作的防治指标探讨. 西藏农业科技, 1994, 15(1): 18-21. |
| [30] |
Couzin I D, Krause J, Franks N R, Levin S A. Effective leadership and decision-makingin animal groups on the move. Nature, 2005, 433(7025): 513-516. DOI:10.1038/nature03236 |
| [31] |
Reebs S G. Can a minority of informed leaders determine the foraging movements of a fish shoal?. Animal Behaviour, 2000, 59(2): 403-409. DOI:10.1006/anbe.1999.1314 |
| [32] |
Conradt L, Roper T J. Group decision-making in animals. Nature, 2003, 421(6919): 155-158. DOI:10.1038/nature01294 |
| [33] |
Amé J M, Halloy J, Rivault C, Detrain C, Deneubourg J L. Collegial decision making based on social amplification leads to optimal group formation. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(15): 5835-5840. DOI:10.1073/pnas.0507877103 |
| [34] |
Geva N, Guershon M, Orlova M, Ayali A. Memoirs of a locust:density-dependent behavioral change as a model for learning and memory. Neurobiology of Learning and Memory, 2010, 93(2): 175-182. DOI:10.1016/j.nlm.2009.09.008 |