 2020, Vol. 40
2020, Vol. 40文章信息
- 赵倩, 任广伟, 王杰, 王新伟, 韦建玉, 王晓强, 卢燕回, 陈信, 王静
- ZHAO Qian, REN Guangwei, WANG Jie, WANG Xinwei, WEI Jianyu, WANG Xiaoqiang, LU Yanhui, CHEN Xin, WANG Jing
- 施用韩国假单胞菌(Pseudomonas koreensis)CLP-7对连作烟田土壤质量及微生物群落功能多样性的影响
- Effects of adding Pseudomonas koreensis CLP-7 on soil quality and soil microbial community functional diversity in continuous cropping tobacco fields
- 生态学报. 2020, 40(15): 5357-5366
- Acta Ecologica Sinica. 2020, 40(15): 5357-5366
- http://dx.doi.org/10.5846/stxb201905201037
-
文章历史
- 收稿日期: 2019-05-20
- 网络出版日期: 2020-05-22





2. 广西中烟工业有限公司, 南宁 530000;
3. 广西壮族自治区烟草公司, 玉林 537000
2. Guangxi China Tobacco Industry Co., Ltd., Nanning 530000, China;
3. Tobacco Company of Guangxi Zhuang Autonomous Region, Yulin 537000, China
烟草栽培需要特定的地理和气候条件, 受土地资源和水利条件制约, 连作障碍严重[1]。连续种植导致土壤pH、化学性质以及细菌丰度等显著下降, 土壤微生物多样性各指标均随连作时间的延长而下降, 而某些土传根部病害, 如烟草青枯病(Tobacco bacterial wilt)病原青枯劳尔氏菌(Ralstonia solanacearum)在连作过程中积累, 其数量与连作时间呈显著正相关[2], 且R. solanacearum具有嗜酸性[3], 因此酸化植烟土壤中青枯病发生愈发严重。
土壤微生物多样性, 是指群落中的微生物种群类型和数量、种的丰度和均度以及种的分布情况[4], 一直是生物学研究热点之一。土壤微生物与土壤pH调节和植物病害发生密切相关。首先, 土壤pH值是土壤细菌群落变化的最佳预测因子, 细菌相对丰度和多样性受土壤pH强烈影响[5], 已有研究表明, 土壤酸化可引起青枯病的爆发, 酸性条件(pH 4.5—5.5)有利于病原菌的生长[6];其次, 土传病害的发生与根际土壤微生物的数量关系密切, 土壤微生物群体可在一定程度上抑制作物的土传病害, Li等[7]通过“采用变性凝胶梯度电泳图谱分析微生物群落多样性和丰富度”研究, 表明细菌群落多样性与青枯病抑制性呈负相关, 也就是说, 微生物群落结构越丰富, 多样性越高, 对抗病原菌的综合能力越强。再次, 增加土壤微生物功能多样性可以创造有益于植物生长的条件[8], 而微生物功能多样性可以通过BIOLOG自动微生物鉴定技术分析。BIOLOG-ECO微平板是一种基于生物和生化特性的新方法, 通过描述混合微生物群体水平的生理轮廓及群落功能多样性, 能够快速地表征环境样品的生态状况, 以反映微生物种群总体活性, 表征其生理特征[9]。邹春娇等[10]通过BIOLOG-ECO技术阐明了黄瓜连作营养基质中微生物群落结构多样性特征:采用营养基质栽培黄瓜, 连作第5茬黄瓜产量、微生物碳代谢能力、微生物多样性各项指标及对单一碳源的高利用碳源数均显著高于各茬次;在连作第11茬后微生物碳代谢能力显著下降, 微生物多样性水平显著降低, 微生物对羧酸类碳源利用率明显提高, 微生物群落结构发生单一化现象。表征微生物活性的AWCD值和微生物功能多样性指数虽然能反映土壤微生物的总体变化情况, 并不能反映微生物群落代谢的详细信息, 但是微生物对不同碳源的利用可以反映微生物的代谢功能类群[11]。本研究通过BIOLOG-ECO技术研究施用CLP-7后根际土壤微生物功能代谢多样性变化, 将土壤微生物碳源代谢利用情况与土壤中实际可利用碳源情况相联系, 根据碳源利用率了解根际微生物群落功能多样性。目前, 我国主产烟区土壤酸化趋势明显, 且酸化土壤加重了烟草青枯病的发生和为害, 针对有益微生物改善酸性植烟土壤质量和土壤微生物群落功能多样性的影响研究甚少。专利菌株P. koreensis CLP-7(ZL.201710081711.X)[12]是本实验室自主分离的嗜酸性防病促生菌(PGPR, Plant Growth Promotion Rhizobacteria), 对R. solanacearum具有较强的拮抗活性, 该菌株在pH5.0—5.5条件下生物活性最强, 包括分泌蛋白酶、葡聚糖酶和解钾能力, 适用于酸性土壤条件下烟草真菌和细菌病害的生物防治(专利已授权)。本试验在大田条件下将嗜酸性P. koreensis CLP-7施入连作酸化植烟土壤中, 以施菌后不同时间的根际土壤为研究对象, 主要通过BIOLOG-ECO技术和主成分分析方法研究P. koreensis CLP-7对烟草根际土壤pH、养分含量、酶活活性以及微生物群落功能多样性的影响, 可以为合理施用嗜酸性P. koreensis CLP-7菌株、改善酸性土壤质量和烟草青枯病绿色防控提供生态学方面参考。
1 材料与方法 1.1 试验材料供试:CLP-7液体发酵液, 有效浓度1×1011 CFU /mL。
供试烟草:烤烟品种云烟87。
供试培养基:营养琼脂培养基(NA, Nutrient Agar)、营养肉汤培养基(NB, Nutrient Broth)、添加1%溴麝香草酚蓝水溶液的NA培养基[13]。
1.2 P. koreensis CLP-7施用方法试验地点与时间:本试验于2016年5—7月在湖南省张家界市永定区三家馆岩水井(28°52′—29°48′N, 109°40′—111°20′E)连作烟田进行, 供试烟田(100 m×80 m, 每行240株, 共72行)平分为2个处理区, 处理1是CLP-7, 处理2是空白对照(CK, Check), 每处理3次重复, 每重复2行, 共400株。以云烟87为供试材料(生育期为出苗后45 d), 初移栽后将CLP-7发酵液500倍稀释后灌根做定根水, 每株用量为100 mL, 施用等量灌溉水还苗后(间隔10 d), 再等量施用1次, 共施用2次;CK处理同于上述施菌方法, 施用等量灌溉水, 试验田间管理同于大田标准管理。
1.3 根际土壤采集及处理分别于第二次施菌后10 d(团棵期)、30 d(旺长期)、50 d(成熟期)采用5点取样法, 如图 1所示, 先确定处理小区对角线的中点作为中心取样点, 再在对角线上选择4个与中心样点距离相等的点为取样点, 处理组烟草, 每点5棵烟株, 共25株。根际土样收集采用抖根法[14], 即拔出根系, 去除根围土, 将离主根和须根根轴表面数毫米范围之内附着的土壤抖到自封袋中, 混合均匀共约100 g;另第50天同时采集CK处理土样, 采集方法同上, 取好的土样尽快置于-20 ℃保存、备用。

|
| 图 1 五点取样法取样示意图 Fig. 1 Five-point sampling |
(1) 室内培养基pH测定:通过添加酸碱指示剂——溴麝香草酚蓝测定平板pH变化。在添加1%溴麝香草酚蓝水溶液的NA平板上划线接种CLP-7, 于28 ℃培养箱中黑暗培养48 h。
(2) 烟田土壤pH测定:采用电位法[15]。称取各处理(通过2 mm筛孔)风干土样5 g于50 mL烧杯中, 加入12.5 mL无二氧化碳的水, 搅拌5 min, 静置30 min, 使用PHS-3E型pH计测定土壤pH, 3次重复。
1.4.2 根际土壤酶活活性与养分含量测定土壤酶活测定均参照关松荫[16]方法, 包括土壤脲酶(S-UE, soil urease)、蔗糖酶(S-IA, soil invertase)和过氧化氢酶(S-CAT, soil catalase);土壤养分含量测定均参照鲍士旦[17]方法:速效钾(AK, Available K)用火焰光度计法测定、速效磷(AP, Available P)用钼蓝比色法测定、铵态氮(AN, Ammonium N)用靛酚蓝比色法测定、硝态氮(NN, Nitrate N)用酚二磺酸比色法测定和有机质(OM, Organic matter)用重铬酸钾容量法(外加热法)测定。试验均为3次重复。
1.4.3 BIOLOG-ECO分析根际土壤微生物群落功能多样性BIOLOG-ECO含有31种碳源, 分为氨基酸类、糖类、羧酸类、胺类、双亲化合物及聚合物六类物质[18]。ECO接种液制备方法:将土样放在25 ℃条件下活化24 h, 取5 g(当量干重)土样于100 mL三角瓶中, 加入45 mL灭菌水, 30 min后, 用移液枪取1 mL泥浆到2 mL离心管中。在10000 r/min下离心20 min, 弃去上清液, 加1 mL生理盐水, 在振荡器上振动5 min使之混匀;再于10000 r/min下离心20 min, 重复2次, 除去其中的碳源;弃去上清液, 加1 mL生理盐水, 在振荡器上振动5 min使之混匀, 于2000 r/min下离心1 min。取上清液倒人装有20 mL已灭菌生理盐水(NaC1, 0.85%)的离心管中, 并使其OD590 nm维持在0.13±0.02, 制备出接种液并移至储液槽。使用8孔道排枪向ECO板培养基孔中加150 μL接种液, 每个样品3次重复。接种好的微孔板用自封袋封好, 作好标记后放入28 ℃恒温培养箱中。分别于24、48、72、96、120、144、168、192、216、240 h使用BIOLOG微孔板读数仪自动测定吸光度。
1.5 数据处理(1) 孔的平均颜色变化率:微平板孔中溶液吸光值平均颜色变化率[19](average well color development, AWCD)AWCD=∑(Ci-R)/n。式中, Ci为第i个非对照孔的吸光值, R为对照孔的吸光值, n为培养基碳源种类数(本研究中n=31)。
(2) Shannon-Wiener多样性指数[10]
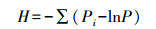
|
式中Pi表示第i个非对照孔中的吸光值与所有非对照孔吸光值总和的比值, 即Pi=(Ci-R)/∑(Ci-R)。
(3) Simpson优势度指数[10]
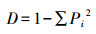
|
(4) Pielou均匀度指数[10]
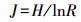
|
式中, R为被利用的碳源总数。
(5) McIntosh指数[10]是基于群落物种多维空间上的Euclidian距离的多样性指数, 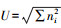
(6) 丰富度指数(R)[10], 指被利用的碳源总数目, 本研究中为每孔中(Ci-R)的值大于0.25的孔数。
主成分分析应用SPSS 19.0软件的Data Reduction工具, 多样性指数、碳源相对利用率, 采用168 h各施菌时间样AWCD值计算[9], 绘图应用Origin 9.0软件。
2 结果与分析 2.1 添加P. koreensis CLP-7对培养基和根际土壤pH的影响首先, CLP-7培养48 h后, 添加指示剂的培养基由初始的黄色(pH 6.0)变为绿色(>pH 7.0)(图 2)说明该菌株的生长代谢过程能引致培养基pH明显升高。其次, 田间试验(3次重复)结果显示(图 3), 随着施用CLP-7时间的增加, 根际土壤pH发生较明显的变化, 第一次采样(团棵期, 施菌10 d)的根际土壤pH是5.1±0.0, 处理间差异显著(P < 0.05);第二次采样(旺长期, 施菌30 d)的根际土壤pH是5.3±0.1;第三次采样(成熟期, 施菌50 d)的根际土壤pH升高至5.4±0.0, CK处理的pH是5.2±0.1, 各处理与对照组差异显著(P < 0.05)。说明施菌后, 土壤pH呈升高趋势。

|
| 图 2 CLP-7划线前后添加指示剂培养基的显色变化 Fig. 2 Color change of indicator amended medium before and after innoculating CLP-7 by plate scribing method |

|
| 图 3 CLP-7不同处理时间烟草根际土壤pH变化 Fig. 3 Change of soil pH in tobacco rhizosphere soil under different time treatments by CLP-7 不同字母代表差异显著(P<0.05) |
如表 1所示, CLP-7处理对根际土壤酶活性存在不同程度的影响。施菌后10 d、30 d和50 d时, 脲酶、蔗糖酶和过氧化氢酶活性均呈先上升后下降的趋势, 其中, 蔗糖酶、脲酶活性高于CK, 30 d时活性最高, 分别比CK增加5.8%和35.5%, 处理间差异显著;过氧化氢酶活性在整个取样时间内均低于CK。
| 处理 Treatment |
蔗糖酶 Invertase/(mg/g±/d) |
过氧化氢酶 Catalase/(mL/g±/h) |
脲酶 Urease/(mg/g±/d) |
| 10 d | 3.3±0.1c | 0.1±0.0c | 28.6±0.1d |
| 30 d | 6.0±0.1a | 0.4±0.1b | 37.5±0.3a |
| 50 d | 3.3±0.0c | 0.3±0.0b | 32.6±0.4b |
| CK | 5.7±0.2b | 1.2±0.2a | 27.6±0.3e |
| 同列数据后不同字母代表差异显著(P<0.05) | |||
由表 2数据可知, P. koreensis CLP-7可以不同程度提高土壤养分含量。铵态氮和硝态氮含量在施菌后10 d最高, 分别比CK增长34.9%和36.3%;然后随施菌时间增加而下降, 处理间差异显著;速效钾含量随施菌时间增加呈先上升后下降趋势, 但3个处理的含量均高于CK, 处理间差异显著;且30 d时, 速效钾含量最高, 比CK增加13.4%;CLP-7对根际土壤有效磷含量影响较大, 施菌10 d时, 根际土壤有效磷含量最高, 比CK增加394.3%, 然后逐渐下降, 至50 d时, 有效磷含量仍比CK增加110.0%, 处理间差异显著;根际土壤有机质含量增加明显, 所有取样时间的土壤有机质含量呈先上升后下降的趋势, 且均高于CK处理, 分别增加了11.1%, 19.0 %和16.7%, 30 d时含量最高。
| 处理 Treatment |
速效钾 Available K/(mg/kg) |
有效磷 Available P/(mg/kg) |
铵态氮 Ammonium N/(mg/kg) |
硝态氮 Nitrate N/(mg/kg) |
有机质 Organic matter/(g/kg) |
| 10d | 393.5±0.9c | 456.5±0.5a | 118.8±0.4a | 50.7±0.8a | 28.9±0.4c |
| 30d | 434.3±0.8a | 235.5±0.4b | 93.5±0.7b | 21.5±0.6b | 36.8±0.5a |
| 50d | 413.8±0.2b | 172.1±1.8c | 65.6±0.6d | 11.1±0.4e | 34.5±1.2b |
| CK | 383.1±1.6d | 62.1±0.6f | 83.9±0.2c | 14.4±0.7c | 17.8±1.0d |
| 同列数据后不同字母代表差异显著(P<0.05) | |||||
由图 4可以看出, 随着培养时间的增加, 根际土壤中的微生物活性不断升高, 自24 h起AWCD值迅速升高, 168 h后趋于稳定状态。对比不同施菌时间根际土壤微生物AWCD值发现, 其变化趋势为30 d>50 d>CK>10 d, 即施菌30 d时, AWCD值达到最高值1.3, 比CK增加371.6%;施菌后50 d时, AWCD值下降为0.67, 但仍比CK增加1.23%, 施菌10 d时的AWCD值最小, 为0.19, 比CK减少0.38%。上述结果说明, CLP-7能显著提高根际土壤微生物对碳源的利用率。

|
| 图 4 CLP-7不同处理时间烟草根际土壤微生物的AWCD值变化 Fig. 4 AWCD variation under different time treatments by CLP-7 in tobacco rhizosphere soil microbes |
对不同处理根际土壤培养168 h后各碳源AWCD值进行主成分分析, 在31种因子中提取3个主成分因子, 第1、2、3主成分得分贡献率分别为49.0%、36.9%、11.3%, 累计贡献率达97.3%, 其余各主成分的贡献率很小, 因此将前3个主成分得分作图, 来表征施菌后不同时间根际土壤微生物碳源代谢特征。结果如图 5所示, 10 d在PC1、PC2上得分最高, 说明施菌10 d烟株土壤微生物碳源利用影响最大;反之, 30 d和CK得分相近且较小, 施菌30 d后与CK处理土壤微生物对碳源利用能力相似程度较高。

|
| 图 5 CLP-7不同处理时间烟草根际土壤微生物碳源代谢主成分分析 Fig. 5 Principal component analysis (PCA) of carbon sources metabolism of tobacco rhizosphere soil microbes by CLP-7 under different time treatments A—D:10 d, 30 d, 50 d, CK |
施用CLP-7后, 根际土壤微生物Shannon-wiener多样性指数、Simpson优势度指数、Pielou均匀度指数、Richness Index丰富度指数以及Mclntosh指数在不同取样时间内发生了不同程度的变化, 从表 3可以看出, Shannon-wiener指数、Simpson指数、Richness指数和Mclntosh指数随着施菌时间的增加呈升高趋势, 施菌30 d时达到最高, 分别为3.3、1.0、26.7和8.0, 比CK分别增长16.5%、3.2%、53.7%和27.5%, 处理间差异显著;而Pielou指数随着施菌时间的增加变化不明显, 与CK基本持平。
| 处理 Treatment |
香浓指数(H) Shannon-wiener Index |
优势度指数(D) Simpson Index |
丰富度指数(R) Richness Index |
均匀度指数(J) Pielou Index |
Mclntosh指数(U) Mclntosh Index |
| 10 d | 2.3±0.3c | 0.4±0.3b | 6.3±1.2d | 1.3±0.3a | 2.67±0.41c |
| 30 d | 3.3±0.0a | 1.0±0.0a | 27.7±1.2a | 1.0±0.0b | 8.02±0.41a |
| 50 d | 2.5±0.1c | 0.9±0.0a | 10.0±1.0c | 1.1±0.0ab | 2.92±0.24c |
| CK | 2.8±0.0b | 0.9±0.0a | 8.0±1.0d | 1.0±0.0b | 2.90±0.68b |
| 同列数据后字母不同代表差异显著(P<0.05) | |||||
BIOLOG ECO微孔板含有31种碳源, 基于碳源官能团的不同, 将其分为6类, 分别是氨基酸类、羧酸类、胺类、聚合物类、双亲化合物类和糖类。本试验各处理土壤微生物群落对6类碳源的相对利用率如图 6所示, CLP-7对根际土壤微生物碳源利用有不同程度的影响。随着施菌时间的增加, 根际土壤微生物对碳源的利用率逐渐增强, 尤其是对羧酸类和糖类碳源的利用率较高, 均超过50%, 但利用率也存在明显差异, 对羧酸和糖类碳源利用率强弱顺序分别为10 d>50 d>CK>30 d和50 d>10 d>30 d>CK。施菌10 d时, 土壤微生物对土壤中羧酸类碳源相对利用率达到100%;施菌50 d, 土壤微生物对糖类、胺类和聚合物类碳源相对利用率均达到100%, 氨基酸类和双亲类碳源相对利用率在施菌后30 d达到最高, 分别为70.3%和100%。

|
| 图 6 CLP-7不同处理时间烟草根际土壤微生物对6类碳源相对利用率 Fig. 6 Relative utilization ratio in six groups of carbon sources of tobacco rhizosphere soil under different time treatments by CLP-7 不同字母代表差异显著(P < 0.05) |
如图 7所示, 碳源低利用率预示着微生物对其利用能力的衰退, 碳源高利用率表明微生物对其利用需求较强。施用CLP-7后, 土壤微生物对31种碳源的利用能力显著提高, 变化规律为30 d>CK>50 d>10 d, 与CK差异显著。施菌50 d时, 碳源基本被土壤中微生物完全利用。另外, 随着施菌时间增加, 土壤微生物对大部分氨基酸类碳源(L-精氨酸、L-天冬酰氨酸、L-苯基丙氨酸、L-丝氨酸、L-苏氨酸)、部分羧酸类碳源(D-葡萄氨酸、D-半乳糖醛酸、2-羟苯甲酸)、部分聚合物类(吐温40、肝糖)、双亲类化合物碳源的丙酮酸甲酯和大部分糖类碳源(α-D-乳糖、i-赤藻糖醇、N-乙酰基-葡萄胺)代谢活性有明显的提高, 而对氨基酸类碳源的L-苏氨酸、羧酸类的D-半乳糖醛酸、γ-羟基丁酸和衣康酸、胺类碳源中的苯乙胺和部分糖类碳源, 包括葡萄糖-1-磷酸、D, L-α-甘油和β-甲基-D-葡萄糖苷的代谢活性降低。

|
| 图 7 CLP-7不同处理时间烟草根际土壤微生物对31种碳源利用率 Fig. 7 Utilization ratio of soil microorganism to 31 carbon sources by CLP-7 under different time treatments |
无论是从土壤pH、养分、酶活性的物质代谢, 还是从土壤微生物对碳源的利用率和土壤微生物功能多样性的角度出发, P. koreensis CLP-7有助于改善连作酸化土壤质量, 包括提高土壤pH, 增加土壤速效钾、有效磷、铵态氮、硝态氮含量, 提高土壤脲酶和土壤蔗糖酶活性, 增强土壤微生物功能多样性, 提高对烟草青枯病的防治和促生效果[12]。
已有研究表明, 长期连续耕作可导致严重的土壤酸化、营养失衡和根际微生态环境恶化, 如植烟土壤缓解pH的能力降低、土壤养分含量发生非均衡变化等;当连作达到一定年限时, 土壤pH显著下降, 有机质、全氮、全钾和有效磷含量也出现不同程度的降低[19]。而土壤细菌群的组成、相对丰度和多样性是由土壤pH、土地利用和恢复等因素决定的[5]。本试验在室内添加指示剂培养和田间试验条件下, 验证了P. koreensis CLP-7能一定程度提高其生长环境的pH, 施用于连作导致的酸化植烟土壤后, 烟草根际土壤pH呈升高趋势, 施菌50 d时土样的pH为5.4, 高于CK(pH=5.2), 说明P. koreensis CLP-7可通过生物修复在一定程度上改善土壤酸化。沈灵凤等[20]研究表明, 土壤硝态氮含量与pH值呈极显著负相关关系, 硝态氮在土壤中大量累积会造成土壤pH下降。而本研究中, 施菌后土壤中硝态氮含量呈下降趋势, 进一步验证了CLP-7减少了硝态氮在土壤中的累积, 从而提高土壤pH。因此, 在酸性土壤中施用嗜酸性P. koreensis CLP-7可以通过减少硝态氮的积累缓解土壤酸化, 创造良好的、适宜烟株生长而不利于青枯病等根茎病害发生的土壤生态环境。
据Luo等[21]研究发现, 土壤pH与土壤酶活性正相关且随着连作时间的增加, pH下降的同时土壤酶活性也呈下降趋势。土壤脲酶是一种酰胺酶, 是土壤中唯一一种能将尿素转化为有效氮的酶[22], 能促进土壤尿素水解生成氨, 而氨是植物重要的氮源之一, 间接为植物提供氮源。除此之外, 土壤脲酶还可以催化土壤有机磷化合物的矿化, 直接影响土壤中有机磷的分解、转化及其生物有效性[23]。而本研究发现随CLP-7处理时间的延长, 土壤脲酶活性显著提高, 在施菌30 d达到最高值, 且施菌10 d时, 土壤中铵态氮含量远高于对照组, 烟草根际土壤中可溶性磷含量也随施菌时间增加呈上升趋势, 由此可知CLP-7可通过提高土壤脲酶的活性将土壤中的尿素转化成氨, 同时将土壤中难溶性磷酸盐转化成可溶性磷供烟株吸收, 提高植物对土壤中氮和磷元素的利用, 从而促进植物生长。与徐欣等[24]研究的土壤脲酶活性与氮磷含量呈显著正相关关系基本一致。除此之外, 在本研究中, CLP-7处理的根际土壤中蔗糖酶和过氧化氢酶活性以及养分硝态氮和铵态氮含量均显著提高。而土壤蔗糖酶又称转化酶, 是一种可以把土壤中高分子量蔗糖分解成能够被植物和土壤微生物吸收利用的葡萄糖和果糖的水解酶, 可为土壤生物体提供充分能源, 通常情况下, 土壤肥力越高, 蔗糖酶活性越强, 它不仅能够表征土壤生物学活性强度, 也可以作为评价土壤熟化程度和土壤肥力水平的一个指标[25];过氧化氢酶主要分解土壤中的过氧化氢, 降低土壤中过度累积的过氧化氢对植物根系的危害。土壤蔗糖酶、过氧化氢酶活与土壤中全氮含量极相关。CLP-7菌株不但可以通过影响土壤酶活改善土壤中氮、磷、钾等营养元素的含量, 同时其自身也具有解钾特性[12]。Viruel等[26]指出, 韩国假单胞菌SP28具有溶磷作用以及促进玉米生长特性。由此可知, CLP-7可通过提高酸化土壤酶活和解磷解钾增加氮磷元素含量, 进而促进植株的生长发育, 抑制青枯病的发生。
一般来说, 土壤有机质含量的多少, 是土壤肥力的一个重要指标, 另外, 据吴迪等[27]研究发现, 根际微生物丰富度和多样性指数均与土壤有机质含量呈显著正相关。本试验中CLP-7菌株能显著提高根际土壤有机质含量, 施菌后三个时间处理比CK处理分别增加约62.9%、109.2%和93.8%, 分析其原因可能是土壤有机质含量的升高, 有利于某些微生物数量的增加, 进而能提高根际土壤微生物群落的丰富度, 本试验也验证了这一结论, 但P. koreensis CLP-7可以促进哪类微生物数量的增加, 还有待进一步研究。
植物根际土壤微生物是根部最活跃的部分, 能敏感地反映出土壤生态系统的变化[28]。通过BIOLOG-ECO微平板检测获得的AWCD值提供不同微生物对相同碳源利用的差异性, 反映了微生物对碳源的利用能力, 从功能代谢方面显示了微生物群落功能多样性, 用于描述土壤微生物代谢活性[10]。除此之外, 土壤微生物优势度越大, 则其群落结构受优势种的影响越大, 若其物种多样性指数同时随之增大, 则说明该群落结构越趋于稳定[27], 通常土壤微生物优势度由Shannon指数、Simpson指数、Richness指数、Mclntosh指数和Pielou指数体现。而本试验中CLP-7处理组根际土壤有机质含量显著提高, 分析其原因可能是CLP-7有利于根际土壤中微生物数量和微生物群落丰富度的增加, 进而提高土壤有机质含量, 本推论通过BIOLOG-ECO微平板检测获得的AWCD值和土壤微生物多样性指数变化得到验证。
除了土壤微生物的变化, 有机酸类烟草根系分泌物在R. solanacearum侵染烟草过程中也扮演着重要的作用。前期研究鉴定出了肉桂酸、延胡索酸、肉豆蔻酸能够作为化学信号物, 诱导R. solanacearum的运动趋化活性, 刺激生物膜的形成, 促进烟草青枯病发生, 同时R. solanacearum对多种氨基酸(甘氨酸)、糖类(半乳糖)和有机酸(柠檬酸)具有特殊的吸引力[29]。本研究还发现, 随着施用CLP-7时间的增加, 根际微生物对羧酸类、双亲化合物类、聚合物类和糖类利用率明显提高, 说明偏好羧酸类、双亲化合物类、聚合物类和糖类微生物种群得到富集, 微生物群落结构发生偏单一化现象。糖类、羧酸类和氨基酸类碳源与根系分泌物关系最密切[30], 本研究对土壤微生物代谢功能主成分分析明确了施菌10 d—30 d期间, 烟株根际土壤中微生物对碳源利用差异显著, 主要是氨基酸类、羧酸类和糖类碳源, 与孙家骏等[31]研究的施用生物有机肥对土壤微生物群落功能多样性试验结果一致, 即土壤微生物群落主要是利用碳水化合物、氨基酸和羧酸。再加上R. solanacearum对这三种碳源有特殊吸引力, 因此在施用P. koreensis CLP-7后, 喜好氨基酸类、羧酸类和糖类碳源的土壤微生物增多, 土壤中这三类碳源被彻底利用, 与R. solanacearum竞争氨基酸类、羧酸类和糖类碳源, 从而降低青枯病的发生, 解释了前期试验结果, 即在酸性土壤条件下, P. koreensis CLP-7能推迟烟草青枯病发病10 d, 对烟草青枯病的防效为85.6%, 且高于中性土壤条件下防效71.6%[12]。
综上所述, P. koreensis CLP-7能够明显提高土壤pH和酶活性, 增加土壤养分含量和土壤微生物群落功能多样性, 进而改善酸化植烟土壤质量, 在微生物修复酸化土壤和减轻根茎病害发生的烟草绿色防控中具有较大的应用潜力。
| [1] |
刘艳霞, 李想, 蔡刘体, 石俊雄. 生物有机肥育苗防控烟草青枯病. 植物营养与肥料学报, 2017, 23(5): 1303-1313. |
| [2] |
Li H, Wang J Q, Liu Q, Zhou Z F, Chen F L, Xiang D. Effects of consecutive monoculture of sweet potato on soil bacterial community as determined by pyrosequencing. Journal of Basic Microbiology, 2019, 59(2): 181-191. DOI:10.1002/jobm.201800304 |
| [3] |
王贻鸿.酸碱度对烟草青枯菌(Ralstonia solanacearum)生长及其根部定殖的影响[D].北京: 中国农业科学院, 2017. http://cdmd.cnki.com.cn/Article/CDMD-82101-1017267380.htm
|
| [4] |
Garbeva P, Van Veen J A, Van Elsas J D. Microbial diversity in soil:selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annual Review of Phytopathology, 2004, 42: 243-270. DOI:10.1146/annurev.phyto.42.012604.135455 |
| [5] |
Hartman W H, Richardson C J, Vilgalys R, Bruland G L. Environmental and anthropogenic controls over bacterial communities in wetland soils. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(46): 17842-17847. DOI:10.1073/pnas.0808254105 |
| [6] |
Li S L, Liu Y Q, Wang J, Yang L, Zhang S T, Xu C, Ding W. Soil acidification aggravates the occurrence of bacterial wilt in South China. Front in Microbiology, 2017, 8: 703. DOI:10.3389/fmicb.2017.00703 |
| [7] |
Liu L J, Sun C J, Liu S R, Chai R S, Huang W Q, Liu X X, Tang C X, Zhang Y S. Bioorganic fertilizer enhances soil suppressive capacity against bacterial wilt of tomato. PLoS One, 2015, 10(4): e0121304. DOI:10.1371/journal.pone.0121304 |
| [8] |
柴苗苗. 土壤微生物在促进植物生长方面的作用. 乡村科技, 2019(21): 86-87. DOI:10.3969/j.issn.1674-7909.2019.21.046 |
| [9] |
郑丽萍, 龙涛, 林玉锁, 于赐刚, 刘燕, 祝欣. Biolog-ECO解析有机氯农药污染场地土壤微生物群落功能多样性特征. 应用与环境生物学报, 2013, 19(5): 759-765. |
| [10] |
邹春娇, 齐明芳, 马建, 武春成, 李天来. Biolog-ECO解析黄瓜连作营养基质中微生物群落结构多样性特征. 中国农业科学, 2016, 49(5): 942-951. |
| [11] |
赵兰凤, 张新明, 程根, 张丽娟, 刘小锋, 李华兴. 生物炭对菜园土壤微生物功能多样性的影响. 生态学报, 2017, 37(14): 4754-4762. |
| [12] |
王静, 彭德元, 孔凡玉, 王振华, 王新伟, 李斌, 余祥文, 王贻鸿, 陈德鑫.一株耐酸性的韩国假单胞菌CLP-7及其应用: 中国, 201710081711.X. 2019-08-23.
|
| [13] |
刘海龙.湖北省烟草青枯病的病原学研究[D].武汉: 华中农业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10504-1015380751.htm
|
| [14] |
华菊玲, 刘光荣, 黄劲松. 连作对芝麻根际土壤微生物群落的影响. 生态学报, 2012, 32(09): 2936-2942. |
| [15] |
施河丽, 向必坤, 谭军, 彭五星, 孙玉晓, 王瑞, 吴文昊, 魏国胜, 丁才夫. 烟草青枯病发病烟株根际土壤细菌群落分析. 中国烟草学报, 2018, 24(5): 57-65. |
| [16] |
关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986: 274-328.
|
| [17] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000.
|
| [18] |
孔滨, 杨秀娟. Biolog生态板的应用原理及碳源构成. 绿色科技, 2011(7): 231-234. DOI:10.3969/j.issn.1674-9944.2011.07.100 |
| [19] |
顾美英, 刘洪亮, 李志强, 刘晓伟, 唐光木, 徐万里. 新疆连作棉田施用生物炭对土壤养分及微生物群落多样性的影响. 中国农业科学, 2014, 47(20): 4128-4138. DOI:10.3864/j.issn.0578-1752.2014.20.021 |
| [20] |
沈灵凤, 白玲玉, 曾希柏, 王玉忠. 施肥对设施菜地土壤硝态氮累积及pH的影响. 农业环境科学学报, 2012, 31(07): 1350-1356. |
| [21] |
Luo X S, Fu X Q, Yang Y, Cai P, Peng S B, Chen W L, Huang Q Y. Microbial communities play important roles in modulating paddy soil fertility. Scientific Reports, 2016, 6(1): 20326. DOI:10.1038/srep20326 |
| [22] |
张昀, 可欣, 张广才, 关连珠. 乙草胺对土壤脲酶动力学特征的影响. 植物营养与肥料学报, 2012, 18(04): 915-921. |
| [23] |
Wang Y, Zhang M, Zhao L F, Zhang W, Zhao T, Chu J X, Qiu Y H, Gong H Y, Li R J. Effects of tetrabromobisphenol A on maize (Zea mays L.) physiological indexes, soil enzyme activity, and soil microbial biomass. Ecotoxicology, 2019, 28(1): 1-12. |
| [24] |
徐欣, 王晓军, 谢洪宝, 周珂, 张锦源, 陈一民, 高洪生, 焦晓光. 秸秆腐解对不同氮肥水平土壤脲酶活性的影响. 中国农学通报, 2018, 34(34): 99-102. DOI:10.11924/j.issn.1000-6850.casb18080043 |
| [25] |
骆爱兰, 余向阳. 氟啶胺对土壤中蔗糖酶活性及呼吸作用的影响. 中国生态农业学报, 2011, 19(04): 902-906. |
| [26] |
Viruel E, Erazzú L E, Martínez Calsina L, Ferrero M A, Lucca M E, Siñeriz F. Inoculation of maize with phosphate solubilizing bacteria:effect on plant growth and yield. Journal of Soil Science and Plant Nutrition, 2014, 14(4): 819-831. |
| [27] |
张建峰, 何佩佩, 周奇, 马丽娜, 田磊, 田春杰. 五大连池火山岩土壤发育过程中根际细菌多样性分析. 东北农业大学学报, 2017, 48(05): 65-71. |
| [28] |
李兰君, 刘玳含, 刘建斌, 武凤霞, 朱英波. 连作对设施番茄土壤微生物及酶活性的影响. 江苏农业科学, 2018, 46(18): 130-134. |
| [29] |
Li Sh L, Chen X, Jiao W, Guo B, Liang Y, Chen J N, Wei D. Cinnamic, Myristic and Fumaric Acids in Tobacco Root Exudates Induce the Infection of Plants by Ralstonia Solanacearum. Plant and Soil, 2017, 412.1-2: 381-95. |
| [30] |
胡婵娟, 傅伯杰, 刘国华, 靳甜甜, 刘宇. 黄土丘陵沟壑区典型人工林下土壤微生物功能多样性. 生态学报, 2009, 29(02): 727-733. DOI:10.3321/j.issn:1000-0933.2009.02.022 |
| [31] |
孙家骏, 付青霞, 谷洁, 王小娟, 高华. 生物有机肥对猕猴桃土壤酶活性和微生物群落的影响. 应用生态学报, 2016, 27(03): 829-837. |