 2020, Vol. 40
2020, Vol. 40文章信息
- 罗庆辉, 徐泽源, 许仲林
- LUO Qinghui, XU Zeyuan, XU Zhonglin
- 天山雪岭云杉林生物量估测及空间格局分析
- Estimation and spatial pattern analysis of biomass of Picea schrenkiana forests
- 生态学报. 2020, 40(15): 5288-5297
- Acta Ecologica Sinica. 2020, 40(15): 5288-5297
- http://dx.doi.org/10.5846/stxb201904240841
-
文章历史
- 收稿日期: 2019-04-24
- 网络出版日期: 2020-05-21

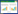




2. 新疆大学资源与环境科学学院绿洲生态教育部重点实验室, 乌鲁木齐 830046
2. Key Laboratory of Oasis Ecology, College of Resource and Environmental Science, Xinjiang University, Urumqi 830046, China
森林生态系统作为陆地生态系统的主体, 在全球碳循环过程中起着决定性作用。同时, 森林生态系统植被所固定的碳量约占陆地植被总固碳量的82.5%[1], 森林生物量约占陆地生态系统生物量的90%[2], 在研究全球碳循环、全球气候变化和减缓大气温室气体浓度上升等方面具有重要意义。近年来, 森林生物量估算已成为现代林业研究的热点问题, 很多学者对其进行了研究[3-5]。
传统的森林生物量样地调查方法, 准确度高, 但耗时耗力, 而且难以精确了解森林分布状态。遥感技术虽然准确度较低, 但能够监测森林大面积覆盖范围, 已成功应用到森林生物量估算中[6-9]。天山雪岭云杉林是新疆天山山地森林的优势种, 同时也是亚洲中部山地的特有种, 在新疆水源涵养、水土保持和气候调控等方面发挥着重要的作用[10]。郑拴丽[11]和刘贵峰等[12]通过野外实测样地数据测定和研究了天山不同林场雪岭云杉林生物量, 此外, 李虎等[13]建立了雪岭云杉林生物量模型, 估算了西天山雪岭云杉林生物量;然而采用遥感方法应用多元逐步回归法、偏最小二乘法和主成分分析法估算整个天山雪岭云杉林生物量的对比研究鲜见报道。本文以天山雪岭云杉林作为研究对象, 将Landsat 8 OLI影像和实测数据相结合, 采用多元逐步回归法、偏最小二乘回归法和主成分分析法建立雪岭云杉林生物量估算模型, 并对三种模型的估算能力进行对比分析。对森林生物量进行估测时, 各生物量自变量因子间可能存在严重的多重共线性, 多元回归法解决不了该问题, 因此本研究不考虑多元回归法建模, 尽可能多地选取植被指数、气候因子、地学信息及影像信息等可能与雪岭云杉林生物量有关的自变量, 利用多元逐步回归法、偏最小二乘回归法和主成分分析法建立雪岭云杉林生物量遥感估测模型, 择优对天山雪岭云杉林生物量进行估测和评价。本研究方法将为天山雪岭云杉林生物量模型的构建提供新的途径, 研究结果对于了解天山雪岭云杉林生物量空间分布和保护雪岭云杉林生态系统可持续管理具有重要意义。
1 研究区概况天山位于塔克拉玛干沙漠的北部和西部地区, 塔里木盆地以北[14], 在我国新疆境内长达1760 km, 分布在天山北坡中山带海拔1500—2800 m处的雪岭云杉林是新疆山地森林分布最广的树种, 而且雪岭云杉是天山最主要的地带性森林植被[15-16]。天山的植被分布具有明显的垂直带性, 从低海拔到高海拔, 依次为山地荒漠草原、山地草原、山前灌木、山地针叶林、亚高山灌丛草甸、高山草甸、高山垫状植被[17]。研究区为海拔1500—2800 m的山地针叶林(建群种为雪岭云杉林)分布区域, 属于温带大陆性气候, 土壤是山地灰褐色森林土。主要地被植物有黑果小檗(Berberis heteropoda)、红果小檗(Berberis nummularia)、忍冬(Lonicera sp.)、天山花楸(Sorbus tianschanica)、黑果荀子(Cotoneastermelanocarpus)、蔷薇(Rosa sp.)、绣线菊(Spiraeahypericifolia)、老鹳草(Geraniumdahuricum)、天山雨衣草(Alchemilla tianschanica)、羊角芹(Aegopodiumpodagraria)等[18]。

|
| 图 1 研究区雪岭云杉林采样点的空间分布 Fig. 1 Distribution of sampling points in the study area |
本研究采用2017年6月至9月Landsat-8 OLI遥感数据。天山雪岭云杉林分布范围涉及8景Landsat-8 OLI影像, 其行列号分别是:143条带的30行;144条带的30行;145条带的29和30行;146条带的29和30行;147条带的29和30行。对8景Landsat-8 OLI影像分别进行几何校正、辐射定标和FLAASH大气校正, 然后进行拼接和裁剪, 得到研究区地表反射率图像。本研究所用Landsat-8 OLI影像所用波段为波段2, 波段3, 波段4, 波段5, 波段6, 波段7, 投影方式为UTM-WGS84, 空间分辨率为30.0 m。
本研究采样点位于分布于成片雪岭云杉天然林的地段, 林地没有明显的人类干扰和自然破坏情况。首先在样地中确定一棵中心树, 用激光测距仪测出半径为10 m的样圆, 在样圆内的每株树底部插上写有标号的小旗, 将样圆内的所有树木每木检尺(胸径大于5 cm), 用激光测高仪测量树高, 使用手持GPS记录样地的经度、纬度、海拔等数据。通过对研究区进行野外采样调查, 共计66块样地, 每块样地面积为0.0314 hm2。根据样地每木检尺数据及已建立的天山云杉林器官生物量估算方程[19]计算每株树木生物量(表 1), 最终求出样地总生物量。在Landsat-8 OLI影像上提取样地中心点所在像元及相邻4个像元的灰度值, 求出平均值。将66块样地随机分为2部分, 其中46块样地用于建模, 20块样地用于检验。
| 模型Model | 模型参数Model parameter | ||
| a | b | ||
| W=a(D2H)b | 树干生物量 | 0.033759 | 0.907882 |
| W=a(D2H)b | 树皮生物量 | 0.005498 | 0.830362 |
| W=a(D2H)b | 树枝生物量 | 0.019814 | 2.602171 |
| W=a(D2H)b | 树叶生物量 | 0.027648 | 2.252940 |
| W=a(D2H)b | 树根生物量 | 0.042731 | 2.187144 |
本研究尽可能多地选出与生物量有关的各种自变量因子来估测天山雪岭云杉林生物量, 包括遥感影像各波段灰度值、不同波段灰度值之间的线性和非线性组合(常用的植被指数)(表 2)、气候因子及地形因子如海拔、坡度、坡向和坡位等, 共计42个自变量因子。将各自变量因子与对应样地生物量作相关系数显著性检验, 结果如表 3所示, 与样地生物量相关性较高且关系显著的自变量因子包括:容重, 经度, 纬度, 高程, b2, b3, b4, b12, b13, b16, b18等。
| 植被指数Vegetation index | 计算公式Calculation formula |
| 差值植被指数Different vegetation index | DVI=B4-B3 |
| 比值植被指数Ratio vegetation index | RVI=B4/B3 |
| 土壤调整植被指数Soil adjusted vegetation index | SAVI=(1+L)×(B4-B3)/(B4+B3) |
| 归一化植被指数Normalized difference vegetation index | NDVI=(B4-B3)/(B4+B3) |
| 修正的土壤调整植被指数Modified soil-adjusted vegetation index |  |
| 式中:B4为近红外波段;B3为红光波段;L为土壤调节因子, 通常取0.5[20] | |
| 变量 Vari |
相关系数 Correlation coefficient |
|
| 原始波段 Original |
Band2 | 0.016 |
| Band3 | -0.059 | |
| Band4 | -0.060 | |
| Band5 | 0.018 | |
| Band6 | -0.010 | |
| Band7 | -0.007 | |
| 植被指数 Vegetati |
DVI | -0.203 |
| RVI | 0.094 | |
| SAVI | -0.139 | |
| NDVI | -0.139 | |
| MSAVI | -0.098 | |
| 土壤因子 Soil fac |
有机碳含 | -0.014 |
| 沙含量 | 0.235 | |
| pH | -0.064 | |
| 容重 | 0.349* | |
| 有效含水 | -0.106 | |
| 气候因子Climatic factor | b1 | -0.122 |
| b2 | 0.339* | |
| b3 | 0.336* | |
| b4 | -0.336* | |
| b5 | -0.169 | |
| b6 | -0.180 | |
| b7 | -0.030 | |
| b8 | -0.313 | |
| b9 | -0.010 | |
| b10 | -0.263 | |
| b11 | 0.027 | |
| b12 | 0.318* | |
| b13 | 0.291* | |
| b14 | 0.242 | |
| b15 | -0.264 | |
| b16 | 0.304* | |
| b17 | 0.274 | |
| b18 | 0.324* | |
| b19 | 0.274 | |
| 地学信息 Geographic information |
经度 | -0.505** |
| 纬度 | -0.432** | |
| 高程 | 0.374* | |
| 坡度 | -0.010 | |
| 坡向 | -0.225 | |
| 坡位 | 0.008 | |
| 林分密度 Stand density |
-0.016 | |
| b1为年平均温度, b2为日平均范围, b3为等温性, b4为季节性温度, b5为最热月份最高温度, b6为最冷月份最低温度, b7为年平均温度范围, b8为最湿季平均温度, b9为最干季平均温度, b10为最热季平均温度, b11为最冷季平均温度, b12为年平均降水量, b13为最湿月份降水量, b14为最干月份降水量, b15为季节性降水量, b16为最湿季降水量, b17为最干季降水量, b18为最暖季降水量, b19为最冷季降水量; *:α = 0.05;**:α = 0.01 | ||
多元逐步回归法可以避免多元线性回归模型中存在的共线性和个别自变量解释因变量的贡献率较小的问题, 同时可以剔除对因变量不显著的自变量, 筛选出显著性因子强的变量。对样本数据进行标准化处理, 利用SPSS 22.0软件采用逐步回归法建立多元逐步回归模型。通过检验的变量为经度、容重和b13。由表 4可知, 模型3的决定系数最大(R2=0.456), 并且呈极显著性关系(P<0.01), 因此, 模型3为最优生物量估算模型;模型形式如下:
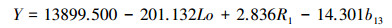
|
式中:Y为样地生物量;Lo为经度;R1为容重;b13为最湿月份降水量。
| 模型Models | 相关系数(R) Correlation coefficient |
决定系数(R2) Coefficient of determination |
F | 显著性 Significance |
| B=8480.462-96.803Lo | 0.505 | 0.255 | 15.078 | 0.000 |
| B=3691.127-96.223Lo+3.203R1 | 0.611 | 0.374 | 12.828 | 0.000 |
| B=13899.500-201.132Lo+2.836R1-14.301b13 | 0.675 | 0.456 | 11.745 | 0.000 |
偏最小二乘回归是一种将多元线性回归分析、主成分分析和相关分析相结合, 在算法中实现回归建模、数据结构简化的分析方法, 由Wold等[21]于1983年提出, 在处理自变量多、变量间有多重相关性等问题时具有极大的优势[21-22], 算法实现均在Matlab2016a软件中进行。偏最小二乘回归的具体步骤可以参考《偏最小二乘回归的线性与非线性方法》[23]。
回归分析和模型建立的一个重要内容是多重共线性诊断。通常诊断的方法主要有:(1)特征根条件指数和分解比例:若条件指数值在10与30间为弱相关, 在30与100间为中等相关, 大于100表示有强相关;若最大条件指数≥10, 且2个或多个的估计回归系数有较大的方差分解比, 一般大于0.5时, 可认为自变量间存在共线性;(2)自变量间的相关系数法:当相关系数值趋近于1时, 表示相应的自变量之间有较强的线性相关关系, 若变量间的相关系数的绝对值大于0.7, 表明自变量之间的线性相关性较强;(3)根据方差膨胀因子(VIF)判断:VIF值越大, 相关系数越接近于1, 说明对应自变量间的共线性越强;当VIF≥10时, 可认为自变量间存在严重共线性[24]。本研究以天山雪岭云杉林生物量各环境因子和遥感信息为自变量, 分别计算了生物量各自变量因子的特征值、条件数、方差分解比例, 得知各因子间的条件数为13386.556, 根据特征根条件指数和方差分解比例综合分析可知, 生物量各自变量因子间存在很强的多重共线性, 需要用偏回归分析方法进行生物量估测研究。
3.2.2 偏最小二乘回归模型的建立本文采用46个样地生物量数据及与生物量相关性较高的11个自变量进行偏最小二乘回归, 当提取3个有效成分时, 偏最小二乘回归的模型表达式为:
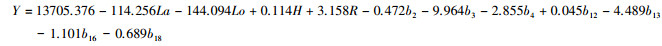
|
式中, La为纬度;H为高程;b2为平均月较差;b3为等温性;b4为季节性温度;b12为年平均降水量;b16为最湿季降水量;b18为最暖季降水量。
3.3 基于主成分分析的生物量模型多元线性回归法或逐步回归法多被用于建立森林生物量与遥感和地学因子的模型, 但回归模型中的各变量可能存在共线性问题, 进而影响模型的精度, 最终出现病态模型[25]。采用主成分分析的方法, 提取各自变量因子的主成分, 再建立主成分与生物量之间的回归模型, 既可以保留多个自变量因子的主要信息, 达到降维的目的, 又可以避免变量之间多重相关问题。
3.3.1 主成分提取及因子载荷分析利用SPSS22.0软件中的因子分析功能对46个样本相关性较高的12个变量因子进行主成分提取。对自变量因子进行KMO和Bartlett的球状检验, KMO值:0.722>0.6, Bartlett的球状检验呈显著情况(P < 0.05), 表明可以进行主成分提取。
从表 5可知, 前3个主成分的累计贡献率达到了81.92%, 反映了自变量因子样本的主要信息, 变量的数量降为3个, 在保留了样本的主要信息的同时, 又起到了降维和简化模型的作用。根据载荷分析可知, 第一主成分PC1主要反映了经度, b12, b13, b16和b18的主要信息;第二主成分PC2主要反映b2, b3, b4, 海拔, 纬度和DVI;第三主成分PC3反映的信息已经很少, 主要反映了容重的信息。
| 主成分 Principal components |
第一主成分 First principal component |
第二主成分 Second principal component |
第三主成分 Third principal component |
| 纬度Latitude | -0.541 | 0.342 | -0.084 |
| 经度Longitude | -0.922 | -0.117 | -0.171 |
| 海拔Altitude | 0.614 | 0.409 | 0.199 |
| 容重Bulk density | -0.051 | -0.064 | 0.803 |
| 平均日较差Mean diurnal range | 0.798 | -0.532 | -0.157 |
| 等温性Isothermality | 0.832 | -0.496 | -0.159 |
| 季节性温度Temperature seasonality | -0.873 | 0.368 | 0.193 |
| 年平均降水量Annual precipitation | 0.951 | 0.201 | -0.004 |
| 最湿月份降水量Precipitation of wettest month | 0.932 | 0.315 | 0.011 |
| 最湿季降水量Precipitation of wettest quarter | 0.937 | 0.306 | 0.013 |
| 最暖季降水量Precipitation of warmest quarter | 0.941 | 0.303 | 0.033 |
| 差值植被指数Different vegetation index | -0.160 | 0.475 | -0.572 |
| 特征值Eigenvalue | 7.176 | 1.517 | 1.137 |
| 贡献率Contribution /% | 41.337 | 30.847 | 9.736 |
| 累计贡献率Cumulative contribution /% | 41.337 | 72.184 | 81.920 |
利用已建立的3个主成分方程, 计算46个样地的3个主成分值。将主成分值作为自变量, 样地生物量作为因变量, 建立森林生物量的估测模型;由表 6可知, 森林生物量各自变量因子的主成分和生物量具有显著性相关关系, 相关系数R=0.619;模型形式如下所示:
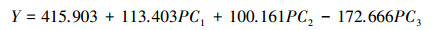
|
| 方差来源 Source of variance |
平方和 Sum of square |
df | 均方 Mean square |
F |
| 回归Regression | SSR=2371775 | 3 | MSR=790592 | 8.688 |
| 残差Residual | SSE=3822038 | 42 | MSE=91001 | |
| 总和Sum | SST=6193813 | 45 |
式中:Y:森林生物量(t/hm2);PC1-PC3:分别为3个主成分值
4 讨论 4.1 天山雪岭云杉林生物量估测模型的比较为检验多元逐步回归法、偏最小二乘回归法和主成分分析法构建天山雪岭云杉林生物量模型的精度, 本文利用未参与模型构建的20个样地实测生物量数据对各模型进行预测精度检验(图 2)。由计算可知, 多元逐步回归模型的平均拟合精度为69.07%, 绝对误差为64.50 t/hm2, 平均相对误差为10.89%;偏最小二乘模型的平均拟合精度为74.36%, 绝对误差为144.94 t/hm2, 平均相对误差为28.78%, 该模型可以用来估测天山雪岭云杉林的生物量;主成分回归模型生物量实测值和预测值呈极显著正相关关系(R2=0.730, P < 0.01), 且平均估算精度达到71.22%;说明由主成分分析得到的该模型也可以用来估测天山雪岭云杉林的生物量。

|
| 图 2 20个样地实测值和预测值比较 Fig. 2 Comparisons between measured and predicted biomass in 20 samples |
通过对比三种生物量估测方法所建立的生物量估算模型, 表明偏最小二乘回归法在解释变量较多、样本数量较少且各变量间存在严重的多重共线性时要优于多元逐步回归法和主成分分析法;利用偏最小二乘回归法建立的样地生物量模型基本满足生物量估算精度的要求, 更适合估算天山雪岭云杉林分布区域的森林生物量。
近年来森林生物量估测方法有很多, 包括树木年轮法[26]、相对生长关系法[27]、生物量估算参数法[28]和3S法[29]等, 如何提高估测精度是当前森林生态学研究的关键。本研究中偏最小二乘法对天山雪岭云杉林生物量有较好的拟合和估测效果, 但对于各自变量因子估算生物量的精度仍有很大的提升空间。随着“3S”技术的进一步发展, 气象网络的加密和野外调查数据准确度的提高等[30], 气象数据、算法和遥感信息相结合对于森林生态学的发展会有更大的促进作用。
4.2 天山雪岭云杉林生物量的空间分布特征 4.2.1 天山雪岭云杉林生物量与经纬度的关系天山雪岭云杉林生物量随纬度的增加而降低, 回归表明样地生物量的38.63%可以用纬度来解释(P < 0.01), 生物量最高值分布在42°—43°N区域(图 3);生物量随经度的增加呈现降低的变化趋势, 回归表明样地生物量的25.38%可以用经度来解释(P < 0.01), 生物量最高值分布在80°—82°E区域(图 3)。综上所述, 研究区雪岭云杉林生物量在空间上具有明显的分布规律, 整体上呈现出天山西部生物量水平较高, 中部和东部生物量水平较低的趋势(图 3), 这与刘贵峰等[12]和许文强等[31]的研究结论一致, 产生这种现象的主要原因是因为天山中部和东部的土壤养分含量低于天山西部, 且降水量较少, 蒸腾作用较强, 影响了雪岭云杉林的正常生长, 体现了雪岭云杉林生物量在空间上对生境条件适应的差异性。

|
| 图 3 天山雪岭云杉生物量随经纬度的变化 Fig. 3 Biomass of Picea schrenkiana forest with latitude and longitude |
地形因子作为生境条件的一种综合指示, 一般包括海拔、坡度、坡向和坡位等等, 不仅决定光、热、水、土壤等植被生境要素的空间分布, 还影响着植被群落分布和种群格局[32-33]。
海拔的变化综合了光照、温度等各种环境要素, 导致水热因子产生差异进一步影响植物群落的结构组成和生产力[34-35]。本研究表明, 随着海拔的升高, 研究区样地生物量呈“单峰”变化趋势, 这与李凯辉等[35]和张雷等[36]的研究结果一致。雪岭云杉林生物量在海拔2100—2400 m处生物量最大, 说明该海拔区段最适宜天山雪岭云杉林的生长;自该海拔段向下, 温度升高, 降水减少, 土壤蒸发增强导致土壤水分减少, 抑制了雪岭云杉林的生长, 使得雪岭云杉林生物量减小;自该海拔向上, 温度降低, 同时天山高海拔冻土层阻碍了雪岭云杉林根系对水分的获取, 因此生物量减小。从图 4可以得知, 天山雪岭云杉林主要分布于天山北坡的上坡和山脊处, 其生物量随坡度增加的变化趋势较复杂, 随着坡向的增大呈现递减的趋势, 表明地形中的坡度和坡位因子不是影响雪岭云杉林生物量变化的决定性因素, 其主要原因是, 坡度和坡位的变化影响着雪岭云杉林生长发育过程中对林下土壤水分的需求, 而坡向的变化影响着雪岭云杉林对光照的需求, 在本研究区内坡度和坡位因子变化幅度较小, 因此对雪岭云杉林生物量的影响居于海拔和坡向因子之下。

|
| 图 4 天山雪岭云杉生物量随地形因子的变化 Fig. 4 Biomass of Picea schrenkiana forest with topographical factors 数据为平均值±标准误差, n=66;图中各组间标不同字母者表示组间差异显著(P < 0.05) |
综上所述, 由于经纬度、海拔、坡度、坡向和坡位等的变化, 导致了降水量、温度、水分和光照等条件的不同, 进一步引起了研究区天山雪岭云杉林生物量空间分布随经纬度和地形因子的规律性变化趋势。此外, 实际环境中影响森林生物量的因素还有很多, 如湿度、土壤养分、CO2等等, 这些因素在地里空间上具有明显的差异性。从生态位角度出发, 这些因素会共同构成一个多维资源空间[37], 在不同资源的配置下会使天山雪岭云杉林生物量在空间上呈现较大的差异性。因此从生态位角度来研究天山雪岭云杉林生物量的空间分布特征是将来研究的重点问题。
5 结论本文以天山雪岭云杉林为研究对象, 基于Landsat-8 OLI遥感影像数据和野外实测数据, 构建了天山雪岭云杉林生物量估算模型并进行空间格局分析。主要结论如下:(1)分别采用逐步回归分析法、偏最小二乘回归法和主成分分析法建立了天山雪岭云杉林生物量估测模型, 各模型平均拟合精度分别为69.07%、74.36%和71.22%, 表明偏最小二乘回归法构建的模型拟合效果更好, 预测精度最高;本研究利用遥感手段估测大面积森林生物量空间分布的成果, 对于提高区域尺度的森林健康评价具有重要意义。(2)本研究尽可能多地考虑了与森林生物量有关的各种自变量因子, 包括与遥感信息相关的各种植被指数、波段值之间的组合、土壤因子和气候因子以及地学因子。此外, 本研究还需确定更合理的取样方案, 扩大取样范围, 同时考虑研究区内雪岭云杉林的林分结构、土壤立地条件等生态环境因子, 提高模型对研究区大面积生物量估算的精度和稳定性。(3)天山雪岭云杉林生物量在空间上呈西部高, 中部和东部低的趋势, 主要是由于天山西部的水热条件优于中部和东部, 这与王文栋等[38-39]的研究结果相符合。森林生物量的空间分布格局是多种因素综合作用的结果, 不仅与经度、纬度和地形因子有关, 同时还与研究区地貌特征、土壤类型和采样地的小气候密切相关, 因此下一步工作将结合其他环境因子数据深入分析, 以揭示影响天山雪岭云杉林生物量空间分布格局的主导因素。
| [1] |
Sabine C L, Heimann M, Artaxo P, Bakker D C E, Chen C T A, Field C B, Gruber N, Quere L C, Prinn R G, Richey J E, Lankao P R, Sathaye J A, Valentini R.Current Status and Past Trends of the Global Carbon Cycle.The global carbon cycle.Integrating humans, climate and the natural world. Island Press, 2004.
|
| [2] |
刘广文. 森林与碳循环. 现代农业科技, 2007(19): 220-221. |
| [3] |
苏华, 李静, 陈修治, 廖吉善, 温达志. 基于森林群落和光谱曲线特征分异的福建省森林生物量遥感反演. 生态学报, 2017, 37(17): 5742-5755. |
| [4] |
秦立厚, 张茂震, 袁振花, 杨海宾. 基于人工神经网络与空间仿真模拟的区域森林碳估算比较——以龙泉市为例. 生态学报, 2017, 37(10): 3459-3470. |
| [5] |
陆君, 刘亚风, 齐珂, 樊正球. 福州市森林碳储量定量估算及其对土地利用变化的响应. 生态学报, 2016, 36(17): 5411-5420. |
| [6] |
王新云, 郭艺歌, 何杰. 基于HJ1B和ALOS/PALSAR数据的森林地上生物量遥感估算. 生态学报, 2016, 36(13): 4109-4121. |
| [7] |
庞勇, 蒙诗栎, 李增元. 机载高分辨率遥感影像的傅氏纹理因子估测温带森林地上生物量. 林业科学, 2017, 53(3): 94-104. |
| [8] |
Zhao K G, Popescu S, Nelson R. Lidar remote sensing of forest biomass:a scale-invariant estimation approach using airborne lasers. Remote Sensing of Environment, 2009, 113(1): 182-196. DOI:10.1016/j.rse.2008.09.009 |
| [9] |
Wang X, Wang S, Dai L. Estimating and mapping forest biomass in northeast China using joint forest resources inventory and remote sensing data. Journal of Forestry Research, 2018, 29(3): 1-15. |
| [10] |
刘萍, 张国林, 黄涛. 不同地域天山云杉林分结构及森林经营措施研究. 林业资源管理, 2013(1): 45-48. DOI:10.3969/j.issn.1002-6622.2013.01.009 |
| [11] |
郑拴丽. 新疆天山雪岭云杉和阿尔泰山西伯利亚落叶松生物量、碳储量及空间分布格局研究. 新疆大学, 2016. |
| [12] |
刘贵峰, 刘玉平, 郭仲军, 白志强. 天山云杉林生物量及其变化规律的研究. 西北林学院学报, 2013, 28(5): 13-17. DOI:10.3969/j.issn.1001-7461.2013.05.03 |
| [13] |
李虎, 慈龙骏, 方建国, 陈冬花, 刘玉锋. 新疆西天山云杉林生物量的动态监测. 林业科学, 2008, 44(10): 14-19. |
| [14] |
常亚鹏, 李路, 许仲林. 天山北坡雪岭云杉林地开垦的土壤有机碳损失估算. 生态学报, 2017, 37(4): 1168-1173. |
| [15] |
王慧杰, 常顺利, 张毓涛, 李翔, 韩燕梁. 天山雪岭云杉林土壤有机碳密度空间分异及其与森林发育的关系. 山地学报, 2017, 35(3): 300-307. |
| [16] |
张艳静, 于瑞德, 郑宏伟, 甘淼, 杨美琳, 石冰冰. 天山东西部雪岭云杉径向生长对气候变暖的响应差异. 生态学杂志, 2017, 36(8): 2149-2159. |
| [17] |
谢锦, 常顺利, 张毓涛, 王慧杰, 宋成程, 何平, 孙雪娇. 天山北坡植物土壤生态化学计量特征的垂直地带性. 生态学报, 2016, 36(14): 4363-4372. |
| [18] |
阿米娜木·艾力, 常顺利, 张毓涛, 仇瑶, 何平. 天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素. 生态学报, 2014, 34(7): 1626-1634. |
| [19] |
刘广路. 天山云杉生长规律与天山植物群落生产力研究. 河北农业大学, 2006. |
| [20] |
陶立超. 白马林场森林碳储量遥感估测. 北京林业大学, 2014. |
| [21] |
Wold S, Ruhe A, Wold H, W. J. DUNN. The Collinearity Problem in Linear Regression.The Partial Least Squares (PLS) Approach to Generalized Inverses. Siam Journal on Scientific & Statistical Computing, 1984, 5(3): 735-743. |
| [22] |
陈长生, 徐勇勇, 赵东涛. 医学重复观测数据裂区方差分析的假定条件的检验. 医学争鸣, 2001, 22(7): 642-644. |
| [23] |
王惠文. 偏最小二乘回归的线性与非线性方法. 国防工业出版社, 2006.
|
| [24] |
周贝贝, 张强, 孙健, 李兴亮, 魏钦平. 偏最小二乘回归在苹果土壤养分与果实品质关系的研究与应用. 农业科学与技术:英文版, 2016, 17(2): 362-366. DOI:10.3969/j.issn.1009-4229.2016.02.027 |
| [25] |
徐文科, 蔡体久, 琚存勇. 基于RS和GIS的毛乌素沙地荒漠化程度定量估测. 林业科学, 2007, 43(5): 48-53. |
| [26] |
SH AO, Quanqin, HUANG Lin, LIU Jiyuan, YANG Haijun, CHEN Zhuoqi. Dynamic analysis on carbon accumulation of a plantation in Qianyanzhou based on tree ring data. Journal of Geographical Sciences, 2009, 19(6): 691-706. DOI:10.1007/s11442-009-0691-y |
| [27] |
Ketterings Q M, Coe R, Noordwijk M V, Ambagau Y, Palm C A. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. Forest Ecology & Management, 2001, 146(1): 199-209. |
| [28] |
Guo Z, Fang J, Pan Y, Birdsey R. Inventory-based estimates of forest biomass carbon stocks in China:A comparison of three methods. Forest Ecology & Management, 2010, 259(7): 1225-1231. |
| [29] |
Luo Y J, Zhang X Q, Wang X K, Zhu J H, Hou Z H, Zhang Z J. Forest biomass estimation methods and their prospects. Scientia Silvae Sinicae, 2009, 45(8): 129-134. |
| [30] |
张正健, 刘志红, 郭艳芬, 韩建宁, 李扬. 偏最小二乘在遥感监测西藏草地生物量上的应用. 草地学报, 2009, 17(6): 735-739. |
| [31] |
许文强, 杨辽, 陈曦, 高亚琪, 王蕾. 天山森林生态系统碳储量格局及其影响因素. 植物生态学报, 2016, 40(4): 364-373. |
| [32] |
孔宁宁, 曾辉, 李书娟. 四川卧龙自然保护区植被的地形分异格局研究. 北京大学学报, 2002, 38(4): 543-549. DOI:10.3321/j.issn:0479-8023.2002.04.016 |
| [33] |
沈泽昊, 金义兴, 张新时. 地形对亚热带山地景观尺度植被格局影响的梯度分析. 植物生态学报, 2000, 24(4): 430-435. DOI:10.3321/j.issn:1005-264X.2000.04.008 |
| [34] |
柳妍妍, 王鑫, 公延明. 天山南坡中段高寒草地物种多样性与生物量的垂直分异特征. 生态学杂志, 2013, 32(2): 311-318. |
| [35] |
李凯辉, 胡玉昆, 王鑫, 范永刚, 吾买尔·吾守. 不同海拔梯度高寒草地地上生物量与环境因子关系. 应用生态学报, 2007, 18(9): 2019-2024. |
| [36] |
张雷, 于澎涛, 王彦辉, 王顺利, 刘贤德. 祁连山北坡青海云杉中龄林生物量随海拔的变化. 林业科学, 2015, 51(8): 1-7. |
| [37] |
Hutchinson GE. Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology, 1957, 22: 415-427. DOI:10.1101/SQB.1957.022.01.039 |
| [38] |
王文栋, 李建贵, 张毓涛, 李吉玫, 李翔, 芦建江. 天山林区不同地段6种灌木生物量组成. 水土保持研究, 2015, 22(5): 127-132. |
| [39] |
王文栋, 白志强, 阿里木·买买提, 刘端, 郭忠军. 天山林区6种优势种灌木林生物量比较及估测模型. 生态学报, 2016, 36(9): 2695-2704. |