 2020, Vol. 40
2020, Vol. 40文章信息
- 任晴, 袁位高, 吴初平, 王志高, 江波, 朱锦茹, 黄玉洁, 叶明, 方庆
- REN Qing, YUAN Weigao, WU Chuping, WANG Zhigao, JIANG Bo, ZHU Jinru, HUANG Yujie, YE Ming, FANG Qing
- 浙江省红楠生境地群落数量分类和环境解析
- Community quantitative classification and environmental analysis of the Machilus thunbergii habitat in Zhejiang Province
- 生态学报. 2020, 40(15): 5277-5287
- Acta Ecologica Sinica. 2020, 40(15): 5277-5287
- http://dx.doi.org/10.5846/stxb201906031175
-
文章历史
- 收稿日期: 2019-06-03
- 网络出版日期: 2020-05-21

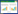




2. 浙江省林业科学研究院, 杭州 310023;
3. 寿昌林场, 建德 311600;
4. 建德林场, 建德 311600
2. Zhejiang Academy of Forestry, Hangzhou 310023, China;
3. Shouchang Forest Farm, Jiande 311600, China;
4. Jiande Forest Farm, Jiande 311600, China
红楠为樟科润楠属的常绿阔叶乔木, 是亚热带和暖温带地区常绿阔叶林的主要建群种或者伴生种, 常散生于山地阔叶混交林中[1-2]。红楠不仅被广泛用作绿化树种和用材树种, 因其十分复杂的树体化学成分, 还多用于香料提取、药品制备等领域[3-5]。但由于红楠天然资源及其生态环境的严重破坏, 使该植物逐渐成为渐危树种[6]。国内对红楠群落的研究主要集中在局部区域的群落调查。黄宗安[7]和曾繁茂[8]调查发现福建省尤溪县境内的红楠种群呈随机分布, 发展稳定。陈子林等[9]发现红楠种群在浙江省六十田自然保护区呈聚集分布状态。廖承川等[10]发现浙江九龙山国家级自然保护区红楠群落以樟科和山茶科植物为主, 并且群落中红楠将长期处于优势地位。但是以上研究仅直观的反映局部区域红楠的生境与分布状况, 并不能归纳出红楠的典型生境和解释环境因子对红楠种群分布及发展的影响。另外红楠林是浙江省典型森林类型, 研究红楠林环境与结构是天然林恢复、珍贵树种等林业工程建设中急需解决的科学问题, 对指导林业生态建设具有重要意义。
植被群落的数量分类和排序是揭示群落与环境关系的重要工具, 双向指示种分析(TWINSPAN)和典范对应分析(CCA)是目前国际上常用的研究方法[11-12]。杨国栋等[13]通过TWINSPAN分类和DCCA排序研究了四川省宝兴县东拉山短丝木犀(Osmanthus serrulatus)植物群落的演替变化趋势;商天其等[14]采用TWINSPAN分类和CCA方法对嵊州市公益林群落进行数量分类与排序, 发现以马尾松(Pinus massoniana)为建群种的群落是该区域主要的优势群落类型;李婷婷等[15]对东北阔叶红松林区的CCA排序结果显示其变化格局主要受温度和降水的影响。目前关于红楠群落的排序研究鲜有报道。
探究群落内部优势种之间的关系, 对揭示群落结构特征及反映群落的稳定程度具有重要意义[16-17]。梁林峰等[18]对山西省文冠果(Xanthoceras sorbifolia)群落优势种的种间关联性进行定量研究, 发现群落整体结构不稳定, 优势物种处于演替变化过程中;俞筱押等[19]对贵州茂兰国家级自然保护区内濒危植物四药门花(Loropetalum subcordatum)群落各层次优势种群的种间联结性进行了分析, 发现四药门花在该群落中表现出较强的独立性;高浩杰等[20]对舟山桃花岛次生林优势种红楠的种内和种间竞争强度进行定量分析, 发现其受到的种内和种间竞争压力均衡。但是红楠与群落物种间的关联关系未见研究。
总体来说, 目前关于红楠群落的研究多集中于局部区域的种群分布和群落结构方面, 而群落类型、群落分布与环境关系、群落内部种间关系等研究较为缺乏, 群落结构、环境因子对红楠分布及生长的影响尚不能解释。本研究从覆盖浙江省11个辖区下属大部分县级行政区的3181个监测样地中, 仅筛选出172个有红楠分布的样地, 说明样地环境、群落结构等对红楠在浙江省的分布及生长有重要影响。因此本文运用双向指示种分类(TWINSPAN)、典范对应分析(CCA)并结合种间联结, 对浙江省172个红楠生境地群落进行分析, 探讨群落类型、环境因子、种间关系对浙江省红楠分布及生长的影响, 以期在省级层次上更为全面的为红楠天然资源的恢复和发展提供科学依据。
1 研究区概况浙江省境内地形起伏较大, 西南、西北部地区群山峻岭, 中部、东南地区以丘陵和盆地为主, 东北地区地势较低, 以平原为主。浙江省属亚热带季风气候, 年平均气温15—18℃, 光照较多, 年平均日照时数1710—2100 h, 降雨丰沛, 年平均雨量980—2000 mm, 空气湿润, 气候资源配制多样。浙江省物种组成丰富, 植被类型主要为常绿阔叶林、针叶林、针阔混交林、落叶阔叶林、常绿落叶阔叶混交林等[1-2]。
2 研究方法 2.1 样地设置与调查2009—2016年间设置的覆盖浙江全省范围的3181个监测样地, 每5年调查1次, 本文采用最近1次(2013—2016年)调查数据, 筛选出有红楠分布的样地共172个(表 1、图 1)。样地面积400 m2(20 m×20 m), 调查内容包括胸径≥5 cm个体的胸径、高度、冠幅等物种数据, 海拔、坡度、坡向、土层厚度等环境数据。气候数据通过样地所在县气象站获取。
| 地区 Region |
含红楠样地数/个 Number of plots containing Machilus thunbergii |
总样地数/个 Total number of plots |
| 富阳 | 1 | 88 |
| 桐庐 | 1 | 145 |
| 柯城 | 1 | 52 |
| 建德 | 2 | 166 |
| 泰顺 | 5 | 160 |
| 景宁 | 44 | 181 |
| 松阳 | 9 | 133 |
| 遂昌 | 26 | 225 |
| 云和 | 9 | 95 |
| 磬安 | 3 | 125 |
| 武义 | 11 | 131 |
| 新昌 | 1 | 55 |
| 德清 | 1 | 31 |
| 龙游 | 3 | 46 |
| 仙居 | 11 | 123 |
| 龙泉 | 16 | 160 |
| 黄岩 | 6 | 95 |
| 临海 | 3 | 150 |
| 定海 | 2 | 58 |
| 普陀 | 17 | 77 |

|
| 图 1 调查样地分布图 Fig. 1 The distribution of plots |
红楠分布的172个样地大部分集中于浙西南丘陵山地区域。样地各环境因子的主要特征如表 2所示, 84.3%的红楠分布在坡度为17°—35°的范围内, 95.35%的红楠分布在海拔1000 m以下的范围内。另外枯落物厚度5 cm以下、腐殖质厚度5—10 cm、土壤厚度30—50 cm的红壤条件下有较多分布。
| 环境因子 Environment factors |
分类 Class |
数量百分比/% Number percentage |
| 坡度/(°) Slope |
17—25 | 34.30 |
| 25—35 | 47.67 | |
| 35—45 | 15.70 | |
| > 45 | 2.33 | |
| 枯落物厚度/cm Litter layer thickness |
<5 | 56.98 |
| 5—10 | 41.28 | |
| >10 | 1.74 | |
| 腐殖质厚度/cm Humus thickness |
<5 | 31.98 |
| 5—10 | 61.05 | |
| >10 | 6.97 | |
| 坡向 Aspect |
阴坡/半阴坡 | 31.98 |
| 阳坡/半阳坡 | 68.02 | |
| 海拔/m Elevation |
<200 | 13.95 |
| 200—500 | 13.95 | |
| 500—1000 | 67.45 | |
| >1000 | 4.65 | |
| 土壤厚度/cm Soil thickness |
<30 | 8.72 |
| 30—50 | 53.49 | |
| >50 | 37.79 | |
| 土壤质地 Soil texture |
壤土 | 68.02 |
| 黏土 | 2.33 | |
| 砂土 | 29.65 | |
| 土壤类型 Soil type |
红壤 | 73.26 |
| 黄壤 | 26.74 |
重要值指标计算在Exccel中完成。计算公式为[21-22]:
相对多度=某一植物种类株数/所有植物种类株数×100%;
相对显著度=某一树种的胸径平方和/所有树种的胸径平方和×100%;
相对频度=某一树种的频度/所有树种的频度×100%;
乔木重要值=(相对显著度+相对频度+相对多度)/3。
2.2.2 双向指示种分析TWINSPAN分类软件采用WinTWINS 2.3, 依据每个样地所有物种重要值数据, 整理成物种编号为横坐标, 样地号为纵坐标的矩阵数据[23]进行载入分析。
2.2.3 典范对应分析CCA采用国际通用软件CANOCO 4.5完成CCA排序。CCA要求两个数据矩阵, 一个是环境数据矩阵, 一个是物种数据矩阵[23-24]。本文选取年均气温、年均降水量、坡度、坡向、海拔高度、土壤类型、土壤质地、土壤厚度、腐殖质厚度、枯落物厚度建立环境因子矩阵, 坡向、土壤类型、土壤质地以数字等级表示, 其他环境指标以实际观测值表示[13-15]。植被数据矩阵为P×N维的物种重要值矩阵, 其中P为植物的种数, N为样地数。
2.2.4 种间联结根据物种在样方内是否出现的二元数据矩阵作为原始数据, 以卡方检验(χ2)为基础, 结合联结系数(AC)和共同出现百分率(PC)对群落优势种进行种间联结分析[17-19]。在计算χ2及联结系数AC时, 为避免a、c、d为0, 导致χ2及AC出现无法比较关联程度的情况, 把a、c、d为0的数值都加权为1。由于本研究的取样方法为非连续性取样, 使用非连续性数据的Yates连续校正公式计算χ2值。
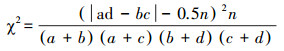
|
式中n为取样总数。当ad>bc时, 两物种为正关联, 存在较弱的竞争;当ad < bc时, 两物种为负关联, 物种之间存在较强竞争。根据χ2查表, 当χ2=0时, 表明两物种间相互独立, 基本上无关联;当P>0.05, 即χ2 < 3.841时, 种间联结独立, 表现出不显著关联性;当0.01≤P≤0.05, 即3.841≤χ2≤6.635时, 种间联结显著, 表现出显著关联性;当P < 0.01, 即χ2>6.635时, 两物种间表现出极显著关联性。
联结系数(AC)计算公式为:
当ad ≥ bc, 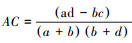
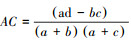
当bc > ad且d < a, 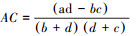
AC的值域为[-1, 1], 其值越接近1, 种对间的正联结性越强;其值越接近-1, 物种间的负联结性越强;其值越接近0, 种对间的联结性越弱;当AC值为0, 种对间完全独立。
共同出现百分率(PC)计算公式为:
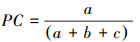
|
PC的值域为[0, 1], 其值越接近1, 种对之间的正联结关系越强;其值越接近0, 该种对的正联结关系越弱。
3 结果与分析 3.1 群落物种组成与重要值172个样地中记录到54科120属167种植物, 其中乔木119种, 灌木或小乔木48种。物种组成以壳斗科、樟科、蔷薇科、桑科、大戟科等科的植物为主(图 2), 其中尤以壳斗科(18种)和樟科(18种)植物最为显著。对整理好的172个样地所有物种数据统一进行重要值计算, 重要值大于1%的共有16个物种(表 3)。

|
| 图 2 物种科属分布图 Fig. 2 Family and Genus Distribution Map |
| 编号 Number |
植物种 Species |
科 Family |
属 Genus |
重要值 Importancevalue/% |
| 1 | 杉木 | 杉科 | 杉木属Cunninghamia lanceolata | 25.49 |
| 2 | 木荷 | 山茶科 | 木荷属Schima superba | 8.86 |
| 3 | 青冈 | 壳斗科 | 青冈属Cyclobalanopsis glauca | 7.64 |
| 4 | 马尾松 | 松科 | 松属Pinus massoniana | 6.17 |
| 5 | 甜槠 | 壳斗科 | 锥属Castanopsis eyrei | 5.47 |
| 6 | 红楠 | 樟科 | 润楠属Machilus thunbergii | 4.54 |
| 7 | 石栎 | 壳斗科 | 石栎属Lithocarpus glaber | 2.30 |
| 8 | 白栎 | 壳斗科 | 栎属Quercus fabri | 1.91 |
| 9 | 枫香 | 金缕梅科 | 枫香树属Liquidambar formosana | 1.89 |
| 10 | 合欢 | 豆科 | 合欢属Albizia julibrissin | 1.56 |
| 11 | 苦槠 | 壳斗科 | 锥属Castanopsis sclerophylla | 1.27 |
| 12 | 榆树 | 榆科 | 榆属Ulmus pumila | 1.13 |
| 13 | 漆树 | 漆树科 | 漆属Toxicodendron vernicifluum | 1.07 |
| 14 | 化香 | 胡桃科 | 化香树属Platycarya strobilacea | 1.02 |
| 15 | 黄山松 | 松科 | 松属Pinus taiwanensis | 1.00 |
| 16 | 赤杨叶 | 安息香科 | 赤杨叶属Alniphyllum fortunei | 1.00 |
根据TWINSPAN的分类结果(图 3), 采取第5级水平的划分, 结合生态学意义, 得到根据不同优势种确定的8种群落类型:

|
| 图 3 浙江省172个红楠生境地群落的双向指示种分类树状图 Fig. 3 Two-way indicator tree classification of 172 Machilus thunbergii communities in Zhejiang Province D:样地分组;N:组内样地数;1—172:样地编号;Ⅰ—Ⅷ:群落编号 |
Ⅰ为杉木+红楠+木荷林, 共计33个样方, 伴生种有青冈、甜槠、苦槠、白栎、枫香、漆树、合欢等。该群落分布于年均气温17.2℃, 年均降水量1673 mm的地区, 坡度22°—38°的阳坡或半阳坡, 海拔高度350—950 m, 平均海拔676.2 m, 土壤为红壤或黄壤。
Ⅱ为杉木林, 共计21个样方, 伴生种有红楠、木荷、甜槠、石栎、赤杨叶、漆树、三角槭(Acer buergerianum)等。该群落分布于年均气温17.3℃, 年均降水量约1593 mm的地区, 坡度25°—44°的半阳坡, 海拔高度300—900 m, 平均海拔616.62 m, 土壤为红壤。
Ⅲ为青冈+甜槠+木荷林, 共计61个样方, 伴生种有红楠、石栎、乌冈栎(Quercus phillyraeoides)、冬青(Ilex chinensis)、杜英(Elaeocarpus decipiens)、赤杨叶、漆树等。该群落分布于年均气温17.2℃, 年均降水量约1622 mm的地区, 坡度25°—47°的半阳坡或半阴坡, 海拔高度400—1300m, 平均海拔731.8m, 土壤为红壤或黄壤。
Ⅳ为马尾松+木荷林, 共计23个样方, 伴生种有青冈、甜槠、苦槠、红楠、黄山松、乌冈栎、枫香、石栎等。该群落样地分布于年均气温17.4℃, 年均降水量1570 mm的地区, 坡度25°—42°的阳坡或半阳坡, 海拔高度100—800 m, 平均海拔440.1 m, 土壤类型为红壤。
Ⅴ为甜槠+红楠+青冈林, 共计8个样方, 伴生树种主要有石栎、香樟(Cinnamomum camphora)、木荷、山矾(Symplocos sumuntia)、三角槭、白栎。该群落分布于年均气温17.1℃, 年均降水量约1544 mm的地区, 坡度25°—45°的阴坡或半阳坡, 海拔高度550—1250 m, 平均海拔831.1 m, 土壤为砂质黄壤。
Ⅵ为青冈+枫香林, 共计6个样方, 伴生种有合欢、白栎、红楠、冬青、化香、黄檀(Dalbergia hupeana)、山矾、木荷等。该群落分布于年均气温16.1℃, 年均降水量约1228 mm的地区, 坡度17°—25°的半阴坡或半阳坡, 海拔高度45—135 m, 平均海拔70.2 m, 土壤为红壤。
Ⅶ为红楠+榆树林, 共计13个样方, 伴生种有香樟、黄檀、化香、野桐(Mallotus japonicus)、臭椿(Ailanthus altissima)、冬青、山矾等。该群落分布于年均气温16.1℃, 年均降水量约1242 mm的地区, 坡度17°—33°的半阴坡或半阳坡, 海拔高度24—220 m, 平均海拔87.1 m, 土壤为红壤。
Ⅷ为红楠+白栎林, 共计7个样方, 伴生种有苦槠、黄檀、化香、盐肤木(Rhus chinensis)、山胡椒(Lindera glauca)等。该群落分布于年均气温16.4℃, 年均降水量约1489 mm的地区, 坡度22°—35°的阴坡或半阳坡, 海拔高度75—800 m, 平均海拔437m, 土壤类型为红壤。
3.3 CCA排序运用Canoco 4.5软件对红楠生境地的环境数据和所有物种数据进行排序。DCA排序结果表明, 所有轴的梯度长度最大为4.214, 应选择单峰模型(CCA)。4个排序轴的特征值分别为0.513、0.377、0.239、0.188, 前3个排序轴特征值占总特征值的85.7%, 包含了绝大部分信息, 表明本研究选取的环境因子对群落分布具有较好的解释意义。
CCA排序(表 4)结果说明环境因子对群落中物种分布具有显著影响(P=0.002)。物种CCA排序轴第一轴特征值为0.274。物种-环境相关性系数为0.759。前四轴物种-环境关系方差累计贡献率为89.8%, 其中前两轴就达到76.4%, 说明CCA前两轴已能较好地反映出研究区群落样地、物种与其环境之间的关系。根据10个环境因子与CCA排序轴的相关性分析(表 5)可知, 海拔高度与CCA第1轴极显著负相关, 与CCA第1轴相关性最强, 相关系数为-0.7608, 其次是坡度, 相关系数为-0.6405, 与CCA第1轴呈显著负相关关系。第二轴与土壤类型的相关系数最高。
| 排序轴Axies | 第一轴 Axies Ⅰ |
第二轴 Axies Ⅱ |
第三轴 Axies Ⅲ |
第四轴 Axies Ⅳ |
| 特征值Eigenvalues | 0.274 | 0.068 | 0.032 | 0.028 |
| 物种-环境相关性Species-environment correlations | 0.759 | 0.443 | 0.396 | 0.311 |
| 物种数据累计百分比方差Cumulative percentage variance of species data | 8.8 | 11.0 | 12.0 | 12.9 |
| 物种-环境因子关系数据累计百分比方差Cumulative percentage variance of species-environment relation | 61.2 | 76.4 | 83.5 | 89.8 |
| 所有典范轴的显著性检验Test of significance of all canonical axes | F-ratio=2.689 P-value=0.0020 | |||
| 环境因子Environment factors | 第一轴 Axies Ⅰ |
第二轴 Axies Ⅱ |
第三轴 Axies Ⅲ |
第四轴 Axies Ⅳ |
| 年平均气温Annual average temperature(AAT) | -0.4596 | -0.0536 | -0.1340 | -0.2654 |
| 年平均降水量Annual average precipitation(AAP) | -0.5099* | 0.4099 | -0.3960 | 0.3709 |
| 坡度Slope(SL) | -0.6405* | 0.1344 | 0.1111 | -0.3469 |
| 坡向Aspect(ASP) | 0.0637 | -0.2288 | -0.1209 | -0.1267 |
| 海拔Elevation(ELE) | -0.7608** | -0.4440 | -0.1753 | 0.0110 |
| 土壤类型Soil type(SOL TYP) | -0.3016 | 0.4855* | 0.3787 | 0.3418 |
| 土壤质地Soil texture(SOL TEX) | 0.0247 | -0.0863 | 0.0853 | -0.5541* |
| 土壤厚度Soil thickness(SOI THIC) | 0.0289 | 0.1259 | -0.5402* | -0.0738 |
| 腐殖质厚度Humus thickness(HUM THIC) | -0.1030 | -0.2927 | -0.4796* | 0.2346 |
| 枯落物厚度Litter layer thickness(LLT) | 0.0966 | -0.2767 | 0.3050 | 0.5522* |
| * P < 0.05, ** P < 0.01 | ||||
通过CanoDRAW分别对样地、优势种与环境因子关系进行分析绘制(图 4、图 5)。由图 4可知,第一轴反映了红楠生境地群落所在生境的海拔、坡度及年平均降水量的情况, 即沿着CCA第一轴从左往右, 海拔、坡度逐渐降低, 光热条件逐渐变好, 年均降水逐渐减少, 海拔为主要影响因子。第二轴基本上表现了群落所在生境土壤类型的变化趋势, 即沿着CCA第二轴从上往下群落物种对红壤的倾向减小, 对黄壤的倾向增加, 但红壤类型下样地数更多, 群落多样性更加显著, 表明浙江省内有红楠分布样地土壤多为红壤类型。图中8个群落类型未能有清晰地界线区分, 存在明显的聚集和交错现象, 这可能是由于样地内物种组成和环境条件的相似性导致的。


|
| 图 5 优势种与环境因子排序图 Fig. 5 CCA ordination diagram of dominant species and environmental factors 1—16为树种编号 |
由图 5可知, 榆树、枫香、合欢分布于第一轴右端, 其生境为坡度平缓、光照条件较好、年均降水量相对较少的低海拔区域;而第一轴的左端并没有物种的倾向性分布, 说明16种优势树种并不适生于高海拔地区, 红楠与白栎第一轴位置接近, 说明二者对海拔、坡度要求相近。土壤类型与第二轴相关性最高, 沿CCA的第二轴由下往上对土壤要求从黄壤向红壤转变。红楠与木荷、杉木、白栎在第二轴位置接近, 说明这些物种对土壤要求相近。
3.4 优势种种间关系种间联结能够更直观地反映种群之间相互关系, 多物种间总体关联性在一定程度上反映了群落的稳定性[14]。
16个优势树种组成的120个种对的χ2检验结果(表 6)显示所有种对间都为正关联关系, 极显著正相关的有16个种对, 显著正关联的有12个种对, 不显著正关联的有92个种对。大多数种对的种间联结并没有达到显著程度, 显著率(包括极显著)为23.3%, 表明种群间联结较为松散。红楠、青冈、甜槠三者互有极显著正关联, 红楠与木荷为显著正关联, 与其他优势种间为不显著正关联。
| 检验方法Test methods | 正联结Positive correlations | 负联结Negative correlations | 无关联None correlations | |||||||
| χ2 | χ2≥6.635 | 3.841 < χ2 < 6.635 | 0 < χ2≤3.841 | χ2≤-6.635 | -6.635 < χ2≤-3.841 | -3.841 < χ2 < 0 | χ2=0 | |||
| 种对数Number of species pairs | 16 | 12 | 92 | 0 | 0 | 0 | 0 | |||
| AC | AC≥0.6 | 0.2≤ AC < 0.6 | 0.05≤ AC < 0.2 | AC < -0.6 | -0.6≤ AC < -0.2 | -0.2≤ AC < -0.05 | -0.05≤ AC < 0.05 | |||
| 种对数Number of species pairs | 1 | 13 | 25 | 4 | 17 | 13 | 47 | |||
| PC | PC≥0.6 | 0.4≤ PC < 0.6 | 0.2≤ PC < 0.4 | - | - | - | 0≤ PC < 0.2 | |||
| 种对数Number of species pairs | 5 | 5 | 50 | - | - | - | 60 | |||
联结系数AC结果(表 6、图 6)中, 共39个正联结种对, 34个负联结种对, 正负关联比为1.1:1, 无关联种对有47个。红楠与甜槠的AC值结果为极显著正关联, 与木荷、青冈为显著正关联, 与杉木为不显著正关联, 与黄山松为不显著负关联;黄山松-合欢、榆树-苦槠、化香-苦槠、赤杨叶-白栎的AC值结果为极显著负关联。

|
| 图 6 优势种种间关联的AC半矩阵图 Fig. 6 Semi-matrix graph of AC correlation coefficients of dominant species 1—16为树种编号 |
共同出现率PC结果(表 6、图 7)中显著率(包括极显著)为8.3%, 不显著正关联的有50个种对。无关联的有60个种对, 占50%。由此可见, 主要优势种间的正联结程度不高。红楠-杉木、红楠-青冈、红楠-木荷、杉木-木荷、木荷-青冈显示为极显著正关联。显著正关联的种对分别是红楠-甜槠、杉木-甜槠、青冈-甜槠、木荷-甜槠、杉木-青冈。

|
| 图 7 优势种种间关联的PC半矩阵图 Fig. 7 Semi-matrix graph of PC correlation coefficients of dominant species 1—16为树种编号 |
本研究利用TWINSPAN等级分类, 将浙江省内172个红楠生境地群落划分为8个类型, 分别归属于浙江省四种典型的植被类型:针阔混交林、杉木林、常绿阔叶林、常绿落叶阔叶混交林。从演替角度来看, 不同群落在环境因子影响下可能会向特定的方向演替发展。因此, 环境状况不同的区域可以指示不同的植被群落类型[15]。而随着演替进展导致的群落内部环境改变(如光照)也有可能是影响红楠分布及发展的主要原因[25]。
针阔混交林包括杉木、红楠、木荷同为共建种的群落Ⅰ和马尾松、木荷为共建种的群落Ⅳ, 分布在坡度42°以下、海拔1000 m以下的低山区域。速生强阳性树种杉木、马尾松和喜光树种木荷占据林冠层, 与同层弱阳性树种红楠对光照资源需求互补。群落Ⅰ优势种种间联结紧密, 红楠、杉木、木荷、青冈、甜槠之间均有显著关联, 红楠在群落中为优势种, 处于林分上层, 群落组成较稳定。群落Ⅳ中红楠与木荷、青冈、甜槠有显著正关联, 与马尾松无显著关联, 与黄山松为不显著负关联, 群落稳定性有待提高。两类群落分别由杉木林和马尾松林演替而来, 正处于向常绿阔叶林发展的过渡阶段, 群落物种组成丰富, 已储存了常绿阔叶林的大部分物种, 种群结构稳定, 更新良好。
群落Ⅱ属于杉木林, 样地集中于坡度37°以下、海拔900 m以下的低山区域。这一群落以杉木为主, 物种丰富度低, 仍属于演替初级阶段, 林分整体质量较差, 通过调整杉木密度, 加快下层树种进入乔木层, 进行杉木林的阔叶化改造, 可以提高群落稳定性。随演替进展, 耐阴树种红楠优势度会逐渐增大, 此时应注意红楠种群密度并疏伐上层高大乔木, 增加林窗或林缘, 增强红楠与林下阳性植物的竞争力, 提高红楠种群的更新能力。
常绿阔叶林包括以青冈、甜槠、木荷为建群种的群落Ⅲ和以甜槠、红楠、青冈为建群种的群落Ⅴ, 多分布在坡度39°以下、海拔1300 m以下的中低山区域。红楠处于林分中上层, 与优势种青冈、甜槠、木荷均有显著正关联。两类群落乔木层树种主要集中在木荷属、栲属、栎属, 仅有少量的白栎、赤杨叶等落叶树种间杂在林分上层。虽然林分中下层多样性较丰富, 但由于常绿阔叶林郁闭度较高, 林下光照较弱, 容易出现林分断层现象, 应该适当疏伐上层高大乔木, 促进群落结构优化, 保证红楠幼苗的补充连续性。
阔叶混交林包括青冈、枫香为共建种的群落Ⅵ, 红楠、榆树同为建群种的群落Ⅶ, 红楠、白栎同为建群种的群落Ⅷ。群落Ⅵ和群落Ⅶ均分布于浙东南沿海岛屿坡度较缓、温暖避风的低海拔区域, 群落Ⅷ分布于坡度较缓的低山区域。物种组成上, 由于常年受海风胁迫, 相对于内陆地区海岛植被物种多样性较低, 主要物种与大陆相似, 以壳斗科、山茶科占优势。这三类群落处于阔叶混交林幼龄阶段, 乔木层树种资源较少, 但灌木层成分较多, 红楠作为防风树种, 能够在多样性相对较低的海岛植被中占据林分上层优势地位, 增长潜力大, 但应注意控制种群密度, 防止种内竞争。
从环境因子来看, 红楠生存群落的分布受到多种环境因子如海拔、坡度、降水的影响, 主导因子是海拔高度。红楠的生存群落在坡度较缓的低山区域有较明显倾向性分布, 体现了水热条件对于红楠在浙江省分布及发展的重要性, 这与张志敏等[3]、胡绍庆[4]、江香梅和俞湘[6]、徐振东等[26]对红楠分布环境的描述相符。在温度及降水条件基本相同的地域, 林下微环境决定了种群的更新和生长状况[27]。腐殖质厚度反映了土壤肥力的不同, 枯落物从一定程度上影响了地表温度、含水量[28-29]以及植被幼苗的萌发, 厚而致密的枯落物阻碍了种子着陆也影响了光向落叶层下的传播[27]。因此红楠在相对腐殖质更厚、枯落物更薄的群落Ⅴ、群落Ⅶ中的生长明显比在群落Ⅲ、群落Ⅵ生长更占优势, 表明了土壤肥力、林下光照对于红楠幼苗更新生长的重要性。充足的光照能够提高红楠幼苗的存活率, 因此相比于郁闭度较大的群落Ⅳ, 红楠在林窗较多的群落Ⅰ中优势更为明显。
从种间关系来看, 一定空间范围内的不同植物个体之间存在相互作用。甜槠、木荷、青冈是红楠生存群落中出现频率最高、优势地位最为显著的树种, 同时也是浙江省森林群落中十分稳定的优势种[30-32]。红楠与青冈、木荷、甜槠等群落优势种具有较紧密的联结关系, 再加上相对稳定的自然环境和低干扰性能够增强群落结构的稳定性, 因此红楠能够在这几类优势种群落中占据较为有利的优势地位。但红楠、杉木、木荷、白栎对海拔、坡度、降水等环境因子的要求相近, 因此对资源的抢夺也可能会导致种间竞争, 影响红楠的生长与更新。而且联结结果显示大多数种对种间联结程度不显著或者无关联, 表明红楠群落整体结构不够稳定。因此在群落恢复和发展过程中, 选择配置联结性高的树种以增强群落稳定性的同时, 要注意环境资源的均衡分配。另外在环境条件基本一致的植被群落中, 邻株植物功能类型对红楠的生长也有显著影响, 邻株植物为落叶阔叶型物种时会促进红楠的生长[33], 这可能是红楠能够在以榆树、白栎为优势种的群落中占据优势地位的重要原因。
本研究还存在一定的局限性。研究中仅系统讨论了红楠生境地群落类型, 下一步仍需对红楠优势林群落和伴生林群落进行比较分析, 以探究影响红楠发展的因素。此外由于本研究仅针对浙江省红楠生境样地, 忽略了其他类型样地中制约红楠分布的生物及非生物因素。在以后的研究中, 可以对浙江省不同类型样地统一考虑, 再根据结果对分布红楠的主要类型区进行环境解析。全面分析的结果也可以和单独分析红楠生境地群落的结果进行比对, 探究是否忽略了其他影响因素。
5 结论(1) 通过TWINSPAN将172个红楠生境地群落划分成8个群落类型。由于群落类型不同, 随着演替而导致的群落内部环境改变(如光照)也有可能是影响红楠分布及发展的主要原因。在不同的演替阶段, 通过人工疏伐或补植等措施, 能够为红楠创造更加有利的生长环境, 提高红楠种群的更新能力。
(2) 海拔高度及坡度是决定红楠分布及发展的主导环境因子。红楠作为优势种的生境地主要特征为海拔900m以下、坡度35°以下。在水热条件基本相同的地域, 郁闭度、枯落物厚度等决定的林下微环境决定了种群的更新和生长状况, 因此红楠的分布也要考虑光照、土壤等其他环境因素的影响。
(3) 青冈、木荷、甜槠均为构成浙江省典型森林群落的稳定优势种。种间联结初步分析显示, 红楠与青冈、木荷、甜槠等群落优势种具有显著正关联。由以上树种构成的群落, 加上相对稳定的自然环境和低干扰性, 能够增强结构稳定性, 但应注意环境资源的均衡分配, 减少种间竞争。
基于物种重要值的TWINSPAN群落分类法结合CCA排序法, 对浙江省红楠生境地群落取得了良好的分类结果, 归纳出浙江省红楠生境地的主要群落类型和环境特征, 解释了环境因子和种间关系对红楠分布及发展的影响, 同时从群落演替的角度提出促进红楠生长及发展的对策, 可为浙江省红楠天然资源的恢复和人工林的经营提供数据支撑。
| [1] |
王希华.中国典型常绿阔叶林植物地理与物种多样性研究[D].上海: 华东师范大学, 2006. http://cdmd.cnki.com.cn/Article/CDMD-10269-2007016276.htm
|
| [2] |
袁位高.浙江省生态公益林主要群落结构的比较研究[D].北京: 中国林业科学研究院, 2009. http://cdmd.cnki.com.cn/article/cdmd-82201-2009194266.htm
|
| [3] |
张志敏, 杨国栋, 谢梦梦, 钟育谦, 翟飞飞, 王贤荣, 伊贤贵. 江苏龙池山自然保护区红楠(Machilus thunbergii)生存群落结构及其环境解释. 生态学杂志, 2019, 38(6): 1637-1645. |
| [4] |
胡绍庆. 园林绿化新秀——红楠. 浙江林业, 2002(12): 29-29. |
| [5] |
马立宇, 黄剑坚. 乡土树种红楠的研究进展及开发应用前景. 防护林科技, 2019(6): 66-68. |
| [6] |
江香梅, 俞湘. 红楠及其研究进展. 江西农业大学学报, 2001, 23(2): 231-235. |
| [7] |
黄宗安. 红楠群落主要种群分布格局的研究. 福建林学院学报, 1996, 16(1): 49-52. |
| [8] |
曾繁茂. 红楠群落特征的初步研究. 福建林业科技, 1999, 26(1): 42-45. |
| [9] |
陈子林, 陈煜, 周钰鸿. 浙江六十田自然保护小区红楠种群结构与更新. 林业科技开发, 2007, 21(4): 33-36. |
| [10] |
廖承川, 李成惠, 陈卫新, 张家银, 巫松标, 廖进平. 浙江九龙山自然保护区红楠群落特征及种群动态的研究. 福建林业科技, 2007, 34(4): 129-133. |
| [11] |
田永强, 俞超超, 王磊, 黄邦钦. 福建九龙江北溪浮游植物群落分布特征及其影响因子. 应用生态学报, 2012, 23(9): 2559-2565. |
| [12] |
赵从举, 康慕谊, 雷加强. 准噶尔盆地典型地段植物群落及其与环境因子的关系. 生态学报, 2011, 31(10): 2669-2677. |
| [13] |
杨国栋, 钱慧蓉, 陈林, 王贤荣. 中国特有植物短丝木犀(Osmanthus serrulatus Rehd.)群落结构及其环境解释. 生态学报, 2018, 38(9): 3059-3068. |
| [14] |
商天其, 郑超超, 高洪娣, 叶诺楠, 伊力塔. 嵊州市公益林群落数量分类、排序及环境解析. 热带亚热带植物学报, 2015, 23(3): 334-342. |
| [15] |
李婷婷, 姬兰柱, 于大炮, 周莉, 周旺明, 毛沂新, 代力民. 东北阔叶红松林群落分类、排序及物种多样性比较. 生态学报, 2019, 39(2): 620-628. |
| [16] |
庾晓红, 李贤伟, 白降丽. 我国植被数量分析方法的研究概况和发展趋势. 生态学杂志, 2005, 24(4): 448-451. |
| [17] |
张明霞, 王得祥, 康冰, 张岗岗, 刘璞, 杜焰玲, 于飞. 秦岭华山松天然次生林优势种群的种间联结性. 林业科学, 2015, 51(1): 12-21. |
| [18] |
梁林峰, 裴淑兰, 张钦弟, 毕润成, 闫明. 山西文冠果群落优势物种种间关联性分析. 林业资源管理, 2018(4): 76-83. |
| [19] |
俞筱押, 余瑞, 黄娟, 袁子勇, 杨芳, 韩国栋. 贵州茂兰国家级自然保护区四药门花群落各层次优势种群的种间联结性分析. 植物资源与环境学报, 2018, 27(2): 24-32. |
| [20] |
高浩杰, 高平仕, 王国明. 舟山群岛红楠林种内和种间竞争研究. 植物研究, 2017, 37(3): 440-446. |
| [21] |
白晓航, 张金屯. 小五台山森林群落优势种的生态位分析. 应用生态学报, 2017, 28(12): 3815-3826. |
| [22] |
张金屯. 历山自然保护区森林群落的典范主分量分析. 生物数学学报, 2005, 20(2): 213-218. |
| [23] |
巨天珍, 王彦, 任海峰, 姚晶晶, 王蒙, 孟凡涛, 王继伟. 小陇山国家级自然保护区次生林分类、排序及演替. 生态学杂志, 2012, 31(1): 23-29. |
| [24] |
苏日古嘎, 张金屯, 张斌, 程佳佳, 田世广, 张钦弟, 刘素军. 松山自然保护区森林群落的数量分类和排序. 生态学报, 2010, 30(10): 2621-2629. |
| [25] |
王世雄, 王孝安, 李国庆, 郭华, 朱志红. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释. 生态学报, 2010, 30(6): 1638-1647. |
| [26] |
徐振东, 杨曼, 胡蝶, 丁琼, 费永俊. 润楠属主要树种的资源状况、研究现状和开发利用前景. 长江大学学报:自然科学版, 2016, 13(3): 13-17. |
| [27] |
徐丹.红楠种群结构及动物对其种子的传播[D].南京: 南京林业大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10298-1016702205.htm
|
| [28] |
童笑笑, 陈春娣, 吴胜军, 贾振毅, 易雪梅, 马茂华. 三峡库区澎溪河消落带植物群落分布格局及生境影响. 生态学报, 2018, 38(2): 571-580. |
| [29] |
丁佳, 吴茜, 闫慧, 张守仁. 地形和土壤特性对亚热带常绿阔叶林内植物功能性状的影响. 生物多样性, 2011, 19(2): 158-167. |
| [30] |
祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构. 植物生态学报, 2008, 32(2): 262-273. |
| [31] |
翁东明, 郭瑞, 张宏伟, 程樟峰, 田云, 余晶晶, 王月霞, 刘金亮. 浙江省自然保护区落叶阔叶林群落物种组成与结构比较研究. 浙江大学学报:理学版, 2014, 41(5): 600-610. |
| [32] |
孙杰杰, 沈爱华, 黄玉洁, 袁位高, 吴初平, 叶诺楠, 朱锦茹, 邱浩杰, 焦洁洁, 江波. 浙江省大叶榉树生境地群落数量分类与排序. 南京林业大学学报:自然科学版, 2019, 43(4): 85-93. |
| [33] |
童跃伟, 项文化, 王正文, Durka W, Fischer M. 地形、邻株植物及自身大小对红楠幼树生长与存活的影响. 生物多样性, 2013, 21(3): 269-277. |