 2020, Vol. 40
2020, Vol. 40文章信息
- 宋明华, 朱珏妃, 牛书丽
- SONG Minghua, ZHU Juefei, NIU Shuli
- 生态系统状态跃变的早期预警信号及检测
- Testing on the early warning signals in approach to ecosystem state transition
- 生态学报. 2020, 40(18): 6282-6292
- Acta Ecologica Sinica. 2020, 40(18): 6282-6292
- http://dx.doi.org/10.5846/stxb201909272032
-
文章历史
- 收稿日期: 2019-09-27
- 修订日期: 2020-08-09





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
生态系统自身具有高度的复杂性, 如不稳定性、高维度性、非线性及随机性等特征[1-2]。当他们处于正、负反馈环的作用下, 会在某种程度上对环境变化表现出时滞现象和稳定状态[1]。生态系统固有的复杂性给人类的认识带来了很大的挑战。然而, 人类对全球生态系统的影响持续增加, 对生态系统结构、功能以及其提供的食物、水等服务产生深刻影响[3], 这些加剧了生态系统复杂性的程度。另外, 模型模拟与预测必须建立在对生态系统动态过程的理解和认识的基础上, 同时需要在大量详尽数据基础上对生态系统机理、过程等进行参数化[2]。因此对生态系统结构和功能的模拟与预测就越加紧迫而艰巨。正是如此多的挑战, 吸引着科学家们不断探索和完善系统跃变理论、解析突变系统的案例, 推动着该领域的发展。早期预警信号的研究通过捕捉生态系统接近临界拐点前的结构和属性上的变化特征, 结合模型模拟以及统计学参数的变化特征, 逐渐完善了早期预警信号的理论框架。利用临界点恢复力的速率减慢、系统状态趋于分叉时方差增加, 以及系统自相关增强等信号, 检测了自然生态系统状态跃变前的行为。随着对系统格局和组分相互作用在状态跃变过程中的响应方式的不断认识, 揭示了生态系统组分在状态跃变过程中的相互作用机制, 认为在环境压力作用下, 系统的自组织, 即资源与消费者之间的正反馈作用是联结系统空间结构特征与状态跃变的重要机制。随着在方法和机理等方面研究的不断深入, 生态系统早期预警信号研究逐渐过渡到利用大尺度、长时间序列的数据集、结合多度和性状等变化特征, 建立多信号的预警体系。生态系统状态跃变关系到生态系统服务功能的提供、生态安全以及社会经济利益损失等, 另外对状态突变后的生态系统进行修复也需耗费巨大的代价。忽略了生态系统状态跃变可能会带来严重的社会经济损失。因此, 深入认识和理解生态系统随环境和外界干扰的渐变与突变转化机理, 认清生态系统演变过程, 探寻其状态转变的特征, 进而捕捉状态转变的预警信号, 对于生态系统的科学经营和管理、界定适度的干扰强度、并对干扰后的生态系统进行及时修复、避免生态系统崩溃带给人类的灾难性后果具有重要的理论和现实意义。
1 生态系统状态跃变的早期预警理论生态系统经历着气候变化、人类活动干扰、养分输入输出等变化的影响。通常生态系统会对气候的波动、营养物质输入输出、生境碎片化、生物多样性增加或减少等等的变化以一种自然平稳或缓慢渐变的方式做出响应, 此时生态系统状态随着环境的变化呈现一种线性且持续的变化, 这些变化往往表现出惰性或缓慢渐变特征。然而, 当环境条件变化的强度或频率接近某一临界水平时, 生态系统会做出强烈响应, 其状态会发生突变。即使环境条件恢复到突变前的水平, 生态系统不会随外界条件的恢复而回到原来的状态, 系统已经处于另一种稳定状态[1]。生态系统在多稳态之间发生状态跃变的点或者区域被称为生态系统阈值。生态系统的阈值标识着生态系统结构、功能、属性的突变, 超过阈值范围, 生态系统会发生不可逆转的状态转变。早期预警从相关概念的提出到理论体系的形成, 其中融合了多学科的知识。如复杂系统理论, 多稳态理论, 动态系统理论, 物理学的相变理论, 分形理论, 自组织机制, 系统动力学理论等等, 这些理论均融入早期预警理论的发展历程。
Holling[4]在1973年论述了生态系统具有一定的弹性和稳定性, 明晰了生态系统稳定性、弹性和恢复力的概念、理论和计算方法。后期的大量有关生态系统稳定性的研究多基于状态之间的对比, 以及特定状态下生态系统稳定性维持机制的探讨、稳定性衡量的多维度性等等[5-8]。随着对生态系统状态转变的线性与非线性过程及其机制的深入研究, 提出了生态系统多稳态、状态转变阈值、临界减速、早期预警信号等概念来刻画生态系统状态转变的特征, 并提出了分叉、折叠分叉和突变性分叉等概念(表 1), 这些为逐步完善生态系统跃变的理论奠定了基础[1-2]。生态学家结合复杂系统理论、动态系统理论、系统动力学理论和物理学的相变理论等, 将其应用于生态系统状态跃变过程研究中, 进而提出系统的一阶和二阶相变。一阶相变是非线性变化过程, 对于外力的响应是不连续的, 较小的外界力量就会导致系统状态发生迅速变化。如生态系统会从由正负反馈环维持的一种状态通过分叉或临界点移动到另一个由反馈环维持的稳定状态, 这种非线性的转变通常称为状态突变、相变或临界转变[1]。二阶相变对外力的响应通常是连续线性的, 外界环境变化足够大时, 才能使生态系统发生较大的变化[1]。跨临界分叉是系统从一种状态连续地变化到另一种状态, 而没有以折叠分叉为特征的快速移动, 被认为是一种二阶相变。
| 名词术语Terminology | 含义Definition |
| 多稳态 Alternative stable state |
生态系统的替代状态, 由于生态系统功能和组成变化导致的生态系统特征改变, 通常由负反馈环维持 |
| 分岔 Bifurcation |
数学术语, 表示系统从一个稳定状态, 即“吸引域(basin of attraction)”, 通过不稳定区域转移到另一个“吸引域” |
| 临界减速现象 Critical slowing down |
在临界点附近, 系统的稳定性下降, 导致系统恢复到以前状态的速度变慢的现象 |
| 早期预警信号 Early warning signal |
表示系统接近临界过渡状态时的一些特征, 也可以是一种统计信号 |
| 折叠分岔 Fold, catastrophic or saddle-node bifurcation |
系统从一种稳定状态快速转换到另一稳定状态, 并有滞后现象, 其特征是系统变率的主要特征值为零。是一阶相变的例子 |
| 强迫Forcing | 使系统不稳定, 将其推向临界点的外部压力 |
| Hopf分岔Hopf bifurcation | 系统从稳定的平衡态过渡到稳定的循环运动态 |
| 滞后 Hysteresis |
在相同环境条件下存在多种稳态-由正反馈环驱动-意味着系统的当前状态也取决于该系统的历史状态。滞后意味着恢复到以前的稳态具有重要意义, 尤其是表现出折叠分岔的系统 |
| 先行指标 Leading indicator |
预测在临界点附近发生显著变化的一组统计数据之一, 常与早期预警信号互换使用。然而, 我们建议早期预警信号将更好地用于描述检测到的并能够指示系统接近崩溃的信号, 而先行指标与不同统计量密切相关, 可用于检测早期预警信号, 如方差或自相关等指标 |
| 非灾难性转变 Non-catastrophic transition |
系统不经过折叠分岔, 而是平稳地移动到新的状态, 以响应强迫。这可能以跨临界分岔的形式或者在强的非线性变化的情况下发生 |
| 格局Pattern | 一般用于自上而下地描述二维景观中植被的空间布局 |
| 一阶相变 Phase transition, first order |
典型的例子是折叠或灾难性分岔, 一阶相变特征是系统对外力的非连续性响应 |
| 二阶相变 Phase transition, second order |
典型的例子是跨临界分岔, 二阶相变特征是系统对外力的连续性响应 |
| 临界转变 Regime shift (critical transition) |
由系统外部强迫引起的生态系统从一个稳定状态转变为另一个稳定状态。多稳态的存在也意味着系统表现出滞后现象, 因此将系统恢复到以前的状态可能是非常重要的。重要的是, 并非所有的临界转变都与临界减速有关 |
| 弹性 Resilience |
通常指与系统所在“吸引域”的深度和形状相关的系统的整体特性, 即系统在维持功能的同时可以吸收的干扰量。当受到压力时, 系统的弹性会发生变化, 因为系统的外力会改变“吸引域”的形状, 使系统对进一步扰动的弹性发生变化 |
| 稳定性 Stability |
系统在扰动后恢复到先前状态的速率。系统受力, “吸引域”形状发生变化, 导致稳定性丧失。这种稳定性损失由基于临界减速现象的早期预警信号测量, 并用作系统弹性损失的代理度量 |
| 临界点 Tipping point |
系统不稳定的点, 即使较弱的扰动也会引起系统状态的剧烈变化。在数学上, 当主特征值达到零时, 就会出现这种情况, 较为典型的现象是在折叠分岔中观察到的快速转变 |
| 跨临界分岔 Transcritical bifurcation |
系统从一种状态连续地变化到另一种状态, 而没有以折叠分岔为特征的快速移动, 但主要特征值也通过零。一个例子是种群的灭绝, 系统从种群现存的一个稳定状态转移到种群灭绝的另一稳定状态。这是一个二阶相变的例子 |
| Ⅰ类错误 Type Ⅰ error |
假阳性信号, 即早期预警信号错误地指示出系统将要崩溃 |
| Ⅱ类错误 Type Ⅱ error |
假阴性信号, 即系统崩溃前未能检测到早期预警信号 |
系统状态跃变的理论构架可以更加直观地用经典的“球-杯”图来解释(图 1)[1-2, 9]。一个系统的可能状态取决于其在多个稳态之间转换时需要的能量。系统的恢复力是指系统状态所处“吸引域”的形状和大小。稳定性是指干扰后系统返回到原初状态的速率。当外界压力打破系统的负反馈环时, “吸引域”收缩, 仅需少量能量就会使系统脱离目前的状态, 系统的恢复力消失, 稳定性状态改变。不同的生态系统阈值各不相同, 但临界点附近的生态系统的动态具有某些共有的特征, 这些特征变化便是生态系统临近其状态突变的阈值的预警信号。生态系统临近其状态跃变发生时, 会表现出临界减速的现象, 即在接近临界点时, 生态系统从扰动中恢复的速率变得越来越慢。另外, 系统接近状态跃变时会发生波动模式的某些特征变化, 一个重要的变化可能是在临界减速所产生的波动模式中, 反映生态系统属性的敏感变量的自相关和方差增加。这三者可以作为系统接近状态突变前的早期预警信号。除了自相关和方差增加外, 在跃变的临界分岔点附近还可以看到闪烁现象, 即系统在两个状态之间来回移动, 如果干扰持续进行, 系统可能会永久的移动到另一个稳定的状态[1]。因此, 理论上统计学的期望是系统处于临近分叉点时会表现出恢复力的速率减慢, 自相关和方差增加, 密度比降低[10]。这一基于“临界减速”的早期预警信号对系统动态的过程不要求有过多假设[1], 因此成功地对很多系统, 如微生物群落, 淡水湖泊, 社会生态系统, 气候系统, 草地生态系统, 以及疾病等进行了模型模拟[11-17]。

|
| 图 1 经典“球-杯”模型解释多稳态、稳定性和恢复力理论 Fig. 1 The classic ball-cup explanation of alterative stable state, stability and resilience theory |
这一理论框架中系统的状态通过蓝色线上的球的位置来指示。(1)和(2)表明了“吸引域”的大小, 较大的和较深的域表明系统的恢复力较高, 较浅的和较窄的域表明系统的恢复力较低。绿色部分指示系统脱离目前的“吸引域”经过阈值点(5和6)到达另一稳定状态(3和4)所需的潜在能量, 黑色虚线指示着路径。要实现这一转变, 具有强恢复力的系统相对具有较弱恢复力的系统需要较多的能量(如:较大的干扰)。系统的恢复速率通过“吸引域”的坡度来指示, 斜坡越陡表明系统能够很快恢复到原来的平衡态(滑回到7状态), 然而, 对于稳定性低的系统, 较浅的坡度意味着恢复原初状态需要较长时间(8状态)。一个系统可能起始于高的恢复力, 但是外部力会改变稳定性景观的形态(蓝线), 因此改变“吸引域”, 某种程度上恢复力和稳定性会丧失(系统从左手侧转变为右手侧状态)。实际上, 系统稳定性的降低可以作为恢复力降低的可测量的待用指标, 形式上表现为系统动态的临界减速(如:种群时间序列上方差增加)[2])。
过去20多年里, 生态系统早期预警信号研究越来越多样化, 从单一统计学上的简单变化趋势转变为基于似然率检测[10, 18-19]和整合多个同步数据集的更加复杂的途径[20, 13]。预测生态学的主要目标是在全球环境变化背景下检测物种、群落和生态系统在气候变化和人类活动压力驱动下的状态跃变的危机[21]。面对全球生物多样性丢失危机, 通过检测拐点、状态改变和种群崩溃等系统共有的早期预警信号, 可望为现实世界中的问题提供更多的工具, 为制定保护决策提供帮助。因此, 将多指标综合起来判定生态系统状态跃变的早期预警是一个更加可靠的途径[2]。
2 生态系统状态跃变的早期预警信号检测方法 2.1 基于统计学参数的早期预警信号早期预警信号是基于生态系统具有多稳态的条件下对生态系统突变的一种研究, 表示生态系统状态在转变的临界点或区域的一种统计学信号。由于其理论发展在前, 早期信号的检测多源于统计学参数的很强的时间变化趋势, 如方差在临界阈值点处的徒然增加, 属性在分叉点前自相关性增强, 恢复力急剧减慢等。然而这些统计学参数的变化趋势依赖于取样的频率, 状态转变模拟时采用的模型类型, 以及生态系统动态的类型, 因此其对系统状态转变指示的准确性很难界定。另外在参数计算过程中数据融合的方法, 数据是否进行去趋势处理, 数据处理时采用滑窗的大小等等的方法, 都会对参数变化的质量产生影响。另外, 从“噪音”中辨识统计变量如方差、自相关性等的变化趋势具有很大挑战性, 对数据样品量和取样频度要求很高[2]。
2.2 基于模型算法选择的信号由于早期预警信号的检测对数据取样与算法具有较强的依赖性, 因此具体采样方面考虑空间异质性采用跨样品方差, 条件异方差等扩大样本容量。另外在模型的算法方面经历了从简单的多度或空间结构数据的趋势测量到更复杂的基于最大似然模型的选择, 尽量考虑随机性因素。迄今为止, 绝大多数工作都集中在临界减速现象产生的信号上。尽管系统“临界减速”可以作为早期预警信号在分叉点前被检测到, 但是某些导致大的空间和长的时间尺度上的变化并不存在明显的分叉现象[22]。这些体现了分叉理论的明显缺点, 经典的早期预警理论的广泛应用被分叉理论的数学定义所妨碍了。因此, 在判别系统状态转移时, 是否表现出某一分叉类型不能作为判断早期预警信号下生态系统状态突变的一个必要条件[2]。
此外, 早期预警信号的检测可能会出现两类错误:Ⅰ类错误是假阳性信号, 即错误地确定存在预警信号;Ⅱ类错误是假阴性信号, 即系统崩溃前未检测到预警信号(表 1)。
2.3 基于多度的信号在最简单的生物模式下, 分叉可以通过以下确定性微分方程模拟, 这些方程描述了以对数增长模式生长的种群由于其个体受到过度收获影响而接近分叉[2]。
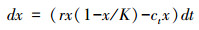
|
式中, dx是种群多度, r是种群增长率, K是环境承载力, c是种群的收获率;以r增长的种群多度受到以线性方式增长的收获率c的影响后, 当种群趋于灭绝过程中产生临界分叉现象。种群的这种变化是在均一的外部驱动力下导致的结果, 种群减少直至灭绝过程是趋于连续的和线性的。这种现象在自然界中并不普遍。在以上方程中, 如果系统对收获的响应做简单修改, 即引入半饱和常数以修饰种群大小降低到一定数量后对收获率的敏感程度, 于是产生了折叠或灾变分叉, 系统会从一个稳定的多度较高的种群经过种群个体数量的灾变性下跌转变为稳定的多度很低的种群系统[2]。
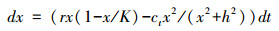
|
式中, h是半饱和常数。这些模型中收获率的增加有效地改变了“吸引域”的形状(图 1), 导致稳定性的丢失, 并且转变到了另一稳定状态(表现出折叠分叉处具有低的种群大小, 而在临界分叉状态下种群开始灭绝)。
2.4 基于性状的信号目前, 早期预警理论的一个主要发展方向是利用性状信息作为早期预警信号, 尤其是与适合度相关的表型性状, 环境对种群的影响通常是通过对种群个体形态大小或其他相关表型的调节体现的, 因此性状的改变通常会发生在整个种群的变化之前[2]。如物种个体大小、植物叶片和根系性状等, 这些都可以指示种群和生态系统的稳定性与健康状况[20, 23]。而且环境扰动驱动的种群动态和稳定性往往伴随着性状值分布的变化[24]。在选择适合的情况下, 表型性状的可塑性能够提供一个种群在当前和过去环境中的具体经历, 并进而推测其在未来环境扰动下种群的动态变化趋势[25]。种群尺度变化的影响会扩大到更大的尺度上, 食物网上一个物种体型的变化会使其他物种的取食习惯发生改变, 进一步会影响食物网的稳定性。因此, 性状信息除了在种群尺度上得到应用, 也可以通过尺度上推应用到更大的尺度上[23, 26]。通过将植物性状预警信号和通用的预警信号, 如自相关增加等, 联系起来可以提高对生态系统状态跃变预警的精准度。性状信息为早期预警研究提供了一个有着广阔前景的方法。尽管性状信息在早期预警信号的研究领域还存在许多不足和局限性, 但是可以通过很多方式产生有用的非临界减速的早期预警信号。总之, 开发替代方法或探寻潜在更加可靠的方法是未来早期预警信号理论发展的挑战, 同时还需明确大尺度上哪些数据可以获取[2](表 2)。
| 方法 Method |
数据 Data |
细节描述 Details |
文献 References |
| 统计信号的趋势 Trends in statistical signals |
多度或空间结构数据 | 强的时间变化趋势——具普遍意义的理论推导过程中采用的统计学变量作为警告信号:例如方差和自相关增加(通常使用滑动窗口方法进行估算)。在空间背景下, 采用空间方差, 空间偏斜度或空间相关性的变化趋势 | [10, 27-29] |
| 模型选择采用似然比进行检验 Model selection using likelihood ratio tests |
多度数据 | 选取代表退化和稳定状态的模型匹配获取的数据, 通过选择模型来确定哪个模型最能描述观察到的数据 | [30] |
| 跨样本方差 Across sample variance |
多样点多度数据 | 估算同一总体中多个采样点的调查样本之间的差异。能够很好甄别出明显的取样误差 | [31] |
| 条件异方差 Conditional heteroskedasticity |
多度 | 条件异方差意味着一个时间序列区域的方差与进行中的时间序列区域的方差高度相关。因此, 与接近分叉点的时间序列区域相比, 随着接近临界点, 时间序列在分叉点附近的部分将显示为高变异性群集 | [32] |
| 降低复杂性的测度 Measures of reduced complexity |
多度或空间 | 通过Kolmogorov算法复杂度的变化可以推断出系统随机性的变化, 降低的随机性(增加的复杂度)表明系统中的白噪声较少, 因此出现了分叉现象 | [33] |
| 光谱密度比 Spectral density ratio |
多度 | 光谱密度比测量一个时间序列中高频与低频过程的比率, 而向低频主导的过程转变(频谱“变红”)则表明系统即将崩溃 | [34] |
| 费希尔信息 Fisher information |
空间或多度 | 从表征系统状态的变量趋势中捕获系统动态中的模式。多个变量组合成一个信号, 该信号可以跟踪系统动态顺序的变化 | [35] |
| 快速设定变更点 Quickest change points |
多度 | 使用两种模型(崩溃与不崩溃, Boettiger C, Hastings A[18]), 这些模型随每次输入的新数据而更新。当似然比超过预定阈值时, 会根据用户对错误警报的容忍度生成信号 | [18-19] |
| 基于模式的空间信号 Pattern-based spatial signals |
空间 | 空间生态系统的组织结构和斑块变化可以指示生态系统状态转变, 尤其是斑块大小分布的变化, 基于“ Turing不稳定性”的自组织模式的出现以及观测到的偏离斑块大小的幂定律分布的迹象 | [29, 36-38] |
| 广义建模 Generalized modelling |
多度和结构信息 | 一般通用模型是在不指定特定功能形式的情况下描述系统结构的。这需要识别关键系统变量(例如多度)以及过程(例如出生率)或其他信息(例如死亡率可能是线性) | [39] |
| BDI模型的统计信号趋势 Trends in statistical signals of BDI models |
比率信息 | Dakos等[12]开发的基于趋势的信号扩展的模型, 由此可以通过寻找信号的变化趋势来预测突发疾病, 例如平稳的出生、死亡、移民过程的瞬间产生函数的方差系数等信号趋势 | [16] |
| 基于网络 Network-based |
多度或空间 | 基于网络中节点的连通性和集群系数, 较高的连通性表明即将发生的系统转变 | [15, 40] |
| 性状特征变化趋势 Trends in fitness-related traits |
性状 | 与适合度相关的性状特征的变化(无论是在种群水平还是在群落水平上的规模大小的下降)都被用来推断即将到来的生态系统崩溃 | [20, 23] |
| 综合信号 Combined signals |
多度和空间, 多度和性状 | 通过对每个指标的趋势进行归一化并在整个时间序列中求和, 将基于多度的稳定性度量(例如, 方差增加)以及基于空间或基于特征的度量合并为一个度量。因此, 可以产生一个可以减少Ⅰ型和Ⅱ型误差的复合指标 | [13, 20] |
生态系统的空间模式也可以作为预警信号, 当生态系统处于临近阈值前, 可能会出现特定的空间模式。比较典型的例子, 如植被在气候变化和人类活动干扰下逆向演替过程中, 往往在大的景观尺度上形成高度自相似的斑块化构型。近年来很多研究利用自然界收集的数据集检测到了空间上的预警信号。有研究发现, 空间异质性较高的酵母菌在临近菌落崩溃时表现出空间上恢复时间的迟滞[28]。利用咖啡树上寄生的害虫的分布结构, 能够很好地预测害虫对咖啡树木生产的影响[41]。另外从淡水水生生态系统到热带萨王纳[14-30]的空间结构变化均检测到状态转变的早期预警信号。如水生生态系统中, 海岸带上河蚌有规则的分布, 如条带状、聚集成堆等比随机分布被涨潮冲走的几率显著降低, 表明规则分布的河蚌对潮汐干扰的抗性更强[42-44]。
3.1 水生系统生态系统状态转变阈值的大小和外界干扰共同决定了生态系统的稳定状态。在对湖泊、珊瑚礁、海洋等生态系统的研究表明, 一些环境因子的变化使生态系统平缓的变化状态急剧切换到另一种状态, 中断了平稳的变化[1]。人类活动导致的湖泊富营养化造成其透明度和沉水植物的减少, 而浮游藻类急剧增加, 湖泊呈现浑浊状态。在湖泊生态系统发生状态转变之前, 营养物质浓度的增加并不会对水体的透明度产生显著影响, 而当营养物浓度继续升高达到湖泊状态突变的临界阈值时, 水体会突然从清澈变浑浊, 呈现非线性的变化。珊瑚礁中食草动物的减少导致肉质褐藻的增加, 造成珊瑚礁不可逆转的退化[45-46]。海洋中鱼类的捕捞强度会使生态系统发生灾难性变化, 单个关键物种的变化影响着系统中营养级内部的潜在机制, 可能导致群落构成发生重大变化[47]。
3.2 林地系统林地中, 木本植物的幼苗被食草动物完全消灭的概率很小, 因此幼苗能够成长为成年林, 林地建立后就很难被破坏。然而树木的过度砍伐会导致林地被破坏, 并且破坏的林地很难再恢复到原初状态。Guttal和Jayaprakash[27]利用简单模拟系统展示了系统由无采伐到过度采伐的转变过程中方差增加和偏斜度达到峰值的过程。草地上偶尔的火灾会减少木本植物的覆盖, 有利于林下草本植物物种的发育, 而过度放牧会导致草本植物减少, 火灾的减少进而形成负反馈, 造成林下草本植物群落的退化[48-53]。这些负反馈驱动着生态系统组成成分在临界分岔点附近往复切换, 决定着生态系统趋于稳定状态的方向。
3.3 草地系统植被斑块的空间格局及斑块构型、属性等信息也可以用于指示植被状态跃变的早期预警信号。例如在草地生态系统, 草地退化过程是植被斑块化和生境破碎化过程。干旱半干旱区的草地经历放牧干扰, 导致植被的斑块化。随放牧强度增加, 植被经历由连接度高、数量少的大斑块向连接度低、数量增多的小斑块转变, 斑块大小和数量关系遵循幂函数变化法则。持续增强的放牧压力导致小斑块迅速消失, 此时斑块大小和数量的幂函数关系出现显著偏离, 预示着草地向荒漠迅速转变[26]。与此相似, 干旱、半干旱区草地灌丛化导致的荒漠化也是植物与水分资源在空间分布上权衡关系的结果。降水的减少使具有深根系的灌木在利用深层地下水方面更具优势, 逐渐形成了以灌木为优势的斑块结构。进而使得土壤养分资源等向植被斑块的聚集与再分配, 在空间上形成了资源高度异质性的格局。最终通过植物-植物、植物-环境之间的正反馈来维持空间构型上的相对稳定。一旦这种正反馈关系受到外界因素的破坏, 系统就会迅速瓦解为高同质性的荒漠生态系统[26]。因此植被自组织下形成的形状规则、大小均匀的斑块空间格局被认为是植被由平缓退化迅速向荒漠化状态跃变的早期预警信号。景观尺度上斑块结构及模型模拟研究已经证实了斑块空间结构对生态系统状态跃变的预警作用。
3.4 空间结构形成的自组织机制斑块化、生态系统状态突变的潜在机制是消费者与限制性资源之间的正反馈, 因为可利用性资源的多寡可以解释生态系统空间结构的多样性[26]。空间上的自组织并非作用于任何系统, 而是表现在小尺度的斑块与资源的相互作用, 可利用资源由多向寡的变化体现了系统由特有空间结构向高同质性状态的转变, 也即从一种稳态向另一种稳态的转变。资源贫瘠化程度的加剧推动消费者与有限资源在空间上重新分配和聚集, 导致资源与消费者在空间上的自组织斑块化格局的形成。气候变化和环境压力驱动的资源越有限, 在资源-消费者正反馈作用下越趋于形成规则、均匀的斑块结构, 表现出大尺上的斑块规则分布的景观格局。从能量耗散角度讲, 规则的斑块构型具有较低的周长-面积比, 能够减少植被与压力环境的接触, 减少内部能量散失, 减少植被经受严酷环境侵蚀的几率等等[54]。而随着环境扰动的增强, 一旦资源的匮乏达到了临界阈值, 资源-消费者之间的正反馈被破坏, 消费者从系统中丢失, 自组织斑块消失, 生态系统就会朝着高同质性的系统转变。一旦跃变到另一稳态的生态系统, 即使环境资源的有效性增加, 系统也不能在短期内恢复到原初状态, 即“滞后现象”。因此自组织斑块化可以预测生态系统状态的跃变。自组织斑块化与资源可利用性机制在很多生态系统中被观察到, 如半干旱生态系统、萨王纳、泥炭地生态系统等, 点状、条带状、环状等规则构型的斑块空间格局是植物与受限的水分、养分资源正反馈作用的结果[36]。干旱、低温、牧压、土地利用、人类消费(收获率)等环境压力是生态系统状态转变的主要驱动力。因此, 资源可利用性机制, 即植物-资源正反馈机制, 是自组织斑块化作为生态系统状态跃变早期预警信号的理论基础。建立自组织斑块化与生态系统状态跃变二者之间的联系填补了目前早期预警信号理论的空白, 对于生态系统的保护和管理也具有重要的现实意义。
4 挑战与展望目前关于生态系统状态跃变预警的研究有很多成果出现, 但是仍然有一些未知和亟待解决的问题。首先, 尺度是研究生态系统的一个重要因素, 对确定早期预警信号的有效性和适用性至关重要。尺度与选择基于多度的时间序列和空间模式的早期预警信号的研究方法息息相关。在空间模式的早期预警信号方法中, 选择正确的尺度有可能会明显改变空间信号的可靠性。在基于多度的时间序列早期预警信号中, 时间尺度又与生态系统的发展进程有关。在各类预警信号方法运用于种群、群落和生态系统之前, 了解尺度对这些信号的可靠性的影响, 从而选择正确的尺度来应用这类预警信号方法。种群不同的发展阶段可能有不同的预警信号, 在选取不同的方法测量生态系统状态转变的预警信号前, 应该确定好时空尺度, 选择不同的方法。其次, 生态系统的复杂性也会影响我们测量早期预警信号, 在正反馈相关的简单生态系统中, 可以使用基于临界减速现象的信号。在复杂的生态系统中, 基于临界减速的信号更难监测, 需要采用其他方法。探索新的方法仍然是状态突变研究的重要任务。另外, 在实际收集早期预警信号数据的过程中, 存在收集数据质量差的问题, 同时观测数据较少, 尤其是长期和大尺度的观测数据极其缺乏, 这在很大程度上限制了系统跃变理论的进一步发展与应用, 也限制了对时间尺度和空间尺度早期信号的验证。基于偏斜率的空间信号尚未应用于生态数据, 有广阔的未来发展前景。此外, 模型是系统跃变及其早期信号的研究中重要的研究方法, 模型的采用、算法等不同, 得到的结果也不同, 因此将强模型之间的比对与校验是早期预警信号研究的重中之重。
生态系统状态跃变的早期预警信号是否能被可靠检测到, 而这个检测时间是否能够确保通过人为干预阻断生态系统状态突变, 这是预测生态学面临和思考的问题。生态系统一直在受人类活动的干扰, 这使得生态系统有相对不退化和退化之分, 在已退化的系统检测到预警信号是否可靠, 其适用范围如何等问题需要认真界定。而以植物性状动态变化为早期预警信号, 不能确定植物物种性状的变化是生态系统突变前稳定性丧失的预警信号还是植物适应新环境所产生的突变。因此, 以植物性状动态变化的早期预警信号需要进一步量化, 除形态大小的变化外还可以考虑遗传多样性等信号。加强物种性状、斑块空间构型、斑块属性及自组织斑块化在资源有效性梯度下的变化趋势是生态系统早期预警信号研究需要回答的紧迫问题。另外, 生态系统功能的变化与结构上的变化存在时滞性, 对环境压力的响应存在显著的异步性, 因此, 在利用生态系统结构信号进行早期预警的同时, 了解生态系统多功能性变化的多阈值势在必行[55-57]。
| [1] |
Scheffer M, Carpenter S, Foley J A, Folke C, Walker B. Catastrophic shifts in ecosystems. Nature, 2001, 413(6856): 591-596. DOI:10.1038/35098000 |
| [2] |
Clements C F, Ozgul A. Indicators of transitions in biological systems. Ecology Letters, 2018, 21(6): 905-919. DOI:10.1111/ele.12948 |
| [3] |
Costanza R, d'Arge R, De Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O'Neill R V, Paruelo J, Raskin R G, Sutton P, van den Belt M. The value of the world's ecosystem services and natural capital. Nature, 1997, 387(6630): 253-260. DOI:10.1038/387253a0 |
| [4] |
Holling C S. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics, 1973, 4: 1-23. DOI:10.1146/annurev.es.04.110173.000245 |
| [5] |
Pimm S L. The complexity and stability of ecosystems. Nature, 1984, 307(5949): 321-326. DOI:10.1038/307321a0 |
| [6] |
Donohue I, Petchey O L, Montoya J M, Jackson A L, McNally L, Viana M, Healy K, Lurgi M, O'Connor N E, Emmerson M C. On the dimensionality of ecological stability. Ecology Letters, 2013, 16(4): 421-429. DOI:10.1111/ele.12086 |
| [7] |
De Keersmaecker W, Lhermitte S, Tits L, Honnay O, Somers B, Coppin P. A model quantifying global vegetation resistance and resilience to short-term climate anomalies and their relationship with vegetation cover. Global Ecology and Biogeography, 2015, 24(5): 539-548. DOI:10.1111/geb.12279 |
| [8] |
Radchuk V, De Laender F, Cabral J S, Boulangeat I, Crawford M, Bohn F, De Raedt J, Svenning C S J C, Thonicke K, Schurr F M, Kramer-Schadt V G S. The dimensionality of stability depends on disturbance type. Ecology Letters, 2019, 22(4): 674-684. DOI:10.1111/ele.13226 |
| [9] |
Holling C S. The strategy of building models of complex ecological systems//Watt K E F, ed. Systems Analysis in Ecology. New York and London: Academic Press, 1966: 195-214.
|
| [10] |
Dakos V, Carpenter S R, Brock W A, Ellison A M, Guttal V, Ives A R, Kéfi S, Livina V, Seekell D A, van Nes E H, Scheffer M. Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data. PLoS One, 2012, 7(7): e41010. DOI:10.1371/journal.pone.0041010 |
| [11] |
Matsumoto G, Kunisawa T. Critical slowing-down near the transition region from the resting to time-ordered states in squid giant axons. Journal of the Physical Society of Japan, 1978, 44(3): 1047-1048. DOI:10.1143/JPSJ.44.1047 |
| [12] |
Dakos V, Scheffer M, van Nes E H, Brovkin V, Petoukhov V, Held H. Slowing down as an early warning signal for abrupt climate change. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(38): 14308-14312. DOI:10.1073/pnas.0802430105 |
| [13] |
Drake J M, Griffen B D. Early warning signals of extinction in deteriorating environments. Nature, 2010, 467(7314): 456-459. DOI:10.1038/nature09389 |
| [14] |
Carpenter S R, Cole J J, Pace M L, Batt R, Brock W A, Cline T, Coloso J, Hodgson J R, Kitchell J F, Seekell D A, Smith L, Weidel B. Early warnings of regime shifts:a whole-ecosystem experiment. Science, 2011, 332(6033): 1079-1082. DOI:10.1126/science.1203672 |
| [15] |
Tirabassi G, Viebahn J, Dakos V, Dijkstra H A, Masoller C, Rietkerk M, Dekker S C. Interaction network based early-warning indicators of vegetation transitions. Ecological Complexity, 2014, 19: 148-157. DOI:10.1016/j.ecocom.2014.06.004 |
| [16] |
Brett T S, Drake J M, Rohani P. Anticipating the emergence of infectious diseases. Journal of the Royal Society Interface, 2017, 14(132): 20170115. DOI:10.1098/rsif.2017.0115 |
| [17] |
Clements C F, Blanchard J L, Nash K L, Hindell M A, Ozgul A. Body size shifts and early warning signals precede the historic collapse of whale stocks. Nature Ecology & Evolution, 2017, 1(7): 0188. |
| [18] |
Boettiger C, Hastings A. Early warning signals and the prosecutor's fallacy. Proceedings of the Royal Society B:Biological Sciences, 2012, 279(1748): 4734-4739. DOI:10.1098/rspb.2012.2085 |
| [19] |
Carpenter S R, Brock W A, Cole J J, Pace M L. A new approach for rapid detection of nearby thresholds in ecosystem time series. Oikos, 2014, 123(3): 290-297. DOI:10.1111/j.1600-0706.2013.00539.x |
| [20] |
Clements C F, Ozgul A. Including trait-based early warning signals helps predict population collapse. Nature Communications, 2016, 7(1): 10984. DOI:10.1038/ncomms10984 |
| [21] |
Petchey O L, Pontarp M, Massie T M, Kéfi S, Ozgul A, Weilenmann M, Palamara G M, Altermatt F, Matthews B, Levine J M, Childs D Z, McGill B J, Schaepman M E, Schmid B, Spaak P, Beckerman A P, Pennekamp F, Pearse I S. The ecological forecast horizon, and examples of its uses and determinants. Ecology Letters, 2015, 18(7): 597-611. DOI:10.1111/ele.12443 |
| [22] |
Boettiger C, Ross N, Hastings A. Early warning signals:the charted and uncharted territories. Theoretical Ecology, 2013, 6(3): 255-264. DOI:10.1007/s12080-013-0192-6 |
| [23] |
Spanbauer T L, Allen C R, Angeler D G, Eason T, Fritz S C, Garmestani A S, Nash K L, Stone J R, Stow C A, Sundstrom S M. Body size distributions signal a regime shift in a lake ecosystem. Proceedings of the Royal Society B:Biological Sciences, 2016, 283(1833): 20160249. DOI:10.1098/rspb.2016.0249 |
| [24] |
Cheung W W L, Sarmiento J L, Dunne J, Frölicher T L, Lam V W Y, Palomares M L D, Pauly D. Shrinking of fishes exacerbates impacts of global ocean changes on marine ecosystems. Nature Climate Change, 2013, 3(3): 254-258. |
| [25] |
Ozgul A, Bateman A W, English S, Coulson T, Clutton-Brock T H. Linking body mass and group dynamics in an obligate cooperative breeder. Journal of Animal Ecology, 2014, 83(6): 1357-1366. DOI:10.1111/1365-2656.12239 |
| [26] |
Kéfi S, Guttal V, Brock W A, Carpenter S R, Ellison A M, Livina V N, Seekell D A, Scheffer M, van Nes E H, Dakos V. Early warning signals of ecological transitions:methods for spatial patterns. PLoS One, 2014, 9(3): e92097. DOI:10.1371/journal.pone.0092097 |
| [27] |
Guttal V, Jayaprakash C. Changing skewness:an early warning signal of regime shifts in ecosystems. Ecology Letters, 2008, 11(5): 450-460. DOI:10.1111/j.1461-0248.2008.01160.x |
| [28] |
Dai L, Korolev K S, Gore J. Slower recovery in space before collapse of connected populations. Nature, 2013, 496(7445): 355-358. DOI:10.1038/nature12071 |
| [29] |
Kéfi S, Rietkerk M, van Baalen M, Loreau M. Local facilitation, bistability and transitions in arid ecosystems. Theoretical Population Biology, 2007, 71(3): 267-400. DOI:10.1016/j.tpb.2006.11.006 |
| [30] |
Boettiger C, Hastings A. Quantifying limits to detection of early warning for critical transitions. Journal of the Royal Society Interface, 2012, 9(75): 2527-2539. DOI:10.1098/rsif.2012.0125 |
| [31] |
Heffley T J, Tyre A J, Blankenship E E. Statistical indicators and state-space population models predict extinction in a population of bobwhite quail. Theoretical Ecology, 2013, 6(3): 319-331. DOI:10.1007/s12080-013-0195-3 |
| [32] |
Seekell D A, Carpenter S R, Pace M L. Conditional heteroscedasticity as a leading indicator of ecological regime shifts. The American Naturalist, 2011, 178(4): 442-451. DOI:10.1086/661898 |
| [33] |
Dakos V, Soler-Toscano F. Measuring complexity to infer changes in the dynamics of ecological systems under stress. Ecological Complexity, 2016, 32: 144-155. |
| [34] |
Biggs R, Carpenter S R, Brock W A. Turning back from the brink:detecting an impending regime shift in time to avert it. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(3): 826-831. DOI:10.1073/pnas.0811729106 |
| [35] |
Sundstrom S M, Eason T, Nelson R J, Angeler D G, Barichievy C, Garmestani A S, Graham N A J, Granholm D, Gunderson L, Knutson M, Nash K L, Spanbauer T, Stow C A, Allen C R. Detecting spatial regimes in ecosystems. Ecology Letters, 2017, 20(1): 19-32. |
| [36] |
Rietkerk M, Dekker S C, de Ruiter P C, van de Koppel J. Self-organized patchiness and catastrophic shifts in ecosystems. Science, 2004, 305(5692): 1926-1929. DOI:10.1126/science.1101867 |
| [37] |
Kéfi S, Rietkerk M, Alados C L, Pueyo Y, Papanastasis V P, ElAich A, de Ruiter P C. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature, 2007, 449(7159): 213-217. DOI:10.1038/nature06111 |
| [38] |
Deblauwe V, Couteron P, Lejeune O, Bogaert J, Barbier N. Environmental modulation of self-organized periodic vegetation patterns in Sudan. Ecography, 2011, 34(6): 990-1001. DOI:10.1111/j.1600-0587.2010.06694.x |
| [39] |
Lade S J, Gross T. Early warning signals for critical transitions:a generalized modeling approach. PLoS Computational Biology, 2012, 8(2): e1002360. DOI:10.1371/journal.pcbi.1002360 |
| [40] |
Yin Z, Dekker S C, Rietkerk M, van den Hurk B J J M, Dijkstra H A. Network based early warning indicators of vegetation changes in a land-atmosphere model. Ecological Complexity, 2016, 26: 68-78. DOI:10.1016/j.ecocom.2016.02.004 |
| [41] |
Vandermeer J, Perfecto I. A keystone mutualism drives pattern in a power function. Science, 2006, 311(5763): 1000-1002. DOI:10.1126/science.1121432 |
| [42] |
Burov S, Ali Tabei S M, Huynh T, Murrell M P, Philipson L H, Rice S A, Gardel M L, Scherer N F, Dinner A R. Distribution of directional change as a signature of complex dynamics. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(49): 19689-19694. DOI:10.1073/pnas.1319473110 |
| [43] |
Liu Q X, Doelman A, Rottschäfer V, de Jager M, Herman P M J, Rietkerk M, van de Koppel J. Phase separation explains a new class of self-organized spatial patterns in ecological systems. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(29): 11905-11910. DOI:10.1073/pnas.1222339110 |
| [44] |
de Paoli H, van der Heide T, van den Berg A, Silliman B R, Herman P M J, van de Koppel J. Behavioral self-organization underlies the resilience of a coastal ecosystem. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(30): 8035-8040. DOI:10.1073/pnas.1619203114 |
| [45] |
Weisner S E B, Strand J A, Sandsten H. Mechanisms regulating abundance of submerged vegetation in shallow eutrophic lakes. Oecologia, 1997, 109(4): 592-599. DOI:10.1007/s004420050121 |
| [46] |
Carpenter S R, Ludwig D, Brock W A. Management of eutrophication for lakes subject to potentially irreversible change. Ecological Applications, 1999, 9(3): 751-771. |
| [47] |
Meijer M L, Jeppesen E, Van Donk E, Moss B, Scheffer M, Lammens E, van Nes E, van Berkum J A, de Jong G J, Faafeng B A, Jensen J P. Long-term responses to fish-stock reduction in small shallow lakes:interpretation of five-year results of four biomanipulation cases in the Netherlands and Denmark. Hydrobiologia, 1994, 275(1): 457-466. |
| [48] |
Walker B H. Diversity and stability in biodiversity conservation. In:Weston D, Pearl M, eds. Conservation Biology for the Twenty-First Century. Oxford: Oxford University Press, 1989: 121-130.
|
| [49] |
Wilson J B, Agnew A D Q. Positive-feedback switches in plant communities. Advances in Ecological Research, 1992, 23: 263-336. DOI:10.1016/S0065-2504(08)60149-X |
| [50] |
Dublin H T, Sinclair A R E, McGlade J. Elephants and fire as causes of multiple stable states in the Serengeti-Mara woodlands. Journal of Animal Ecology, 1990, 59(3): 1147-1164. DOI:10.2307/5037 |
| [51] |
Holmgren M, Scheffer M. El Niño as a window of opportunity for the restoration of degraded arid ecosystems. Ecosystems, 2001, 4(2): 151-159. DOI:10.1007/s100210000065 |
| [52] |
Walker B H. Rangeland ecology:understanding and managing change. Ambio, 1993, 22(2/3): 80-87. |
| [53] |
Ludwig D, Walker B, Holling C S. Sustainability, stability, and resilience. Conservation Ecology, 1997, 1(1): 7. |
| [54] |
Crotty S M, Sharp S J, Bersoza A C, Prince K D, Cronk K, Johnson E E, Angelini C. Foundation species patch configuration mediates salt marsh biodiversity, stability and multifunctionality. Ecology Letters, 2018, 21(11): 1681-1692. DOI:10.1111/ele.13146 |
| [55] |
Allan E, Weisser W W, Fischer M, Schulze E D, Weigelt A, Roscher C, Baade J, Barnard R L, Beßler H, Buchmann N, Ebeling A, Eisenhauer N, Engels C, Fergus A J F, Gleixner G, Gubsch M, Halle S, Klein A M, Kertscher I, Kuu A, Lange M, Le Roux X, Meyer S T, Migunova V D, Milcu A, Niklaus P A, Oelmann Y, Pašalić E, Petermann J S, Poly F, Rottstock T, Sabais A C W, Scherber C, Scherer-Lorenzen M, Scheu S, Steinbeiss S, Schwichtenberg G, Temperton V, Tscharntke T, Voigt W, Wilcke W, Wirth C, Schmid B. A comparison of the strength of biodiversity effects across multiple functions. Oecologia, 2013, 173(1): 223-237. DOI:10.1007/s00442-012-2589-0 |
| [56] |
Dooley Á, Isbell F, Kirwan L, Connolly J, Finn J A, Brophy C. Testing the effects of diversity on ecosystem multifunctionality using a multivariate model. Ecology Letters, 2015, 18(11): 1242-1251. DOI:10.1111/ele.12504 |
| [57] |
Manning P, van der Plas F, Soliveres S, Allan E, Maestre F T, Mace G, Whittingham M J, Fischer M. Redefining ecosystem multifunctionality. Nature Ecology & Evolution, 2018, 2(3): 427-436. |