 2020, Vol. 40
2020, Vol. 40文章信息
- 徐艺逸, 曹敏, 徐国瑞
- XU Yiyi, CAO Min, XU Guorui
- 云南省三种典型气候带凋落物层弹尾类多样性分布格局
- Diversity distribution patterns of Collembola in litter layers along three typical climate zones in Yunnan Province
- 生态学报. 2020, 40(14): 5008-5017
- Acta Ecologica Sinica. 2020, 40(14): 5008-5017
- http://dx.doi.org/10.5846/stxb201904290879
-
文章历史
- 收稿日期: 2019-04-29
- 网络出版日期: 2020-04-28





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
土壤动物作为重要分解者, 在生态系统的物质循环与营养供应中起着重要作用。弹尾类作为土壤动物中优势类群, 数量巨大、分布广泛、食性多样, 在土壤食物网占有关键位置[1-2]。同时, 弹尾类通过对植物木质部的取食和对凋落物的破碎作用参与分解作用, 促进微生物继续分解凋落物[3]。弹尾类通过直接或间接的方式影响土壤团聚体的形成, 进而对生物地球化学过程、水分渗透、气体交换以及抵抗侵蚀损失等方面产生重要影响[2]。在全球变化背景下, 对不同气候带下弹尾类多样性沿海拔分布格局的研究有助于进一步理解土壤动物多样性如何响应气候变化。
之前的研究表明弹尾类多样性在亚热带高于寒温带, 影响其多样性分布的环境因子为温度[4];亚热带弹尾类多样性随海拔升高而升高, 且与土壤pH呈负相关[5], 在温带的研究也得到相同的结果[6];也有研究表明, 在亚热带弹尾类的丰富度和数量随海拔呈单调上升格局且与土壤有机质含量正相关[7];而在热带弹尾类多度随海拔升高而降低, 其主要与温度呈正相关[8]。在不同气候带的研究中弹尾类的多样性沿海拔分布格局差异较大, 主要影响其分布格局的环境因素也不尽相同。这些研究多数具有气候带的局限性, 经度差异较大, 无法去除经度差异带来的影响。
可以看出, 以往研究很少关注弹尾类多样性跨气候带的分布格局, 尤其是在同一经度不同纬度梯度下。云南省分布着热带、亚热带和亚高山三种典型气候带, 是全球生物多样性研究的热点地区, 为研究不同气候带下弹尾类多样性沿海拔分布格局创造了地理优势。本研究在云南省内同一经度(E 101°)的热带雨林(西双版纳补蚌, 800—1400 m), 亚热带常绿阔叶林(哀牢山, 2000—2600 m)和亚高山针叶林(玉龙雪山, 3200—3800 m)展开, 样地均处于青藏高原的东缘且经度较为一致, 最大程度减少了经度、地形差异和地质历史对研究结果造成的影响。通过调查与分析弹尾类多样性沿气候带的分布格局与环境因子, 回答如下科学问题:(1)弹尾类多样性是否沿热带、亚热带、亚高山气候带呈递减格局?(2)影响弹尾类多样性沿不同气候带分布格局的环境因素是什么?针对以上科学问题, 我们做出如下科学假设:(1)弹尾类多样性沿热带、亚热带、亚高山气候带降低;(2)温度是影响不同气候带下弹尾类多样性分布格局的主要环境因子。
1 研究地区与方法 1.1 研究地概况本研究在云南省3个典型气候带开展, 分别为热带(西双版纳补蚌地区21°08′—22°36′N, 99°56′—101°50′E)、亚热带(哀牢山千家寨地区23°36′—24°56′N, 100°44′—101°14′E)和亚高山(丽江玉龙雪山27°10′—27°40′N, 100°10′—100°20′E)。在每个气候带设置4个海拔样带, 海拔样带间的垂直高差200 m, 不同气候带间的高差为600 m。具体而言包括热带(800 m、1000 m、1200 m、1400 m)、亚热带(2000 m、2200 m、2400 m、2600 m)、亚高山(3200 m、3400 m、3600 m、3800 m), 每个海拔带上选择4个20 m × 20 m的样地进行土壤动物取样。
| 研究地区 Study site |
气候带 Climate zone |
植被类型 Vegetation type |
年降水量/mm Mean annual precipitation |
年均温/℃ Mean annual temperature |
林冠郁闭度/% Canopy closure |
林冠高度/m Canopy height |
参考文献 References |
| 玉龙雪山 Yulong Mountain |
亚高山 | 针叶林 | 935 | 12.79 | 85—90 | 30—40 | [9-12] |
| 哀牢山 Ailao Mountain |
亚热带 | 中山湿性常绿 阔叶林 |
1931 | 11 | 89—95 | 25—30 | [12-15] |
| 西双版纳 Xishuangbanna |
热带 | 热带季节 雨林、季风常 绿阔叶林等 |
1493 | 21.8 | 80—85 | 45—60 | [12, 16] |
热带样地处于低山沟谷及低丘上, 主要的森林类型为热带季节雨林、热带季雨林和热带山地常绿阔叶林等。原始林的林冠高达50—60 m, 由望天树(Parashorea chinensis)占优势, 中下层乔木主要包括毛猴欢喜(Sloanea tomentosa)、绒毛番龙眼(Pometia pinnata)、假海桐(Pittosporopsis kerrii)、云树(Garcinia cowa)、蚁花(Mezzettiopsis creaghii)、染木(Saprosma ternata)等, 次生林主要由壳斗科(Fagaceae)植物组成[16]。亚热带样地分布于滇中高原南、北两侧的几条大山脉的中山地带, 植被类型主要为中山湿性常绿阔叶林, 优势植物包括壳斗科(Fagaceae)、木兰科(Magnoliaceae)、樟科(Lauraceae)、山茶科(Theaceae)等[15]。亚高山样地位于横断山脉地区, 由下而上分别为云南松(Pinus yunnanensis)林、丽江云杉(Picea likiangensis)林、大果红杉(Larix potaninii var. australis)林和冷杉(Abies fabri)林[10-11]。热带1200m及1400m的样地在20世纪60年代曾被间伐, 其他样地较少受人类干扰[12]。
1.2 土壤动物采集与鉴定2017年10—11月(雨季末期)在云南省3个典型气候带下的48个海拔样地内取样, 每个样地随机选择3个0.6 m × 0.6 m的小样方, 抓取凋落物层于布袋中, 做好标签, 三处合并为一个混合样。在实验室中用Berlese-Tullgren法(干漏斗法)烘48 h后获得土壤动物。将分离的土壤动物保存在75%酒精中, 在解剖镜下依照《中国土壤动物检索图鉴》、《中国亚热带土壤动物》、《中国土壤动物》等书籍资料和 http://www.collembola.org/ 等分类网站进行弹尾类的分类鉴定(属)并计数[17-19]。
1.3 环境数据采集采集的环境数据包括(1)温度:在样地内布置的土壤温度自动记录设备(iButton纽扣式温度计), 获得每小时的样地土壤温度。(2)水分:随机选择3个小样方, 分别用环刀取100 cm3的土壤放入铝盒, 使用烘箱烘干法(105℃)测定土壤含水量。(3)pH:使用pH测量仪测定。(4)凋落物产量:测量凋落物厚度。(5)土壤容重(容重法)和土壤孔隙度。
1.4 数据统计分析类群数量等级划分:个体数量占总捕获量的10%以上者为优势类群;1%—10%者为常见类群;1%以下者为稀有类群[18]。
弹尾类密度:
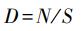
|
其中N为采样点弹尾类所有的个体数之和, S为所取凋落物的体积。
弹尾类丰富度采用以下两个指数计算:
(1) Margalef′s index (D)[20]:

|
(2) Menhinick′s index (D)[21]:
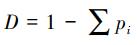
|
多样性采用以下两个指数计算:
(1) Shannon-Wiener index (H′)[22]:
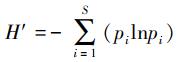
|
(2) Simpson index (D)[23]:
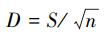
|
均匀度计算指数:
Pielou evenness index (JSW)[24]:
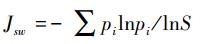
|

|
其中, S为采样点弹尾类共有属数目;n为采样点弹尾类所有的个体数之和;ni为采样点弹尾类属i的个体数;pi为第i属的比例多度。
多样性指数使用R软件中的“vegan”包计算。
环境因子的筛选:使用最小二乘回归法对弹尾类多样性的海拔格局进行回归分析, 通过比较一次与二次项回归的模型AIC值与P值判定海拔分布格局。以多样性为响应变量, 以温度(雨季土壤平均温度、最高温度、最低温度)、凋落物产量、pH、土壤含水量、土壤容重以及土壤孔隙度为解释变量, 通过全模型子集回归筛选最佳环境模型(AIC值最小)。同时分析ΔAIC < 2的模型总数与出现在这些模型中的参数频次, 进行模型平均分析。使用R软件进行环境因子分析, 主要使用“MuMIn”、“car”和“QuantPsyc”包等。使用Sigmaplot 12.0软件绘制弹尾类多样性海拔分布格局。
2 结果与分析 2.1 不同气候带弹尾类组成概况本研究采集到凋落物中的弹尾类共19150只, 隶属于10科29属。在科的分类水平上看, 如图 1所示, 等节䖴科数量最多(59.9%), 其次为棘䖴科(21.8%)和球角䖴科(10.1%), 这3个科合计占总体的91.8%。在属的分类水平上看, 如图 1所示, 其中符䖴属(39.9%, 等节䖴科)数量最多, 其余优势属为棘䖴属(21.7%, 棘䖴科)和球角䖴属(10.1%, 球角䖴科), 这3个属合计占总体的71.7%。

|
| 图 1 不同科(属)弹尾类占总体的百分比 Fig. 1 Percentage of Collembola in different families (genera) |
就弹尾类密度而言, 如图 2所示, 热带雨林和亚热带常绿阔叶林捕获的弹尾类密度相差不多, 在亚高山针叶林有明显增加。就弹尾类的属数多少而言, 如图 2所示, 由多到少依次为热带雨林、亚高山针叶林、亚热带常绿阔叶林。

|
| 图 2 不同气候带弹尾类的密度与属数 Fig. 2 Density and genus numbers of Collembola in different climatic zones |
如图 3所示, 土壤弹尾类的Margalef′s指数(R2=0.2996, P<0.0001)、Menhinick′s指数(R2=0.5140, P<0.0001)、Shannon-Wiener多样性指数(R2=0.5268, P<0.0001)、Simpson多样性指数(R2=0.1738, P<0.01)和Pielou均匀度指数(R2=0.1034, P<0.05)随海拔及纬度上升呈现单调递减格局, 整体呈现为热带雨林最高、亚热带常绿阔叶林居中、亚高山针叶林最低。

|
| 图 3 弹尾类多样性沿海拔分布格局 Fig. 3 Collembola diversity distribution patterns along elevation in different climatic zones |
以多样性为响应变量, 以温度、凋落物产量、pH、土壤含水量、土壤容重以及土壤孔隙度为解释变量, 通过全模型子集回归筛选最佳环境模型(AIC值最小)可以看出(表 2), 弹尾类Margalef′s指数与凋落厚度负相关(Coefficient=-0.1381, P=0.0210), 与雨季极高温度正相关(Coefficient=0.1143, P=0.0007), 与土壤容重微弱负相关(Coefficient=-1.1021, P=0.0971), 与土壤含水量微弱正相关(Coefficient=1.4415, P=0.0877);弹尾类Menhinick′s指数与土壤容重负相关(Coefficient=-2.0223, P=0.0000), 与凋落物厚度负相关(Coefficient=-0.0925, P=0.0026), 与土壤孔隙度负相关(Coefficient=-0.0213, P=0.0228), 与雨季平均温度正相关(Coefficient=0.0875, P=0.0000);弹尾类Shannon-Wiener多样性指数与雨季极低温度正相关(Coefficient=0.0326, P=0.0002), 与雨季平均温度呈正相关(P=0.0358);弹尾类Pielou均匀度指数与雨季极低温度正相关(Coefficient=0.0061, P=0.0484)。
| 响应变量 Response variable |
调整 R2 Adj.R2 |
最佳模型中的 环境变量 Environmental variables in best models |
模型平均估计 Model- averaged estimate |
标准误 Std.Error |
P 值 Pr(>|z|) |
变量相对 重要值 Relative variable importance |
该变量出现的 最佳模型数量 N containing models |
最佳模型个数 Number of best models |
|
| 玛格列夫指数 |
0.5836 | Litter amount# | -0.1381 | 0.0588 | 0.0210 | * | 1 | 7 | 7 |
| T-maxR# | 0.1143 | 0.0331 | 0.0007 | *** | 0.91 | 6 | |||
| Soil bulk density# | -1.1021 | 0.6480 | 0.0971 | . | 0.43 | 3 | |||
| Soil water content | 1.4415 | 0.8249 | 0.0877 | . | 0.4 | 3 | |||
| T-minR | 0.0691 | 0.0533 | 0.2018 | 0.28 | 3 | ||||
| Soil porosity | -0.0184 | 0.0159 | 0.2608 | 0.12 | 1 | ||||
| 门希尼克指数 Menhinick |
0.6444 | Soil bulk density# | -2.0223 | 0.4719 | 0.0000 | *** | 1 | 2 | 2 |
| Litter amount# | -0.0925 | 0.0298 | 0.0026 | ** | 1 | 2 | |||
| Soil porosity# | -0.0213 | 0.0091 | 0.0228 | * | 1 | 2 | |||
| T-rain# | 0.0875 | 0.0115 | 0.0000 | *** | 1 | 2 | |||
| Soil water content | -0.4941 | 0.6022 | 0.4255 | 0.27 | 1 | ||||
| 香农-维纳指数 Shannon-Wiener |
0.2968 | T-minR# | 0.0326 | 0.0085 | 0.0002 | *** | 0.6 | 3 | 5 |
| T-rain | 0.0358 | 0.0080 | 0.0000 | *** | 0.27 | 1 | |||
| Litter amount | -0.0357 | 0.0189 | 0.0662 | . | 0.13 | 1 | |||
| T-maxR | 0.0371 | 0.0087 | 0.0000 | *** | 0.13 | 1 | |||
| Soil water content | 0.2448 | 0.3912 | 0.5425 | 0.13 | 1 | ||||
| Bulk density | -0.1514 | 0.2464 | 0.5500 | 0.13 | 1 | ||||
| 辛普森指数 Simpson |
0.1697 | T-minR# | 0.0122 | 0.0138 | 0.3830 | 0.72 | 3 | 4 | |
| T-rain | -0.0039 | 0.0224 | 0.8620 | 0.46 | 2 | ||||
| pH | 0.0132 | 0.0209 | 0.5390 | 0.15 | 1 | ||||
| 皮埃卢指数 Pielou evenness |
0.07213 | T-rain | 0.0100 | 0.0084 | 0.2392 | 0.54 | 4 | 7 | |
| Soil bulk density | -0.1470 | 0.1222 | 0.2388 | 0.44 | 4 | ||||
| T-minR# | 0.0061 | 0.0030 | 0.0484 | * | 0.33 | 2 | |||
| T-maxR | -0.0020 | 0.0116 | 0.8644 | 0.22 | 2 | ||||
| Soil water content | -0.2755 | 0.2266 | 0.2371 | 0.09 | 1 | ||||
| #:进入AIC值最小模型的解释变量; N containing models:该解释变量出现在最佳模型(ΔAIC < 2)的数量; Number of best models:最佳模型的总个数(ΔAIC < 2);Litter amount:凋落物厚度;T-maxR:雨季土壤最高温度;T-minR:雨季土壤最低温度;T-rain:雨季土壤平均温度;Soil bulk density:土壤容重;Soil water content:土壤含水量;Soil porosity:土壤孔隙度;pH:土壤酸碱度; Signif. codes: 0 ‘***’ 0.001 ‘**’0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1 | |||||||||
3个典型气候带下弹尾类属的数量排序为:热带雨林>亚高山针叶林>亚热带常绿阔叶林, 但3个气候带属数相差很小且绝对优势类群均为符䖴属(等节䖴科), 表明弹尾类组成在属级分类单位上具有一定的跨气候带稳定性, 这与前人的研究结果基本一致[5, 7, 25]。3个典型气候带下弹尾类密度排序为:亚高山针叶林>热带雨林>亚热带常绿阔叶林, 热带雨林和亚热带常绿阔叶林凋落物层弹尾类的密度相差较小, 亚高山针叶林则明显高于另外两个气候带, 这主要是由于符䖴属(占亚高山总体捕获量46.67%)和棘䖴属(占亚高山针叶林整体捕获量28.26%)的个体数量急剧增加引起的。符䖴属在所有气候带均为绝对优势属, 但在亚高山针叶林的数量与占比更高;棘䖴属在热带雨林和亚热带常绿阔叶林仅占1.11%和2.07%, 但在亚高山针叶林骤增至28.26%, 使得弹尾类的密度在亚高山针叶林达到最高。在日本亚高山针叶林凋落物层开展的相关研究也发现符䖴属(33.5%)与棘䖴属(15.9%)在弹尾目捕获的总数量占据绝对优势地位[26]。据此推测, 亚高山针叶林的凋落物层提供的生境可能更有利于符䖴属和棘䖴属内的种群生长, 未来需要更多的对比研究以探索各典型气候带不同植被类型下凋落物层弹尾类的优势种与稀有种的多度分布格局。
结果显示弹尾类的多样性呈现热带雨林最高、亚热带常绿阔叶林居中、亚高山针叶林最低的分布格局, 这与该地区幼苗物种丰富度[27]和蛾类物种丰富度沿海拔分布格局相同[28]。可能是由于这3个气候带最显著的差异为温度, 而影响弹尾类、蛾类和幼苗多样性分布格局的主要环境因子也为温度, 温度的变化规律相同使得物种丰富度形成了相似的海拔分布格局。本研究得到的弹尾类多样性在亚热带高于亚高山的结果与在澳大利亚莱明顿国家公园得到的弹尾类多样性在亚热带高于寒温带[4]的结果相一致, 而得到的弹尾类多样性沿海拔呈单调下降格局的结果与贡嘎山东坡[5]和法国的马克拉普拉涅(Macot La Plagne)[6]得到的结果相反。这可能是由于在澳大利亚和法国的实验中, 不同海拔的弹尾类的群落组成与土壤的酸碱度相关性较强, 弹尾类的密度和多样性与土壤的pH呈负相关, 而本实验弹尾类多样性沿海拔分布格局与温度相关性最强, 与土壤酸碱性的相关性则很微弱。
在本研究中, 通过全模型子集回归筛选最佳环境模型得出, 影响弹尾类的Margalef′s丰富度指数、Menhinick′s丰富度指数、Shannon-Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数的环境因子中, 均有雨季土壤极高温度、雨季土壤极低温度和雨季土壤平均温度中的一个, 且均为正相关。温度是驱动土壤动物群落结构与多样性的重要环境因子。就直接影响而言, 温度作为土壤动物生存的必须环境因子, 可通过影响土壤动物体内酶的活性影响其生长发育, 而且在一定范围内提高土壤动物的活性[29]。间接影响而言, 温度可以通过影响地上植物凋落物的种类、质量和数量来影响土壤动物的多样性分布格局[30]。温度可以影响土壤动物对凋落物的分解, 使土壤动物对分解的贡献率随海拔升高而逐渐降低[31], 本研究中亚高山的植被类型为针叶林, 产生的凋落物种类较单一, 使得弹尾类的丰富度指数(Margalef′s指数、Menhinick′s指数)在3个气候带中最低。而优势物种数量巨大, 这使得弹尾类的多样性指数(Shannon-Wiener多样性指数、Simpson多样性指数)和均匀度指数(Pielou均匀度指数)也达到了最低。本研究的结果进一步证实了温度与弹尾类多样性呈正相关关系。
本研究主要关注雨季末期不同气候带土壤弹尾类多样性沿海拔分布格局, 雨季的湿度和温度都较为适宜弹尾类的生存, 环境胁迫较少, 而在旱季随着水热条件的改变, 其分布格局可能有所不同, 有待进一步研究。由于捕获的弹尾类数量较多, 通过形态分析将弹尾类鉴定到种的难度巨大, 本研究在属一级分类水平探讨了弹尾类多样性在不同气候带下的分布格局。土壤动物多样性巨大且分类信息大量缺失, 土壤动物分布格局研究多基于较高的分类单元(科、属或形态种)开展[32], 如在属级分类精度上的线虫多样性研究[33], 以及在科级水平上开展的多类群土壤动物多样性研究[34-36]。最近的研究表明, 大多数同属土壤动物在土壤食物网中的营养级位置相同[37], 而在科一级分类水平的信息可更好地反映弹尾类的生境选择偏好[38]。未来的研究需要结合分子生物学手段与传统的形态鉴定方法, 在更高的分类精度上探索弹尾类及其他土壤动物类群的物种多样性分布格局及其潜在的环境影响机制。
4 结论在云南省热带、亚热带和亚高山这3个典型气候带下的森林凋落物层中, 弹尾类绝对优势类群均为符䖴属。从总捕获量上看, 符䖴属占39.9%(等节䖴科), 其余优势属为棘䖴属(21.7%, 棘䖴科)和球角䖴属(10.1%, 球角䖴科)。弹尾类的丰富度指数(Margalef′s指数、Menhinick′s指数)、多样性指数(Shannon-Wiener多样性指数、Simpson多样性指数)和均匀度指数(Pielou均匀度指数)均沿热带雨林、亚热带常绿阔叶林、亚高山针叶林气候带呈单调递减格局。温度是影响弹尾类多样性沿不同气候带分布格局的主要环境因子。
| 䖴科名 Family |
䖴科拉丁名 Family Latin Name |
䖴属名 Genera |
䖴属拉丁名 Genera Latin Name |
| 长角䖴科 | Entomobryidae | 刺齿䖴属 | Homidia |
| 鳞长䖴属 | Lepidocyrtus | ||
| 裸长角䖴属 | Sinella | ||
| 球角䖴科 | Hypogastruridae | 球角䖴属 | Hypogastrura |
| 等节䖴科 | Isotomidae | 隐䖴属 | Cryptopygus |
| 符䖴属 | Folsomia | ||
| 裔符䖴属 | Folsomides | ||
| 类符䖴属 | Folsomina | ||
| 似等䖴属 | Isotomodes | ||
| 陷等䖴属 | Isotomurus | ||
| 近缺䖴属 | Paranurophorus | ||
| 拟缺䖴属 | Pseudanurophorus | ||
| 二刺䖴属 | Uzelia | ||
| 短角䖴科 | Neelidae | 短角䖴属 | Neelus |
| 疣䖴科 | Neanuridae | 疣䖴属 | Neanura |
| 副䖴属 | Paranura | ||
| 伪亚䖴属 | Pseudachorutes | ||
| 沃䖴属 | Womersleya | ||
| 棘䖴科 | Onychiuridae | 棘䖴属 | Onychiurus |
| 土䖴属 | Tullbergia | ||
| 长角长䖴科 | Orchesellidae | 长角长䖴属 | Orchesellides |
| 爪䖴科 | Paronellidae | 盐长䖴属 | Salina |
| 圆䖴科 | Sminthuridae | 齿棘圆䖴属 | Arrhopalites |
| 异圆䖴属 | Heterosminthurus | ||
| 小圆䖴属 | Sminthurinus | ||
| 圆䖴属 | Sminthurus | ||
| 球圆䖴属 | Sphaeridia | ||
| 环角圆䖴属 | Ptenothrix | ||
| 鳞䖴科 | Tomoceridae | 鳞䖴属 | Tomocerus |
| [1] |
Christiansen K A. Springtails. Kansas: Emporia State University, 2003. |
| [2] |
陈建秀, 麻智春, 严海娟, 张峰. 跳虫在土壤生态系统中的作用. 生物多样性, 2007, 15(2): 154-161. DOI:10.3321/j.issn:1005-0094.2007.02.006 |
| [3] |
Yang X D, Yang Z, Warren M W, Chen J. Mechanical fragmentation enhances the contribution of Collembola to leaf litter decomposition. European Journal of Soil Biology, 2012, 53: 23-31. DOI:10.1016/j.ejsobi.2012.07.006 |
| [4] |
Greenslade P, Kitching R L. Potential effects of climatic warming on the distribution of Collembola along an altitudinal transect in Lamington National Park, Queensland, Australia. Memoirs of the Queensland Museum, 2011, 55(2): 333-347. |
| [5] |
王群, 武崎, 刘兴良, 张荣芝, 吴鹏飞. 海拔高度大尺度变化对土壤跳虫群落的影响. 西南农业学报, 2016, 29(8): 1936-1942. |
| [6] |
Loranger G, Bandyopadhyaya I, Razaka B, Ponge J F. Does soil acidity explain altitudinal sequences in collembolan communities?. Soil Biology and Biochemistry, 2001, 33(3): 381-393. DOI:10.1016/S0038-0717(00)00153-X |
| [7] |
王宗英, 朱永恒, 路有成, 聂献忠, 陈建秀, 孟文新, 黄诚. 九华山土壤跳虫的生态分布. 生态学报, 2001, 21(7): 1142-1147. DOI:10.3321/j.issn:1000-0933.2001.07.016 |
| [8] |
Sadaka N, Ponge J F. Soil animal communities in holm oak forests: influence of horizon, altitude and year. European Journal of Soil Biology, 2003, 39(4): 197-207. DOI:10.1016/j.ejsobi.2003.06.001 |
| [9] |
薛润光, 杨少华. 丽江玉龙雪山自然保护区珍稀濒危植物资源的初步研究. 林业调查规划, 2006, 31(1): 50-53. DOI:10.3969/j.issn.1671-3168.2006.01.014 |
| [10] |
冯建孟, 王襄平, 徐成东, 杨元合, 方精云. 玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局. 山地学报, 2006, 24(1): 110-116. DOI:10.3969/j.issn.1008-2786.2006.01.016 |
| [11] |
《云南植被》编写组. 云南植被. 北京: 科学出版社, 1987.
|
| [12] |
Song L, Ma W Z, Yao Y L, Liu W Y, Li S, Chen K, Lu H Z, Cao M, Sun Z H, Tan Z H, Nakamura A. Bole bryophyte diversity and distribution patterns along three altitudinal gradients in Yunnan, China. Journal of Vegetation Science, 2015, 26(3): 576-587. DOI:10.1111/jvs.12263 |
| [13] |
Liu W Y, Fox J E D, Xu Z F. Leaf litter decomposition of canopy trees, bamboo and moss in a montane moist evergreen broad-leaved forest on Ailao Mountain, Yunnan, south-west China. Ecological Research, 2000, 15(4): 435-447. |
| [14] |
杨国平, 郑征, 张一平, 刘玉洪, 巩合德, 鲁志云. 哀牢山中山湿性常绿阔叶林群落生态学特征. 东北林业大学学报, 2010, 38(9): 16-19. DOI:10.3969/j.issn.1000-5382.2010.09.006 |
| [15] |
李洁琼, 宋晓阳, 曹敏. 云南哀牢山和玉龙雪山森林树种幼苗对海拔梯度的响应及其季节性差异. 应用生态学报, 2016, 27(11): 3403-3412. |
| [16] |
兰国玉, 胡跃华, 曹敏, 朱华, 王洪, 周仕顺, 邓晓保, 崔景云, 黄建国, 刘林云, 许海龙, 宋军平, 何有才. 西双版纳热带森林动态监测样地——树种组成与空间分布格局. 植物生态学报, 2008, 32(2): 287-298. DOI:10.3773/j.issn.1005-264x.2008.02.006 |
| [17] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [18] |
尹文英. 中国亚热带土壤动物. 北京: 科学出版社, 1992.
|
| [19] |
尹文英. 中国土壤动物. 北京: 科学出版社, 2000.
|
| [20] |
Margalef R. Perspectives in Ecological Theory. Chicago: University of Chicago Press, 1968.
|
| [21] |
Menhinick E F. A comparison of some species‐individuals diversity indices applied to samples of field insects. Ecology, 1964, 45(4): 859-861. |
| [22] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: The University of Illinois Press, 1949. |
| [23] |
Simpson E H. Measurement of diversity. Nature, 1949, 163(4148): 688-688. DOI:10.1038/163688a0 |
| [24] |
Pielou E C. The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 1966, 13: 131-144. DOI:10.1016/0022-5193(66)90013-0 |
| [25] |
何振, 赵琴, 李迪强, 李密. 八大公山土壤跳虫群落结构与多样性的季节动态. 林业科学研究, 2018, 31(3): 68-77. |
| [26] |
Niijima K. Collembola of the subalpine coniferous forest of Kita-Yatsugatake volcano. Kontyû, 1966, 34(3): 244-247. |
| [27] |
宋晓阳.云南典型森林林下幼苗的时空动态特征研究[D].北京: 中国科学院大学, 2017.
|
| [28] |
Ashton L A, Nakamura A, Burwell C J, Tang Y, Cao M, Whitaker T, Sun Z, Huang H, Kitching R L. Elevational sensitivity in an Asian 'hotspot': Moth diversity across elevational gradients in tropical, sub-tropical and sub-alpine China. Scientific Reports, 2016, 6: 26513. DOI:10.1038/srep26513 |
| [29] |
陈泮勤. 地球系统碳循环. 北京: 科学出版社, 2004.
|
| [30] |
林英华, 孙家宝, 张夫道. 我国重要森林群落凋落物层土壤动物群落生态特征. 生态学报, 2009, 29(6): 2938-2944. DOI:10.3321/j.issn:1000-0933.2009.06.021 |
| [31] |
王邵军.武夷山不同海拔土壤动物对凋落物分解的影响〖D〗.南京: 南京林业大学, 2009.
|
| [32] |
Decaëns T. Macroecological patterns in soil communities. Global Ecology and Biogeography, 2010, 19(3): 287-302. DOI:10.1111/j.1466-8238.2009.00517.x |
| [33] |
Viketoft M, Sohlenius B, Boström S, Palmborg C, Bengtsson J, Berg M P, Huss-Danell K. Temporal dynamics of soil nematode communities in a grassland plant diversity experiment. Soil Biology and Biochemistry, 2011, 43(5): 1063-1070. DOI:10.1016/j.soilbio.2011.01.027 |
| [34] |
Doblas-Miranda E, Wardle D A, Peltzer D A, Yeates G W. Changes in the community structure and diversity of soil invertebrates across the Franz Josef Glacier chronosequence. Soil Biology and Biochemistry, 2008, 40(5): 1069-1081. DOI:10.1016/j.soilbio.2007.11.026 |
| [35] |
Xu G. R, Lin Y H., Zhang S., Zhang Y X., Li G X., Ma K M.. Shifting mechanisms of elevationaldiversity and biomass patterns in soil invertebrates at treeline. Soil Biology and Biochemistry, 2017, 113,: 80-88. DOI:10.1016/j.soilbio.2017.05.012 |
| [36] |
Xu G. R, Zhang S., Zhang Y X., Ma K M. Environmental correlates underlying elevational richness, abundance, and biomass patterns of multi-feeding guilds in litter invertebrates across the treeline. Science of The Total Environment, 2018, 633: 529-538. DOI:10.1016/j.scitotenv.2018.03.212 |
| [37] |
Potapov A M, Scheu S, Tiunov A V. Trophic consistency of supraspecific taxa in below-ground invertebrate communities: Comparison across lineages and taxonomic ranks. Functional Ecology, 2019, 33(6): 1172-1183. |
| [38] |
Ponge J F, Salmon S. Spatial and taxonomic correlates of species and species trait assemblages in soil invertebrate communities. Pedobiologia, 2013, 56(3): 129-136. DOI:10.1016/j.pedobi.2013.02.001 |