 2020, Vol. 40
2020, Vol. 40文章信息
- 马雄忠, 王新平
- MA Xiongzhong, WANG Xinping
- 阿拉善高原2种荒漠植物根系构型及生态适应性特征
- Root architecture and adaptive strategy of two desert plants in the Alxa Plateau
- 生态学报. 2020, 40(17): 6001-6008
- Acta Ecologica Sinica. 2020, 40(17): 6001-6008
- http://dx.doi.org/10.5846/stxb201905030894
-
文章历史
- 收稿日期: 2019-05-03
- 网络出版日期: 2020-07-09





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
根系是植物与外界环境进行物质和能量交换的关键器官之一[1-3], 根系构型指植物根系在土壤中的分布和排列[4]。根系构型不止受遗传因素控制, 不同的生境也会影响根系构型的表达[5]。根系构型一定程度上反映了植物对环境的适应性及植被的退化和演替状况[6]。然而, 由于根系生长于地下, 不能直观的对根系构型进行认识和了解[3, 7]。以往的研究较多的是通过外形、根冠比、根重等指标反映根系生长情况, 而对根系的分支和结构特征研究较少[8-9]。
20世纪70年代, 分形几何的诞生为根系构型研究提供了一种新的方法。根系是较为明显的分形结构[10], 运用分形理论研究根系, 可以定量描述根系几何形态、分形特征及扩展范围等[11]。分形维数和分形丰度是根系分形结构的两个重要的参数, 分形维数可以说明根系发育程度及分支能力等;分形丰度反映根系分布范围、密度及资源竞争力等。分形丰度和分形维数密切相关, 综合体现了植物的营养吸收、传输、资源利用能力及碳消耗特点[10]。20世纪80年代, 几何拓扑学被引入了根系研究[12-13], 根系拓扑结构与根系连接数量、分支情况相关, 可以用来描述根系营养吸收和固定能力[8, 14]。相较根系形态指标, 拓扑特征能更准确的反映根系构型, 因此近年来学者们对拓扑结构的研究日益增多[15]。杨小林等[16]研究了沙漠腹地3种植物根系拓扑特征与生境的关系, 单立山等[17]研究了河西走廊中部两种荒漠植物根系拓扑和分型特征, 杨振亚等[18]研究了干旱对杉木幼苗根系拓扑结构的影响。拓扑结构和分形特征能够反映根系的分支模式和功能特点, 将两种方法综合应用于根系研究能更准确的反映根系构型特征和生态适应策略。
红砂(Reaumuria soongarica)和珍珠猪毛菜(Salsola passerina)是超旱生植物, 在荒漠区, 几乎能生长在所有的土质上, 是中国荒漠地区分布最广的地带性建群种之一。两种荒漠植物也是阿拉善高原分布较广的两种优势植物, 两者常互为共建种组成混生群落, 混生群落地上部分的“成丛性”分布, 即通过聚集分布以降低蒸腾失水等生理胁迫, 地下根系分布稀疏, 一般情况下红砂根系分布更深。其抗寒抗旱能力强, 且有集沙固沙作用, 对干旱荒漠区植被恢复与重建、水土流失治理等具有重要作用[19-20]。近年来随着人类干扰加剧, 导致阿拉善生态环境退化, 红砂和珍珠猪毛菜的生存环境受到极大的影响[21], 现有的研究较少涉及取样困难的地下部分, 而两种荒漠植物作为沙漠特殊环境的良好适应者, 了解其根系构型特征是探讨环境变化条件下该地区植物根系-土壤互作的重要途径, 对于沙漠地区植被恢复具有重要意义。鉴于此, 本文从根系形态特征、分形特征和拓扑特征三方面刻画红砂和珍珠猪毛菜的根系构型特点, 分析红砂和珍珠猪毛菜在形态结构方面对干旱的响应调节, 探讨其对阿拉善高原西南缘干旱荒漠生境的生态适应性。该研究对揭示干旱区植物共存机制具有重要科学意义, 将为揭示阿拉善高原西南缘灌木多样性成因提供理论依据, 并对该地区生态保护和恢复提供基础资料。
1 材料和方法 1.1 研究区概况研究区域位于阿拉善高原西南缘, 河西走廊东段, 祁连山北麓, 其地理位置为101°34′E, 38°46′N。该区属温带大陆性极干旱气候, 常年干燥, 年均降水量119.5 mm, 年均蒸发量2722 mm。冬冷夏热, 昼夜和四季温差大, 冬夏季长, 春秋季短, 春季升温快, 秋季降温迅速。年均气温9.4℃, 1月最低, 平均-7.0℃;7月最高, 平均24.2℃。年均日照数2991.7h, 年均无霜期170 d。雨量少, 光照强, 风沙多, 地表水资源匮乏。地下水位低, 一般低于地面以下-40 m。土壤类型为灰棕漠土, 土壤层有效厚度较小, 持水持肥能力弱。土壤体积密度为1.45 g/cm3, 沙粒、粉粒和粘粒含量分别为62%、20%和18%, 土壤总C、总N和总P含量分别为12.85、0.47 g/kg和0.77 g/kg, 有机质含量为4.84 g/kg, 总盐含量为3.27 g/kg。地带性植被以天然珍珠猪毛菜、红砂、盐爪爪(Kalidium foliatum)、骆驼蓬(Peganum harmala L.)等旱生灌木为主。研究区总植被盖度为18.6%, 其中红砂和珍珠猪毛菜分别占总盖度的61.8%和6.5%, 两种植被组成混生群落, 是区域内分布最广泛的荒漠植被, 对两种植被的研究可以代表区域内植被根系构型的基本特征。
1.2 样品采集与处理2018年全年降水量108.9 mm, 其中4—7月降水量占全年降水量的56.7%。于2018年7月下旬, 在阿拉善高原西南缘的红砂-珍珠猪毛菜自然混生群落中选择地势平坦、植物分布相对较为均匀的区域设置3个样方, 样方间隔为1000 m, 样方大小设置为10 m × 10 m, 对样方内红砂和珍珠猪毛菜的地上部分指标(株高、冠幅、基径)进行调查, 统计确定标准样株。红砂标准样株株高、冠幅和基径分别为24.77 cm、1653.13 cm-2和13.1 mm, 珍珠猪毛菜标准株株高、冠幅和基径分别为12.43 cm、389.31 cm-2和13.95 mm。参照标准样株, 在每个样方内选择红砂和珍珠猪毛菜各2株, 实验共采集红砂和珍珠各6株, 用毛刷、铁铲等工具小心地挖出根系, 保持根系的自然分布位置, 采用25 cm × 17 cm的网格纸按1:10比例绘制根系的俯视图, 记录粗根(φ>2 mm)的内部、外部连接数量和连接长度。记录完毕后, 将采集的根系带回实验室, 用清水洗净根上附着的泥沙, 用Epson扫描仪扫描, 用Win-RhIZO根系分析软件分析根长、根表面积。将根系按分布深度10 cm为一层进行划分, 红砂采样深度为60 cm, 分6层;珍珠采样深度为40 cm, 分4层, 每一层分为粗根和细根(φ≤2 mm), 分别装于不同的信封标记, 在65℃的烘箱中烘干至恒重, 用百分之一天平称量各层根系生物量。
1.3 分析方法 1.3.1 根系形态参数的确定根据Win-RhIZO测定的根长和根表面积, 以及烘干称重测得的根系生物量计算红砂和珍珠猪毛菜的根系比根长(SRL)、比表面积(SRA)等参数。计算公式如下:
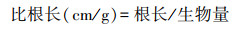
|
(1) |
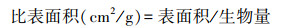
|
(2) |
根系消弱系数β根据植物根系垂直分布的非线性模型计算[22], 计算公式如下:
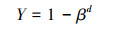
|
(3) |
式中, Y表示从土壤表面到某深度d(cm)的根系生物量累计百分比, β为根系消弱系数, 通过拟合测量值, 得到β的值。β范围为0—1, 越接近1, 表示根系在较深的土层分布比例较多;越接近0, 表示根系在浅层分布比例较大。
R50和R90分别表示50%和90%根系生物量的分布深度。
1.3.2 拓扑参数的计算Fitter等[12, 23-24]和Bouma等[25]提出根系拓扑结构存在鱼尾形分支和叉状分支两种极端模式(图 1), 并提出拓扑指数表达式:
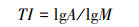
|
(4) |

|
| 图 1 根系拓扑结构示意图 Fig. 1 The schematic diagram of topological structure of root system |
式中, TI为拓扑指数, A为最长通道(连接最多的通道)的内部连接数, M为外部连接总数。TI的值越接近1表明A和M是近似相等的, 即根系分支较少, 根系越近似鱼尾形分支结构, 越接近0.5表明根系存在相对更多的外部连接, 根系越近似叉状分支结构。
Oppelt等[13]为了说明根系拓扑结构的中间过度形式, 提出了新的拓扑参数。其表达式为:
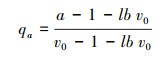
|
(5) |
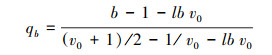
|
(6) |
式中, qa和qb分别表示a和b的修正值;a为最长通道内部连接总数, 其值与Fitter模型的A相等;b为平均拓扑长度(为从基部到根终端连接数量称);lbv0=lnv0/ln2, 其中v0为外部连接总数, 等同于Fitter模型的M。鱼尾形分支的qa=qb=1, 叉状分支的qa=qb=0。
1.3.3 分形参数的计算分形参数采用计盒维数法确定。在根系俯视图上绘制正方形, 其边长设定为18.4 cm, 依次将其分成边长为r=18.4/2n(n为0-5)的正方形, 将每个边长下根系穿过的正方形数目记为Nr, r和Nr存在对应关系。分别取lgr和lgNr为横纵坐标做图, 其回归方程为:lgNr=FDlgr+lgK, 其中, FD表示分形维数, 1 < FD < 2, 根系分支越多, 根的分形维数越高, 表示根系越发达;lgK表示分形丰度, 其值越大, 表示在土壤中扩展的体积越大[26-27]。
1.4 数据分析数据分析在SPSS 23.0中进行, 采用t检验对植物根系相关参数进行差异性分析, 显著性水平设定为α=0.05。采用origin9.0作图。相关图表制作在Excel中完成。
2 结果与分析 2.1 根系形态特征阿拉善西南缘红砂根系的水平分布范围平均为119.0 cm, 垂直分布深度平均为62.3 cm, 平均根幅/平均根深比率为1.9, 根系水平分布占优势。珍珠猪毛菜根系的水平分布范围平均为70.1 cm, 垂直分布深度平均为40.1 cm, 平均根幅/平均根深比率为1.8, 根系水平分布占优势。红砂R50和R90分别为15.1 cm和30.9 cm, 根系消弱系数为0.94, 珍珠猪毛菜R50和R90分别为7.3 m和17.8 cm, 根系消弱系数为0.89, 表明红砂根系主要分布在0—30 cm土层中, 珍珠猪毛菜主要分布在0—20 cm土层中, 红砂密集根深显著大于珍珠猪毛菜(表 1)。
| 物种 Species | 根系深度 Root depth/ cm | 根系水平范围 Root horizontal range/cm | 根水平范围/ 根深度 Root horizontal range/Root depth | R50/cm | R90/cm | β |
| 红砂R. soongorica | 62.3±8.5a | 119.0±21.4a | 1.9±0.4 | 15.1±1.2 | 30.9±2.3 | 0.94±0.01a |
| 珍珠猪毛菜S. passerina | 40.1±5.5b | 70.1±2.9b | 1.8±0.3 | 7.3±0.9 | 17.8±1.4 | 0.89±0.01b |
| R50: 50%根系生物量的分布深度; R90: 90%根系生物量的分布深度; β:根系消弱系数; 同列不同小写字母表示物种间差异显著(P<0.05);表中数据为平均值±标准误差(n=6) | ||||||
阿拉善高原西南缘红砂根系分布深度和总根系生物量均大于珍珠猪毛菜, 且差异均达到显著水平(P<0.05), 红砂单株根系生物量平均值为41.7 g, 珍珠猪毛菜单株根系生物量平均值为18.6 g, 两种荒漠植物根系生物量均随着土壤深度的增加呈递减趋势。在相同的土壤深度, 红砂根系生物量大于珍珠猪毛菜根系生物量。红砂吸收根(细根)主要分布在10—40 cm的土层, 而珍珠猪毛菜细根生物量主要分布在10—20 cm土层(图 2)。

|
| 图 2 2种荒漠植物根系生物量的垂直变化特征 Fig. 2 The vertical distributions of biomass of two desert plants 图中数据为平均值±标准误差(n=6) |
红砂细根的比根长和比表面积(SRL=107.4 cm/g, SRA=21.2 cm2/g)小于珍珠猪毛菜(SRL=158.7 cm/g, SRA=25.6 cm2/g), 粗根的比根长和比表面(SRL=7.4 cm/g, SRA=5.5 cm2/g)大于珍珠猪毛菜(SRL=4.3 cm/g, SRA=4.0 cm2/g), 总比根长(SRL红=21.3 cm/g, SRL珍=22.4 cm/g)和总比表面积(SRA红=7.6 cm/g, SRA珍=6.5 cm/g)两者差异不显著(P>0.05), 说明在小于2 mm的细根范围内, 珍珠猪毛菜的平均直径更小, 而在大于2 mm的粗根范围内, 红砂的平均直径更小(图 3)。

|
| 图 3 2种荒漠植物根系构型参数 Fig. 3 Parameters of root architecture of two desert plants qa: a的修正值; qb: b的修正值; TI:拓扑指数; 相同参数不同小写字母表示差异显著(P < 0.05);图中数据为平均值±标准误差(n=6) |
阿拉善高原西南缘红砂的最长通道内部连接总数a(A), 根系所有外部链接总数v0(M)、平均拓扑长度b和所有连接总数Pe均小于珍珠猪毛菜(表 2)。红砂的修正拓扑指数qa=0.52、qb=0.49, 拓扑指数TI=0.86, 珍珠猪毛菜的修正拓扑指数qa=0.77、qb=0.73, 拓扑指数TI=0.94(图 3), 两者均趋向于鱼尾形分支, 表明两种荒漠植物根系分支结构较简单, 主根明显且分支较少。两种荒漠植物根系修正拓扑指数存在差异显著(P < 0.05), 红砂的qa和qb小于珍珠猪毛菜, 说明红砂根系的拓扑结构比珍珠猪毛菜复杂, 有相对更多的次级分支。独立样本T检验表明, 红砂和珍珠猪毛菜拓扑指数差异不显著(P>0.05), 表明在阿拉善高原西南缘的干旱荒漠生境中, 两种植物的根系分支模式表现出趋同性。
| 物种Species | a(A) | v0(M) | b | Pe | 总根长/cm Total root length | 平均连接长度/cm Average link length |
| 红砂R. soongorica | 9.3±1.3 | 14.3±3.3 | 6.4±0.23 | 99.6±13.3 | 318.5±25.4 | 11.4±1.5 |
| 珍珠猪毛菜S. passerina | 12.0±0.5 | 15.0±2.5 | 7.9±0.45 | 119.0±9.8 | 284.5±16.9 | 12.9±1.2 |
| a(A):最长根系通道内部连接总数; v0(M):为根系所有外部连接总数; b:根系平均拓扑长度; Pe:根系基部到根端部通道的所有连接总数; 表中数据为平均值±标准误差(n=6) | ||||||
如图 3所示, 红砂根系的总长度为318.5 cm, 平均连接长度为11.4 cm;珍珠猪毛菜根系总长度为284.6 cm, 平均连接长度为12.9 cm, 独立样本T检验表明, 两者总长度和平均连接长度差异不显著(P < 0.05)。相较于同地区的草本植物, 两种荒漠灌木根系的总长度和平均连接长度更大, 扩大了根系的营养空间。
2.3 根系分形特征红砂和珍珠猪毛菜根系分形特征明显。2种植物分形维数分别为1.488和1.422(表 3), 独立样本T检验表明, 二者根系分形维数差异不显著(P>0.05), 分支结构均比较简单。红砂的分形维数略大于珍珠猪毛菜, 表明红砂相比珍珠猪毛菜根系分支更多。红砂和珍珠猪毛菜分形丰度分别为1.855和1.744, 独立样本T检验表明, 二者差异不显著(P>0.05), 空间扩展能力均较强。红砂的分形丰度略大于珍珠猪毛菜, 表明红砂根系在土壤中拓展的体积大于珍珠猪毛菜。
| 物种 Species | 分形维数 Fractal dimension | 分形丰度 Fractal abundance | 决定系数 R2 |
| 红砂R. soongorica | 1.488±0.122a | 1.855±0.089a | 0.984 |
| 珍珠猪毛菜 S. passerina | 1.422±0.054a | 1.774±0.033a | 0.949 |
| 同列不同小写字母表示物种间差异显著(P<0.05)。表中数据为平均值±标准误差(n=6) | |||
植物根系空间分布特征受遗传因素和外界环境的双重影响, 为了适应不同生境的水分、养分情况, 植物根系分布随环境发生一定的改变, 表现出较高的可塑性[17, 28-29]。作为干旱荒漠区广泛分布的植物种, 陈永乐等[30]对腾格里沙漠东南缘的红砂、珍珠猪毛菜根系研究发现, 其根系分布深度可达2.9 m, 表现出了深根型的特点。然而, 本研究对阿拉善高原西南缘红砂、珍珠猪毛菜的研究发现, 其最大分布深度分别为70 cm和40 cm, 根幅/根深分别为1.9和1.8, 且90%的生物量均集中于30 cm以上的土层中, 2种荒漠植物根系均表现为水平分布占优势的浅层根。这与单立山等[20]对张掖、酒泉等地红砂、珍珠猪毛菜根系分布特征调查结果相一致。这些根系分布的差异可能是遗传因素造成的, 但更大的可能是与不同生境下不同的土壤和水分条件有关, 腾格里沙漠东南缘多年平均降水量186.2 mm, 土壤以细沙粒为主[30], 其年均降水量显著高于本研究区的119.5 mm, 而根系在砂质土壤内的延展阻力也小于研究区的灰棕漠土。在降雨量和土壤性质与本研究区相近的张掖、酒泉等地, 红砂和珍珠猪毛菜表现出与本研究区相似的根系分布特征。张道远等[31]认为, 浅层化的根能够更高效的利用资源, 有利于种群的扩大。本研究中, 红砂和珍珠猪毛菜的根系分布浅层化, 且水平分布占优, 扩大了对浅表层土壤空间的占有和利用, 能够更高效的利用表层土壤水, 这可能是两种植物对干旱荒漠区极端环境的适应策略。两种荒漠植物根系在浅层化分布的同时, 还表现出错位分布的特点, 体现了空间生态位的分离, 减少了红砂和珍珠猪毛菜的种间竞争, 这种空间格局也可能是2种荒漠植物适应干旱胁迫环境的重要策略。
一般认为, 植物可通过根系形态结构的改变来增强对资源的竞争力[32]。比根长和比表面积是根系形态结构的重要指标, 决定了根系吸收水分和养分的能力, 其值与养分吸收效率成正比[33]。红砂和珍珠猪毛菜均有较大的比根长和比表面积, 对比发现, 珍珠猪毛菜主要吸收根(φ < 2 mm)的比根长和比表面积大于红砂, 可能是珍珠猪毛菜根系相较红砂根系分布更浅, 能够利用浅表层土壤中的凝结水[34], 珍珠猪毛菜通过减小吸收根的平均直径来扩大比根长和比表面积, 以获取更多水分以适应干旱胁迫。而红砂主要利用较深层土壤水分, 其主根较发达, 吸收根平均直径相对较粗, 所以其比根长和比表面积更小。表明植物可通过增加比根长和比表面积促进根系对土壤水分和养分的吸收以适应竞争环境。
3.2 根系拓扑结构特征根系的几何拓扑结构与其对营养物质的竞争力密切相关。通常叉状根系次级分支多, 根系较密集, 内部竞争强, 鱼尾型分支则相反, 所以, 投入等量的碳, 叉状分支根系的分布范围小于鱼尾形分支, 不利于营养空间的占有, 因此相较叉状分支, 鱼尾形分支更适应于干旱贫瘠的生境[23-25]。杨小林等[16]对塔克拉玛干沙漠腹地的荒漠植物研究发现, 沙拐枣(Calligonum mongolicum Turcz.)和罗布麻(A. venetum L.)根系均接近鱼尾形分支, 其根系覆盖范围较大, 有利于水分和养分的吸收。本研究发现, 在阿拉善高原南缘干旱贫瘠的土壤环境中, 红砂和珍珠猪毛菜也趋向于鱼尾形分支模式, 具有分支简单, 次级分支少, 连接长度较大, 分布范围广的特点, 这种根系结构增加了两种荒漠植物对营养物质的获取能力, 是其适应当地自然环境的重要手段。然而, 也有研究表明叉状分支在贫瘠的土壤环境中也存在, 单立山等[17]对河西走廊中部的红砂和白刺(Nitraria tangutorum Bor.)研究发现, 其根系分支结构复杂, 分支模式接近叉状分支结构;杨小林等[16]研究发现, 塔克拉玛干沙漠腹地的柽柳(Tamarix chinensis Lour.)为叉状分支结构。这表明, 在干旱贫瘠的荒漠环境中, 植物可能存在两种不同的根系分支策略, 而分支策略的具体决定因素, 尚需深入研究。
3.3 根系分形维数分形理论可对根系分支的复杂程度及其在土壤中的扩展能力做定量研究[35]。分形维数能反映植物根系的发育程度和对胁迫环境的适应策略。在等量的碳输入下, 根系分支越简单, 其空间占有能力越强;反之, 空间占有能力越弱[10]。单立山等[17]对河西走廊地区的罗布麻进行研究发现, 其分形维数小, 通过简单的分形结构来适应干旱胁迫生境;而在半干旱气候的黄土丘陵地区生长的罗布麻具有较大的分形维数, 分支较复杂。本研究发现阿拉善高原西南缘红砂(FD=1.488)和珍珠猪毛菜(FD=1.422)分形维数较小, 分支结构相对简单, 表明在水分或养分贫瘠条件下根系偏向于减少分支, 扩大营养空间, 以确保获得生存所需的水分和养分。2种荒漠植物之间分形维数的差异可能与根系分布深度相关, 红砂根系分布较深, 土壤水分和养分条件相对较好, 致使其分形维数更大。根系分形丰度与根系的延展范围、拓展能力及对水分和养分的吸收效率成正相关[27]。本研究发现, 在阿拉善高原西南缘的干旱荒漠区, 红砂和珍珠猪毛菜均维持较大的分形丰度。红砂和珍珠猪毛菜根系分形维数和分形丰度的特点, 综合反映了其对阿拉善高原干旱荒漠区生境的适应策略。
4 结论通过对阿拉善高原西南缘红砂、珍珠猪毛菜根系的形态特征、拓扑结构和分形特征进行研究, 从根系构型特征方面揭示了荒漠植物对干旱贫瘠生境的适应策略。
(1) 阿拉善高原西南缘的红砂和珍珠猪毛菜根系均水平分布占优。混生的两种植物根系形成了分层现象, 红砂更深的根系分布有利于利用较深层的土壤水, 而珍珠猪毛菜更易于利用浅层土壤中的非降水性水资源(如凝结水), 这是红砂和珍珠猪毛菜混生群落稳定演替的重要生态机制。同时, 两种植物较大的根长和根表面积也有利于获取水分以适应干旱胁迫。
(2) 研究区红砂和珍珠猪毛菜根系均趋向于鱼尾形分支结构, 降低了物种根系内部对水分和养分资源的竞争。同时, 红砂根系拓扑参数均小于珍珠猪毛菜, 降低了红砂-珍珠猪毛菜混生群落的种间竞争。此外, 两种植物较大的根系连接长度有利于扩大对水分和养分的利用空间。
(3) 研究区的红砂和珍珠猪毛菜具有较低的分形维数和较高的根丰度, 表明其根系分支相对简单、发育程度较高, 且空间拓展能力较强, 有利于对营养空间的占有, 反映了其对阿拉善干旱荒漠生境的适应策略。
| [1] |
Dannowski M, Block A. Fractal geometry and root system structures of heterogeneous plant communities. Plant and Soil, 2005, 272(1/2): 61-76. |
| [2] |
鲍雅静, 曹明, 李政海, 郭鹏, 张靖, 秦洁. 羊草与大针茅根系构型对水分梯度响应的比较研究. 生态学报, 2019, 39(3): 1063-1070. |
| [3] |
邓磊, 关晋宏, 张文辉. 辽东栎幼苗根系形态特征对环境梯度的响应. 生态学报, 2018, 38(16): 5739-5749. |
| [4] |
Lynch J. Root architecture and plant productivity. Plant Physiology, 1995, 109(1): 7-13. |
| [5] |
单立山, 李毅, 张荣, 张正中, 种培芳. 降雨格局变化对白刺幼苗根系形态特征的影响. 生态学报, 2017, 37(21): 7324-7332. |
| [6] |
孙栋元, 赵成义, 王丽娟, 盛钰, 李菊艳. 荒漠植物构型研究进展. 水土保持研究, 2011, 18(5): 281-287. |
| [7] |
梁泉, 廖红, 严小龙. 植物根构型的定量分析. 植物学报, 2007, 24(6): 695-702. |
| [8] |
Glimskär A. Estimates of root system topology of five plant species grown at steady-state nutrition. Plant and Soil, 2000, 227(1/2): 249-256. DOI:10.1023/A:1026531200864 |
| [9] |
朱广龙, 陈许兵, 郭小倩, 焦秀荣, 周桂生. 酸枣根系结构可塑性对自然梯度干旱生境的适应机制. 生态学报, 2018, 38(16): 5810-5818. |
| [10] |
杨培岭, 罗远培. 冬小麦根系形态的分形特征. 科学通报, 1994, 39(20): 1911-1913. |
| [11] |
王义琴, 张慧娟, 白克智, 孙勇如. 分形几何在植物根系研究中的应用. 自然杂志, 1999, 21(3): 143-145, 146-146. |
| [12] |
Fitter A H. The topology and geometry of plant root systems:influence of watering rate on root system topology in Trifolium pratense. Annals of Botany, 1986, 58(1): 91-101. DOI:10.1093/oxfordjournals.aob.a087191 |
| [13] |
Oppelt A L, Kurth W, Godbold D L. Topology, scaling relations and Leonardo's rule in root systems from African tree species. Tree Physiology, 2001, 21(2/3): 117-128. |
| [14] |
Berntson G M. Topological scaling and plant root system architecture:developmental and functional hierarchies. New Phytologist, 1997, 135(4): 621-634. DOI:10.1046/j.1469-8137.1997.00687.x |
| [15] |
Martínez-Sánchez J, Ferrandis P, Trabaud L, Galindo R, Franco J A, Herranz J M. Comparative root system structure of post-fire Pinus halepensis Mill. and Cistus monspeliensis L saplings. Plant Ecology, 2003, 168(2): 309-320. |
| [16] |
杨小林, 张希明, 李义玲, 李绍才, 孙海龙. 塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略. 植物生态学报, 2008, 32(6): 1268-1276. |
| [17] |
单立山, 李毅, 任伟, 苏世平, 董秋莲, 耿东梅. 河西走廊中部两种荒漠植物根系构型特征. 应用生态学报, 2013, 24(1): 25-31. |
| [18] |
杨振亚, 周本智, 陈庆标, 葛晓改, 王小明, 曹永慧, 童冉, 石洋. 干旱对杉木幼苗根系构型及非结构性碳水化合物的影响. 生态学报, 2018, 38(18): 6729-6740. |
| [19] |
Su P X, Yan Q D, Xie T T, Zhou Z J, Gao S. Associated growth of C3 and C4 desert plants helps the C3 species at the cost of the C4 species. Acta Physiologiae Plantarum, 2012, 34(6): 2057-2068. DOI:10.1007/s11738-012-1003-1 |
| [20] |
单立山, 苏铭, 张正中, 王洋, 王珊, 李毅. 不同生境下荒漠植物红砂-珍珠猪毛菜混生根系的垂直分布规律. 植物生态学报, 2018, 42(4): 475-486. |
| [21] |
万炜, 颜长珍. 阿拉善高原生态环境退化研究进展. 地球环境学报, 2018, 9(2): 109-122. |
| [22] |
Gale M R, Grigal D F. Vertical root distributions of northern tree species in relation to successional status. Canadian Journal of Forest Research, 1987, 17(8): 829-834. DOI:10.1139/x87-131 |
| [23] |
Fitter A H. An architectural approach to the comparative ecology of plant root systems. New Phytologist, 1987, 106(1): 61-77. |
| [24] |
Fitter A H, Stickland T R. Architectural analysis of plant root systems:Ⅲ. Studies on plants under field conditions. New Phytologist, 1992, 121(2): 243-248. DOI:10.1111/j.1469-8137.1992.tb01110.x |
| [25] |
Bouma T J, Nielsen K L, Van Hal J, Koutstaal B. Root system topology and diameter distribution of species from habitats differing in inundation frequency. Functional Ecology, 2001, 15(3): 360-369. |
| [26] |
Ketipearachchi K W, Tatsumi J. Local fractal dimensions and multifractal analysis of the root system of legumes. Plant Production Science, 2000, 3(3): 289-295. DOI:10.1626/pps.3.289 |
| [27] |
杨小林, 张希明, 李义玲, 解婷婷, 王伟华. 塔克拉玛干沙漠腹地几种植物根系分形特征. 干旱区地理, 2009, 32(2): 249-254. |
| [28] |
赵艳云, 陆兆华, 夏江宝, 刘京涛. 黄河三角洲贝壳堤岛3种优势灌木的根系构型. 生态学报, 2015, 35(6): 1688-1695. |
| [29] |
Meyer K M, Ward D, Wiegand K, Moustakas A. Multi-proxy evidence for competition between savanna woody species. Perspectives in Plant Ecology, Evolution and Systematics, 2008, 10(1): 63-72. DOI:10.1016/j.ppees.2007.09.002 |
| [30] |
陈永乐, 张志山, 吴攀, 回嵘, 虎瑞, 高艳红. 腾格里沙漠东南缘不同年代固沙区根系分布. 中国沙漠, 2013, 33(2): 515-521. |
| [31] |
张道远, 王红玲. 荒漠区几种克隆植物生长构型的初步研究. 干旱区研究, 2005, 22(2): 219-224. |
| [32] |
班芷桦, 王琼. 喜旱莲子草和接骨草竞争对模拟增温的响应. 植物生态学报, 2015, 39(1): 43-51. |
| [33] |
Bauhus J, Khanna P K, Menden N. Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii. Canadian Journal of Forest Research, 2000, 30(12): 1886-1894. DOI:10.1139/x00-141 |
| [34] |
Pan Y X, Wang X P, Zhang Y F, Hu R. Dew formation characteristics at annual and daily scale in xerophyte shrub plantations at Southeast margin of Tengger desert, northern China. Ecohydrology, 2018, 11(5): e1968. DOI:10.1002/eco.1968 |
| [35] |
Nielsen K L, Miller C R, Beck D, Lynch J P. Fractal geometry of root systems:Field observations of contrasting genotypes of common bean (Phaseolus vulgaris L.) grown under different phosphorus regimes. Plant and Soil, 1999, 206(2): 181-190. |