 2020, Vol. 40
2020, Vol. 40文章信息
- 张琦, 李浙, 吴庆明, 隋媛
- ZHANG Qi, LI Zhe, WU Qingming, SUI Yuan
- 河南民权湿地公园青头潜鸭越冬行为模式及性别差异
- Overwintering behavior pattern and sex difference of Aythya baeri in Henan Minquan Wetland Park
- 生态学报. 2020, 40(19): 7054-7063
- Acta Ecologica Sinica. 2020, 40(19): 7054-7063
- http://dx.doi.org/10.5846/stxb201909131908
-
文章历史
- 收稿日期: 2019-09-13
- 网络出版日期: 2020-07-31

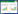




行为时间分配与活动节律是野生动物最基本的行为内容, 也是野生动物行为学研究最基础最重要的部分, 不仅能反映野生动物的生活习性、生理需要、生态需求、种内种间关系等[1-3], 还可以了解野生动物的生存状态、行为策略以及社会地位差异等, 对制定野生动物及其栖息地资源的保护管理措施有着重要的参考价值[4-5]。越冬期是候鸟完成生活史的关键时段, 食物供给性、可获得性、人为干扰等因素对其能否顺利越冬非常关键, 所有这些均通过行为模式进行体现[6-7]。
青头潜鸭(Aythya baeri)隶属雁形目(ANSERIFORMES)鸭科(Anatidae)。2012年世界自然保护联盟(IUCN)将其提升为极危鸟种[8], 成为中国54种鸭类中两种极危(CR)的鸭种之一。全世界种群数量不足1000只, 繁殖区域缩小为俄罗斯东南部及中国的华中和华北地区[9], 越冬区缩小为中国中部至缅甸区间的亚洲南部[10-11]。我国种群数量约为600—800只, 是青头潜鸭的重要分布地[11]。历史上, 青头潜鸭曾与红头潜鸭(Aythya ferina)是分布于东亚—澳大利西亚候鸟迁徙路线上中国区域内的常见潜鸭[12-15], 松嫩平原曾是其主要繁殖地。近年来, 国内外学者加大了对青头潜鸭的关注力度, 有关青头潜鸭的繁殖地、迁徙停歇地、越冬地的信息已经受到关注[10-11, 16-25];其中, 河南民权湿地公园是其中一个特殊的分布地, 该分布地于2017年首次记录到青头潜鸭, 2018年冬季观察期间观测到的最大群体超过150只(本文观察为158只, 人民网报道数量为155只[26]), 2019年9月7日该公园通过新闻发布会确认为“青头潜鸭重要繁殖地”, 这意味着青头潜鸭在该公园属于留鸟, 这对青头潜鸭世界种群的保护具有决定性的作用。
基于此背景, 本研究选择民权湿地公园, 对已发现的青头潜鸭群体进行了初步观察, 并对越冬期行为模式及性别差异进行了初步研究, 以期掌握该公园青头潜鸭越冬期的行为规律, 为青头潜鸭累积最新生物学信息, 为更进一步的保护管理提供科学依据。
1 研究地概况研究区位于河南民权湿地公园(下文简称民权湿地公园)。该公园位于河南省商丘市民权县东北部6 km处, 地处豫东平原东北部, 地理坐标为115°11′56″—115°25′41″E, 34°37′16″—34°42′48″N, 南北宽3 km、东西长30 km, 总面积2303.5 hm2, 其中湿地面积2266.5 hm2, 湿地覆盖率达到98.39%, 平均水深约3 m。该公园所在区域处于北温带, 受季风影响, 属于典型的湿润半湿润大陆性季风气候区, 年均温14.1℃, 地势相对平坦, 海拔30—70 m, 年均降水量为674 mm。该公园共统计到鸟类16目37科121种, 其中国家Ⅰ、Ⅱ级重点保护鸟类有23种, 如青头潜鸭、大天鹅(Cygnus cygnus)、小天鹅(Cygnus columbianus)、灰鹤(Grus grus)、鸳鸯(Aix galericulata)、大鸨(Otis tarda)等。
民权湿地公园是河南商丘黄河故道的重要组成部分, 其辖区范围北至西张庄、西至马庄、南至王庄、东至吴屯大坝, 主要由鲲鹏湖(任庄水库)、秋水湖和龙泽湖3个相互连接的水库组成, 水库内部有很多当地农民围堰而成的鱼塘, 鱼塘内的藕丛为青头潜鸭的生存提供了合适的栖息环境和越冬食物。因此, 鱼塘衍生的直接和间接人为活动与青头潜鸭的越冬栖息存在一定的时空重叠, 对青头潜鸭的越冬行为一定的影响。
2 研究方法 2.1 野外观察本研究野外数据收集于2018年11—12月。
首先采用样线法对民权湿地公园进行了整体性调查, 了解掌握青头潜鸭在该公园的栖息地空间分布情况, 来确定最佳观察群体, 整个公园区域, 观察期间, 共观测到不同区域的群体数量为158只, 这高于人民网报道的155只[26];其次, 在综合筛选最佳观察群体的基础上确定最佳观察区域;然后, 针对最佳观察区域选取最佳观察点, 最佳观察地点的选取原则以不干扰青头潜鸭的日常行为活动、群体数量相对最多、最利于观察为前提, 最终选择鲲鹏湖的5个观察点作为本次野外数据收集工作的最佳观察区域和最佳观察点(图 1), 基本能观察区域内青头潜鸭的分布范围。由于青头潜鸭多活动于藕丛深处, 自身隐蔽性较强, 故观察者选择位于距青头潜鸭约170—200 m的河堤上进行观察。

|
| 图 1 青头潜鸭越冬行为观察区位图 Fig. 1 Observation site of wintering behavior for Baer′s Pochard |
观察分为预观察(时间为3 d)和正式观察两个阶段(时间为31 d)。预观察前根据其他鸭种的行为学研究拟定青头潜鸭越冬行为谱[27-30];进入预观察后, 根据实际观察的青头潜鸭越冬期昼间行为对拟定行为谱进行补充、修改及完善, 并对观察频率间隔和观察时间进行选定, 最终定为5 min观察一次, 根据当地日出日落时间、光线可视度及雾霾影响, 将每天的越冬行为观察时间定为08:30—17:30;同时熟悉青头潜鸭的快速识别与快速定位。
正式观察时, 采用瞬时扫描法进行行为观察取样, 通过单筒望远镜(20到60倍)每隔5 min对视野内的青头潜鸭扫描观察一次行为, 通过双筒望远镜(8×42)观察记录视野内飞行的青头潜鸭群体数量。观察区域的最大群体数量为139只。
实际观察过程中, 由于渔船作业干扰、雾气导致的可视度影响, 每天的有效行为观察时间为09:30—17:30, 共收集数据43331条, 有效数据35633条, 有效率为82.23%。
2.2 越冬行为谱通过拟定、预观察、正式观察等渐进步骤后形成民权湿地公园青头潜鸭越冬行为谱, 分为觅食(Foraging)、运动(Locomotion)、修整(Maintain)、静息(Resting)、飞行(Flying)、社会(Social)和特殊(Special)7大类17种。
(1) 觅食行为类别, 包括取食(本文指以获取水面食物为主, 将喙部贴在水面或深入水下获取食物的过程)、喝水(指喙部进入水中、获取水、并仰头咽下水的一系列动作)、潜食(潜食行为的活动位置与取食行为的活动位置互补, 指将整个身体全部潜入水中获取食物、浮出水面时带有食物痕迹的过程)3种。
(2) 静息行为类别, 包括睡眠(指头颈后转、喙前端埋于翅下、闭眼静止不动的行为)、静息(指个体头颈略为回缩、静止不动的行为)2种。
(3) 修整行为类别, 包括理羽(指用喙部梳理、修饰身体不同部位羽毛羽序的一系列过程)、挠头(指头斜向后转, 抬起脚蹼挠动头部的动作)、抖翅(指躯干部于水中接近直立并快速扇动、拍打翅膀的行为)、洗澡(指个体于水体表面, 通过颈部伸缩、翅膀的张合扇动及身体摆动等动作将水遍布身体背面、净化身体的过程)、伸展(指个体于水中或潜水中, 颈部前伸、跗蹠后伸、翅膀展开等一系列舒展身体的行为, 该行为多发生在静息后)5种。
(4) 运动行为类别, 包括游泳(指个体于水体表面或急或缓的游动过程)、嬉戏(指个体于水中不同位置剧烈游动的一系列过程, 该行为为单只个体发生的行为, 不同于洗澡与游泳)。
(5) 飞行行为类别, 仅飞行1种(指身体飞离水面在一定高度的空间发生位移的过程, 包括主动飞行、惊扰飞行)。
(6) 社会行为类别, 包括内赶(指种内个体两只或两只以上发生相互追赶、攻击的行为)、外赶(指与种间鸟类相互追赶、抢食、打斗的行为)2种。
(7) 特殊行为类别, 包括警戒(指对周围干扰产生的头颈伸直向多个方向持续观看的过程)、特殊(指频次极少的行为如鸣叫, 难以准确定义的行为如上下颠动脖子等)2种。在实际观察中, 青头潜鸭警戒行为发生较少, 当周边有剧烈声响、渔船经过或渔民进行作业时, 表现为与周围其他水鸟一起飞离该区域, 观察记录时记做飞行行为。当周围有大种群其他鸟类飞行时, 青头潜鸭表现为短暂的警戒行为, 随后恢复之前的行为, 由于警戒行为极短且发生次数较少, 故观察过程中观测的频率较低, 归类在特殊行为类型中。
2.3 数据处理通过Excel软件对数据进行了统计, 同时结合SPSS 24.0中的卡方检验、Kolmogorov-Smironov检验、Pearson相关性检验、Spearman相关性检验、Cluster Analysis等方法对青头潜鸭越冬行为时间分配和活动节律及雌雄不同性别间的越冬行为进行了分析, 用Sigmaplot 10.0制作了行为时间分配及行为节律差异图。所有数据均以均值±标准误(M±SE)表示。
3 结果与分析 3.1 越冬行为模式 3.1.1 越冬行为时间分配民权湿地公园, 青头潜鸭的7类越冬行为中(图 2):静息(28.15%±4.15%)、觅食(21.66%±1.62%)、运动(21.09%±1.71%)三类行为是主要行为;其次是修整(13.78%±0.97%)和飞行(12.65%±2.82%)两类行为;社会和特殊两类行为所占时间分配最少。

|
| 图 2 青头潜鸭越冬行为时间分配 Fig. 2 Time allocation of wintering behavior for Baer′s Pochard |
对民权湿地公园青头潜鸭越冬行为时间分配进行了Spearman相关性分析, 结果表明(表 1):运动行为与觅食、修整、社会等行为之间呈极显著正相关(r=0.282/0.559/0.444, df=131, P=0.001), 修整行为与社会行为呈极显著正相关(r=0.253, df=131, P=0.000);静息行为与修整、运动、社会等行为之间呈极显著负相关(r=-0.308/-0.515/-0.372, df=131, P=0.000), 飞行行为与觅食、静息、修整等行为之间呈极显著负相关(r=-0.332/-0.403/-0.240, df=131, P=0.000), 其他行为时间分配之间均不具有显著相关性(P>0.05)。
| 觅食 Foraging | 静息 Resting | 修整 Maintain | 运动 Locomotion | 飞行 Flying | 社会 Social | 特殊 Special | |
| 觅食 Foraging | 1.000 | -0.170 | 0.159 | 0.282** | -0.332** | -0.003 | 0.130 |
| 静息 Resting | — | 1.000 | -0.308** | -0.515** | -0.403** | -0.372** | 0.058 |
| 修整 Maintain | — | — | 1.000 | 0.559** | -0.240** | 0.253** | -0.078 |
| 运动 Locomotion | — | — | — | 1.000 | -0.188* | 0.444** | -0.110 |
| 飞行 Flying | — | — | — | — | 1.000 | 0.088 | -0.020 |
| 社会 Social | — | — | — | — | — | 1.000 | 0.006 |
| 特殊 Special | — | — | — | — | — | — | 1.000 |
| *示0.05的显著水平, **示0.01的显著水平 | |||||||
民权湿地公园, 青头潜鸭的7类越冬行为时间分配曲线中(图 3):作为主要行为的静息行为, 其行为时间分配曲线始终高于其他行为, 不存在峰期和谷期;觅食行为存在3个峰期(分别为10:30—11:30、13:30—14:30、15:30—17:30)和两个谷期(分别为11:30—12:30、14:30—15:30);飞行行为存在4个峰期(分别为9:30—10:30、11:30—12:30、14:30—15:30、16:30—17:30)和3个谷期(分别为10:30—11:30、13:30—14:30、15:30—16:30);修整行为在13:30—14:30有不明显峰期;其他行为均没有明显波动。

|
| 图 3 青头潜鸭越冬行为活动节律 Fig. 3 The wintering behavior rhythms of Baer′s Pochard 图中的09:30代表 09:30—10:30, 依此类推 |
不同性别青头潜鸭越冬行为时间分配(表 2)分析表明:(1)雄性青头潜鸭的7类越冬行为中, 静息、觅食和运动三类行为是主要行为(超过70%);其次是修整和飞行两类行为(超过25%);社会和特殊两类行为时间分配最少。这与青头潜鸭整体层面越冬行为时间分配相一致。(2)雌性青头潜鸭的7类越冬行为中, 用于运动、飞行和觅食三类行为的时间比例较大(接近70%);其次是静息和修整两类行为(超过25%);社会和特殊两类行为时间分配最少。
| 行为类别 Behavior type | 雄性Male M±SE/% | 雌性Female M±SE/% | 卡方值 c2-value | P |
| 觅食 Foraging | 21.53±1.66 | 21.78±2.03 | 1.996 | 0.158 |
| 静息 Resting | 31.13±4.01 | 16.56±5.28 | 70.340 | 0.000 |
| 修整 Maintain | 14.70±1.03 | 10.44±0.98 | 126.286 | 0.000 |
| 运动 Locomotion | 19.82±1.69 | 25.97±2.64 | 87.819 | 0.000 |
| 飞行 Flying | 10.31±2.39 | 22.06±4.58 | 268.626 | 0.000 |
| 社会 Social | 2.43±0.49 | 3.15±0.83 | 3.154 | 0.076 |
| 特殊 Special | 0.09±0.03 | 00.03±0.03 | 1.271 | 0.260 |
卡方检验和显著分析结果(表 2和图 4、图 5)表明:民权湿地公园, 不同性别青头潜鸭越冬行为时间分配存在差异, 二者在静息、修整、运动和飞行四类行为时间分配方面存在显著性差异, 二者用于觅食(c2=1.996, P=0.158)、社会(c2=3.154, P=0.076)和特殊(c2=1.271, P=0.260)行为的时间无显著性差异。

|
| 图 4 不同性别青头潜鸭越冬行为时间分配比较 Fig. 4 Wintering behavior time allocation between female and male for Baer′s Pochard |

|
| 图 5 不同性别青头潜鸭越冬期行为节律 Fig. 5 The wintering behavior rhythms of Baer′s Pochard between female and male |
其中, (1)静息行为时间分配, 雄性极显著高于雌性(c2=70.340, P=0.000);活动节律也明显不同, 雄性全时段高时间分配, 14:30之后出现昼间峰期, 持续到17:30;而雌性除了12:30—13:30出现小峰期外, 16:30—17:30达到昼间大峰期, 接近雄性。这意味着雄性个体进入静息时段较早, 雌性个体较晚。
(2) 修整行为时间分配, 雄性极显著高于雌性(c2=126.286, P=0.000);二者活动节律极为相似, 呈“S”型曲线波动, 其中二者的活动节律在13:30—14:30达到昼间最高峰期, 雄性峰期持续到16:30。
(3) 运动行为时间分配, 雄性极显著低于雌性(c2=87.819, P=0.000);雌性呈“M”型的全时段高时间分配, 峰期为10:30—11:30和15:30—16:30、谷期为12:30—13:30, 而雄性恰恰相反, 呈全时段的低时间分配且峰期不明显, 仅16:30—17:30出现明显的峰期, 这种时间分配格局和活动节律与静息行为恰恰相反。
(4) 飞行行为时间分配, 雄性极显著低于雌性(c2=268.626, P=0.000);二者的活动节律完全一致, 均呈“WV”型曲线波动, 均存在四个峰期(分别为9:30—10:30、11:30—12:30、14:30—15:30和16:30—17:30)和3个谷期(分别为10:30—11:30、13:30—14:30和15:30—16:30)。
(5) 觅食行为时间分配, 二者不存在显著差异, 但雌性个体整体略高于雄性个体, 活动节律也相似, 整体上呈“W”型曲线波动, 但雄性个体在早晚的时间分配偏低, 在9:30—10:30和16:30—17:30两个时段不如雌性频繁;其中, 雄性存在3个峰期(10:30—11:30、13:30—14:30、15:30—16:30)和两个谷期(11:30—12:30、14:30—15:30), 雌性存在3个峰期(09:30—10:30、13:30—14:30、16:30—17:30)和两个谷期(11:30—12:30、14:30—15:30)。
(6) 社会行为时间分配, 二者不存在显著差异, 但11:30—16:30时段, 雌性个体显著高于雄性个体, 而雄性个体仅在15:30—16:30期间出现昼间高峰期。
(7) 特殊行为时间分配, 二者不存在显著差异, 在15:30—17:30期间雄性个体出现昼间高峰期, 雌性个体的昼间峰期为12:30—13:30, 这与雄性个体部分同步。
4 讨论 4.1 越冬行为模式 4.1.1 越冬行为时间分配民权湿地公园的青头潜鸭越冬期主要行为模式与白眼潜鸭(Aythya nyroca)、中华秋沙鸭(Mergus squamatus)、白头硬尾鸭(Oxyura leucocephala)、红头潜鸭、大天鹅、小天鹅、白额雁(Anser albifrons)、豆雁等雁鸭类存有异同。其中, 越冬期日间行为模式基本一致[27-34], 均以觅食和静息为主。觅食是能量补充, 静息是节约能量消耗, 两类行为时间分配约为50%, 这是鸟类维持日常生命活动的基础, 符合行为投资与收益的能量学理论[35-36]。
但静息和觅食行为的时间分配在不同鸭种间存在较大差异, 本文研究中青头潜鸭用于觅食的行为时间分配(21.66%)远低于白眼潜鸭(56.7%)、中华秋沙鸭(40.42%—44.22%)、白头硬尾鸭(48.90%)、红头潜鸭(34%)、小天鹅(30.52%)、白额雁(81.3%)、豆雁(63.8%)等雁鸭类[27-34], 而用于静息的行为时间分配(28.15%)高于或远高于白眼潜鸭(11.2%)、中华秋沙鸭(9.06%—10.08%)、白头硬尾鸭(23.32%)、豆雁(15.6%)等雁鸭类[27-29, 34], 却远低于红头潜鸭(39%)、大天鹅(40.5%)、小天鹅(45.93%)、白额雁(44.9%)等雁鸭类[30-33]。本研究中, 运动行为也是该地区青头潜鸭越冬期的主要行为(21.09%)。本研究的越冬行为谱中, 运动行为(21.09%)主要以游泳为主, 这与白眼潜鸭(13.8%)、中华秋沙鸭(22.72%—24.77%)、白头硬尾鸭(9.48%)、红头潜鸭(10%)、大天鹅(22.8%)、小天鹅(9.49%)等雁鸭类[27-31]存有较大差异。形成这种差异的解释有两种。一种是该区域青头潜鸭维持越冬所需要的能量代谢率与其他雁鸭类存有差异。这方面需要通过能量摄入量、维持正常生理机能的基础代谢率、能量排泄量、运动消耗量等多因素进行综合测量[37-38]。另一种是该区域青头潜鸭观察期间的食物供给充足、食物可获得性高, 其有效觅食时间分配较少。这种解释需要进一步的量化研究。
行为相关性分析中, 部分行为类别呈正相关、部分行为类别呈负相关。其中, 运动行为与觅食、修整、社会等行为之间呈极显著正相关, 修整行为与社会行为之间呈极显著正相关。本文的运动行为包括游泳和嬉戏, 觅食行为包括取食、喝水、潜食, 修整行为包括理羽、挠头、抖翅、洗澡、伸展, 社会行为包括种内驱赶和种间驱赶。研究区内的藕塘生境是青头潜鸭偏爱利用的主要活动区, 该区域藕杆等植物密度极高, 在为青头潜鸭提供隐蔽功能的同时, 也为其觅食活动提供了障碍, 难以固定在某一固定区域觅食, 需要往返式运动, 觅食行为时间分配越高, 其运动行为时间分配也必然会越高, 二者之间呈正相关。同时, 在运动过程中, 必然会出现种内个体、种间个体在食物利用等方面的追赶、抢食、打斗等驱赶性社会行为。在实际观察中发现, 青头潜鸭在进行觅食行为的过程中, 如叼着食物或刚结束潜食时, 常被白骨顶(Fulica atra)、黑水鸡(Gallinula chloropus)、罗纹鸭(Anas falcata)等其他水鸟追赶, 但未出现其他水鸟抢食成功的现象, 推测是青头潜鸭较白骨顶、黑水鸡、罗纹鸭的潜水能力更强, 在深水中取食效率更高, 其他种间水鸟的追赶增加了青头潜鸭运动行为的频率, 运动过程中必然会出现羽枝、羽小枝的错位与环境物质在羽毛中的夹带, 运动后的羽枝与羽小枝需要重新梳理与正位, 理羽、挠头等修整行为是较好的梳理方式[14, 39]。
静息行为与修整、运动、社会等行为之间呈极显著负相关, 飞行行为与觅食、静息、修整等行为之间呈极显著负相关。本文的静息行为是广义概念, 包括睡眠和狭义的静息。静息行为是青头潜鸭昼间活动时间分配最高的行为, 在昼间活动时长固定的前提下, 游泳等运动行为和驱赶性社会行为的时间分配必然会减少, 修整行为一般发生在静息结束后或驱赶行为发生后等特定时段, 静息时间分配越多, 其修整的机会便会越少, 这种极显著的负相关便会越明显。此外, 青头潜鸭在觅食、静息、修整等行为方面分配的时间越多, 其飞行行为时间分配必然会减少, 本研究中的飞行行为时间分配较低, 该行为在冬季除了受到惊扰逃跑和改变活动区域以外, 可用于巡视警戒周围情况, 属于高耗能行为[40-41];对青头潜鸭来说, 在冬季减少能耗、积累能量抵御寒冬更具有生态意义, 呈极显著负相关的低时间分配也符合鸟类越冬生存策略。
4.1.2 越冬行为活动节律本研究中, 青头潜鸭静息行为在昼间全时段均有较高的时间分配, 时间分配波动较缓, 节律性不明显。研究表明, 静息是许多野生动物越冬期间应对低温节省能耗的一种生存策略(如鸳鸯、小天鹅和大天鹅等)[31-32, 42], 青头潜鸭的这种全时段高时间分配的静息方式符合野生动物越冬期减少能耗及有效保存能量的生存对策。
在行为时间节律方面, 觅食行为(峰期为10:30—11:30、13:30—14:30、15:30—17:30, 谷期为11:30—12:30、14:30—15:30)和飞行行为(峰期为9:30—10:30、11:30—12:30、14:30—15:30、16:30—17:30, 谷期为10:30—11:30、13:30—14:30、15:30—16:30)具有明显的节律性。可以看出, 研究区域内青头潜鸭越冬期昼间觅食有早、午、晚3个高峰, 在午前、午后有两个觅食低谷, 觅食低谷是飞行的高峰期, 但青头潜鸭的飞行高峰不仅限于午前与午后, 早、晚还各有一次飞行高峰。
大部分雁鸭类的觅食行为会在早晚出现高峰期, 这种行为有利于雁鸭类更好地应对夜间的能量消耗, 如白头硬尾鸭的取食高峰期为7:00—9:00和16:00—19:00、大天鹅的取食高峰期为7:00—8:00和17:00—18:00、斑头雁的取食高峰期为9:00—9:59和17:00以后[29, 31, 43]。本研究中青头潜鸭越冬期觅食行为节律与上述研究结果在模式上类似, 但具体的峰期存有差异。究其原因, 可以从与觅食有关的外因和内因角度进行考虑。外因中的天气因素是重点, 观察期间, 观察区域的雾气较大, 雾气受太阳辐射的冲击作用[44], 每天8:30—9:30才散去;同时, 观察期间, 10:30—11:30、13:30—14:30是温度最佳的时段, 选取温度最佳的、能耗最低的、可见率最好的时段觅食是鸟类针对越冬期温度的一种策略, 但15:30—17:30这个觅食峰期不符合该理论, 这方面需要进一步的研究分析。内因中的食物可获得性是重点, 青头潜鸭的食谱包括哪些、食物可获得性与温度是否有关等等, 这些内容目前还是未知, 需要进一步的研究。青头潜鸭觅食行为和飞行行为时间分配的峰期与谷期恰恰相反, 这与二者行为相关性呈极显著的负相关相符合, 节律性也特明显。
4.2 越冬行为模式的性别差异不同性别青头潜鸭越冬行为模式比较分析的结果显示出:二者在主要越冬行为类别、越冬行为时间分配、越冬行为时间节律等方面均存在一定程度的性别差异。
其中, 主要越冬行为类别方面, 雄性青头潜鸭以静息、觅食和运动行为为主, 雌性青头潜鸭以运动、飞行和觅食行为为主。觅食行为是雌雄青头潜鸭两性均有的主要越冬行为类别, 这也致使二者的觅食行为时间分配不具有显著性差异。
越冬行为时间分配方面, 雄性个体用于静息和修整行为的时间分配明显高于雌性个体, 而用于运动和飞行行为的时间分配显著低于雌性个体;但二者在静息、修整、运动、飞行、觅食、社会、特殊等行为节律方面均存在程度不一的性别差异。其中, 觅食行为模式方面, 雌雄个体之间在时间分配格局与活动节律基本一致但早晚频率偏低, 这与曾宾宾等(2013)的研究结果相反[28]。经过夜晚的能耗, 需要早上及时补充能量, 但为什么只有雌性出现了高频率的觅食行为, 而雄性没有, 也许还与雌性个体在繁殖期所处的角色地位有关。静息行为时间节律与修整行为时间节律存在极大的重叠, 两种行为中雄性个体均呈现出全时段时间分配显著高于雌性、且进入静息较早、下午修整峰期持续较长的现象, 在符合鸟类冬季减少能耗保存能量的策略外, 也许还存在性别间的社会地位、生活史过程中的角色差异影响, 也许是繁殖期中扮演的角色差异形成的一种行为习惯[31-32, 42]。雌性的修整行为在13:30—14:30时有一个小高峰, 而雄性在该时段也呈增多趋势, 青头潜鸭中午集中修整这一行为对策与越冬黑颈鹤相似[45]。雄性青头潜鸭较雌性个体外貌鲜艳漂亮, 这是繁殖期间赖以求偶、选择配偶的根本, 进行日常的修饰行为也许是一种进化需要。
为什么雄性个体昼间需要更多的静息时间分配, 是夜间静息时间不够还是一种越冬期间保持体能的行为常态?雌性个体更高的运动、飞行时间分配, 为什么会出现这种行为差异, 其后的行为学解释是什么, 对青头潜鸭越冬生物学有着什么样的重要功能?青头潜鸭不同性别间越冬行为模式差异的驱动因素是什么?也许是冬季所有的性别差异需要通过其他季节的行为模式来验证;也许是观察时间影响所致, 本文仅是越冬期的一部分, 并非整个越冬期;也许是不同天气所致。所有这些方面的研究都需要进一步的研究关注。
| [1] |
邵明勤, 龚浩林, 戴年华, 植毅进, 徐宁, 卢萍. 鄱阳湖围垦区藕塘越冬白鹤的时间分配与行为节律. 生态学报, 2018, 38(14): 5206-5212. |
| [2] |
Belovsky G E, Slade J B. Time budgets of grassland herbivores:body size similarities. Oecologia, 1986, 70(1): 53-62. DOI:10.1007/BF00377110 |
| [3] |
张国钢, 孙戈, 孙静, 陆军. 西藏阿里班公错斑头雁的种间巢寄生行为. 动物学杂志, 2017, 52(4): 664-667. |
| [4] |
曹铭昌, 刘高焕, 徐海根. 丹顶鹤多尺度生境选择机制——以黄河三角洲自然保护区为例. 生态学报, 2011, 31(21): 6344-6352. |
| [5] |
刘学昌, 吴庆明, 邹红菲, 李晓民. 丹顶鹤(Grus japonensis)东、西种群巢址选择的分异. 生态学报, 2009, 29(8): 4483-4491. |
| [6] |
车烨, 杨乐, 李忠秋. 西藏拉萨越冬黑颈鹤家庭群的警戒同步性. 生态学报, 2018, 38(4): 1375-1381. |
| [7] |
蒋剑虹, 戴年华, 邵明勤, 黄志强, 卢萍. 鄱阳湖区稻田生境中灰鹤越冬行为的时间分配与觅食行为. 生态学报, 2015, 35(2): 270-279. |
| [8] |
Hearn R D, Tao X, Hilton G. A species in serious trouble:Baer's pochard Aythya baeri is heading for extinction in the wild. Birding Asia, 2013, 19: 63-67. |
| [9] |
Heim W, Wolanksa K, Siegmund A, Schuster U. Possible breeding of Baer's pochard Aythya baeri at Muraviovka Park, Far East Russia. Birding Asia, 2013, 20: 64-66. |
| [10] |
许鹏, 陈潘, 汪国海, 刘彬, 张曼玉, 鲁长虎. 极危物种青头潜鸭在安徽滁州市池杉湖湿地公园繁殖记录. 动物学杂志, 2019, 54(3): 451-452. |
| [11] |
张琦, 朱井丽, 吴可新, 李小琴, 李连山, 吴庆明. 青头潜鸭在向海自然保护区的分布状况. 野生动物学报, 2019, 40(2): 510-512. |
| [12] |
常家传, 桂千惠子, 刘伯文, 张鹏. 东北鸟类图鉴. 哈尔滨:黑龙江科学技术出版社, 1995, 42. |
| [13] |
李桂恒. 四川鸟类原色图鉴. 北京: 中国林业出版社, 1993: 44-44.
|
| [14] |
郑作新. 中国鸟类区系纲要. 北京: 科学出版社, 1987: 65-65.
|
| [15] |
郑作新. 中国鸟类分布名录(第二版). 北京: 科学出版社, 1976: 60-60.
|
| [16] |
王雪峰, 颜军, 魏斌, 谭军. 武汉市黄陂区青头潜鸭繁殖生态观察初报. 野生动物学报, 2019, 40(1): 190-195. |
| [17] |
胡扬, 黄文杰. 九江市柴桑区东湖青头潜鸭种群监测现状与栖息地保护对策. 现代农业科技, 2018(20): 221-221, 227-227. |
| [18] |
颜军, 王雪峰, 谭军, 魏斌, 陈韬, 明亮, 陆峥, 赵学迅, 冯江. 武汉市黄陂区青头潜鸭的分布初步研究. 湖北林业科技, 2018, 47(5): 34-36. |
| [19] |
汪青雄, 肖红. 内蒙古呼和浩特市发现青头潜鸭. 四川动物, 2017, 36(6): 673-673. |
| [20] |
郭玉民, 林剑声, 沈俊峰, 王榄华, 章麟, 何芬奇. 关于青头潜鸭九江群体的最新报道. 野生动物学报, 2017, 38(3): 535-538. |
| [21] |
杨晓菁, 张菁, 汪海兵, 张运晨, 马龙峰. 湖北武汉涨渡湖发现全球极度濒危物种青头潜鸭群. 动物学杂志, 2017, 52(3): 430-430. |
| [22] |
吴渊, 于海龙, 刘亚东, 孙万兵, 洪剑明. 张家口坝上康巴诺尔湖世界极度濒危物种青头潜鸭的新发现. 湿地科学与管理, 2017, 13(1): 65-65. |
| [23] |
郭玉民, 闻丞, 林剑声, 何芬奇. 青头潜鸭(Aythya baeri)在中国的近期分布. 野生动物学报, 2016, 37(4): 382-385. |
| [24] |
杨军, 阳丽萍. 世界极危鸟类青头潜鸭现身沅江漉湖. 林业与生态, 2016(4): 46-46. |
| [25] |
高继宏, 孙相吾, 陶宇, 曾昭文. 青头潜鸭繁殖研究初报. 野生动物, 1992, 13(2): 25-27. |
| [26] |
张毅力, 宋芳鑫.民权黄河故道国家湿地公园成为"青头潜鸭重要繁殖地". (2019-09-14). http://henan.people.com.cn/n2/2019/0914/c351638-33353564.html
|
| [27] |
赵序茅, 马鸣, 张同. 白眼潜鸭秋季行为时间分配及活动节律. 动物学杂志, 2013, 48(6): 942-946. |
| [28] |
曾宾宾, 邵明勤, 赖宏清, 蒋剑虹, 李畅, 戴年华. 性别和温度对中华秋沙鸭越冬行为的影响. 生态学报, 2013, 33(12): 3712-3721. |
| [29] |
赵序茅, 马鸣, 张同, 张建波. 白头硬尾鸭行为时间分配及日活动节律. 生态学杂志, 2013, 32(9): 2439-2443. |
| [30] |
查林松, 宋洋, 王睿姝, 张雪丹, 许青. 齐齐哈尔机场红头潜鸭秋季昼间行为时间分配节律与鸟击防治. 野生动物, 2013, 34(4): 198-201. |
| [31] |
董超, 张国钢, 陆军, 李新平, 马俏飞, 侯韵秋, 关丙役. 山西平陆越冬大天鹅日间行为模式. 生态学报, 2015, 35(2): 290-296. |
| [32] |
邵明勤, 张聪敏, 戴年华, 徐宁, 植毅进, 卢萍. 越冬小天鹅在鄱阳湖围垦区藕塘生境的时间分配与行为节律. 生态学杂志, 2018, 37(3): 817-822. |
| [33] |
杨秀丽.安徽升金湖国家级自然保护区白额雁(Anser albifrons)数量分布, 觅食行为和食性变化研究[D].合肥: 中国科学技术大学, 2011. http://d.wanfangdata.com.cn/Thesis_Y2029947.aspx
|
| [34] |
刘静.安徽升金湖国家级自然保护区豆雁的越冬食性和行为研究[D].合肥: 中国科学技术大学, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10358-1012008991.htm
|
| [35] |
杨志宏, 邹红菲, 高忠燕, 郭立业, 徐慧. 食物可获得性对冬季丹顶鹤能量收支的影响——以扎龙自然保护区人工辅助繁育丹顶鹤为例. 生态学报, 2015, 35(13): 4408-4415. |
| [36] |
Green A, Hughes B. Oxyura leucocephala White-headed duck. BWP Update, 2001, 3(2): 79-90. |
| [37] |
倪小英, 林琳, 周菲菲, 王小华, 柳劲松. 光周期对白头鹎体重、器官重量和能量代谢的影响. 生态学报, 2011, 31(6): 1703-1713. |
| [38] |
周围, 王玉萍, 陈德汉, 柳劲松. 白头鹎体温、体重及能量代谢的昼夜节律. 生态学杂志, 2010, 29(12): 2395-2400. |
| [39] |
黎红辉, 沈猷慧, 马再玉. 鸟类飞羽羽小枝的显微结构比较. 动物分类学报, 2005, 30(4): 666-675. |
| [40] |
谢世林, 曹垒, 逯非, 欧阳志云. 鸟类对城市化的适应. 生态学报, 2016, 36(21): 6696-6707. |
| [41] |
邵明勤, 陈斌. 中华秋沙鸭越冬期昼间行为能量支出及其变化. 湿地科学, 2017, 15(4): 483-488. |
| [42] |
阮云秋. 鸳鸯越冬期日活动行为时间分配的研究. 野生动物, 1995, 16(6): 19-23. |
| [43] |
杨延峰, 张国钢, 陆军, 刘文, 李振吉. 贵州草海越冬斑头雁日间行为模式及环境因素对行为的影响. 生态学报, 2012, 32(23): 7280-7288. |
| [44] |
陈跃浩, 熊明明, 曹经福, 杨艳娟. 雾霾天气对天津市太阳辐射影响的量化研究. 气象与环境学报, 2018, 34(5): 25-30. |
| [45] |
孔德军, 杨晓君, 钟兴耀, 道美标, 朱勇. 云南大山包黑颈鹤日间越冬时间分配和活动节律. 动物学研究, 2008, 29(2): 195-202. |