 2020, Vol. 40
2020, Vol. 40文章信息
- 郑芬, 李兆佳, 邱治军, 赵厚本, 周光益
- ZHENG Fen, LI Zhaojia, QIU Zhijun, ZHAO Houben, ZHOU Guangyi
- 广东南岭天然常绿阔叶林林下光环境对林下幼树功能性状的影响
- Effects of understory light on functional traits of evergreen broad-leaved forest saplings in Nanling Mountains, Guangdong Province
- 生态学报. 2020, 40(13): 4516-4527
- Acta Ecologica Sinica. 2020, 40(13): 4516-4527
- http://dx.doi.org/10.5846/stxb201911152453
-
文章历史
- 收稿日期: 2019-11-15
- 修订日期: 2020-05-09





2. 南京林业大学, 南京 210037;
3. 南岭北江源森林生态系统国家定位观测研究站, 广州 510520
2. Nanjing Forestry University, Nanjing 210037, China;
3. Beijiangyuan National Forest Ecosystem Research Station, Nanling Mts. China, Guangzhou 510520, China
森林冠层是森林与大气环境接触最直接、最活跃的界面, 对森林生态系统内环境因子如光照、温度、水分和土壤养分等有着重要的影响[1-2]。其中光照是影响林下幼树生长、存活以及更新的关键生态因子之一[3-6], 冠层通过对日光的吸收、透射和反射改变林下光照的强弱和分布, 对林下幼树的生长、生物量分配及更新具有重要作用[7-9]。
目前针对冠层结构与林下光环境特征之间关系的研究相对较多[10-13], 国内的大量研究是通过半球摄影技术获得的数据来分析冠层结构与林下直射光、散射光及总光照强度之间的关系[14-15], 但此种方式不能获取林下光照质量特征。而光照强度和光照质量的差别都显著影响林下幼树的生理特征[16]。过去的研究表明:透光率与R/FR之间的关系呈正相关[7, 17-19], 且可以通过双曲线模型用透光率来预测光质。但这些研究都集中在人工纯林或者成分简单的混交林中[20-21], 关于天然林的研究较少。天然林林下光质是否可以由透光率预测?透光率对光质的拟合解释强度为多少?
光环境对植物的生长、竞争、形态和光合生理可塑性等有着重要的影响[6], 研究植物叶片在不同光环境下的响应是探究植物对光资源的利用与分配的有效途径[22]。比叶面积(Specific leaf area, SLA)和叶片氮含量(Leaf nitrogen concentration, LNC)作为两个关键功能性状, 分别表征了叶片对光能的捕获和CO2同化的能力[23]。在光照强度较弱的环境中植物通过增加SLA来获取足够的光资源, 同时增加LNC来提高净光合速率[24]。光线透过森林冠层后, 伴随着光照强度的减弱, 光谱组成也发生了变化。光质对植物的影响作用在栽培与农业领域中应用广泛, 红光和蓝光可以促进幼苗的生长, 提高农作物的产量[25-28]。在较低的光照强度下, 蓝光部分占比的增多能促进植物的光合作用, 混交林林下Bw/Rw的增加可能使得林下幼树单位叶面积光合速率更高[21]。远红光能促进植物茎伸长率的增加, 在对桃树冠层的研究中发现较低的R/FR能减少分支, 促进茎的伸长生长[29]。然而, 在天然林中, 不同林龄梯度林下幼树功能性状及光合特征对光强的变化有何响应?天然林群落R/FR和Bw/Rw的变化对林下幼树光合作用与形态特征是否有影响?
因此, 本研究以广东南岭区域3种林龄(幼龄林, 中龄林, 老龄林)的常绿阔叶林群落为研究对象, 在野外调查的基础上, 选择冠层结构、林下光环境和林下幼树功能性状相关指标, 研究林龄梯度引起的亚热带常绿阔叶林冠层结构与林下光环境特征指标差异, 分析幼树对光环境差异的响应。以期了解林下幼树功能性状等生态学特征、解析林下光环境对林下幼苗幼树更新的影响机制, 从而为森林保护和合理经营提供可靠的理论依据。
1 试验地概况试验样地位于广东省韶关市曲江区小坑镇周边, 地理位置为E112°30′—113°04′, N24°37′—24°57′。属中亚热带湿润性季风气候, 一年四季受季风影响, 冬季盛行东北季风, 夏季盛行西南和东南季风。四季特点为春季阴雨连绵, 秋季降水偏少, 冬季寒冷, 夏季偏热。年平均气温18.8—21.6℃, 年降雨量1400—2400mm, 年日照时数1473—1925h, 地貌以中低山为主, 坡度为15°—40°。该区的地带性植被类型为中亚热带常绿阔叶林。植被种类丰富, 层次结构复杂。
2 研究方法 2.1 样地设置与调查2018年7月在南岭韶关曲江区小坑镇周边, 选择海拔、坡度、坡向、土壤和水分条件相近的天然常绿阔叶林群落, 以最大限度降低立地条件不同引起的差异, 建立3个林龄的森林群落样地, 构成一个林龄梯度的样地系列(幼龄林、中龄林和老龄林)。其中幼龄林林龄约30年, 为1988年左右采伐后的天然次生林, 中龄林林龄约60年, 为20世纪50年代末修水库被破坏后形成的天然次生林, 老龄林是近原始林, 为20世纪50年代采伐时特意留下的一片约23hm2的原始林林地。3个群落土壤均为花岗岩发育的红壤。利用全站仪构建3个大小为40m×40m的样地, 每个样地内划分成10m×10m的小样方16个, 并在每个样方的四角及中心点用PVC管标记。调查林下木本植物(乔木、灌木)的胸径、树高等因子, 同时记录地理坐标、海拔、坡度、坡向等参数。样地概况见表 1。
| 林分 Forest type | 林龄 Forest age/a | 地理坐标 Geographic coordinates | 海拔 Altitude/m | 坡度 Slope/(°) | 坡向 Aspect | 平均胸径 Average DBH/cm | 株密度 Stand density/ (株/m2) |
| YF | 30 | N24°39′11.7″, E113°52′35.7″ | 550 | 32 | 东北偏北 | 7.32 | 0.57±0.044a |
| MF | 50—60 | N24°42′28.6″, E113°50′08.0″ | 250 | 30 | 东北偏北 | 9.45 | 0.18±0.019b |
| OF | >100 | N24°42′01.7″, E113°54′28.8″ | 480 | 34 | 西北偏北 | 6.35 | 0.46±0.061a |
| YF, 幼龄林young forest; MF, 中龄林middle-aged forest; OF, 老龄林old-growth forest; 不同小写字母代表差异显著P < 0.05;每个值代表平均值±标准误 | |||||||
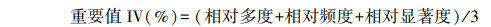
|
式中, 相对多度(%)=某一树种个体数/所有树种个体总数×100;相对频度(%)=某一树种频度/所有树种频度总和×100;相对显著度(%)=某一树种胸径断面积之和/所有树种胸径断面积之和×100
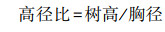
|
于2018年7月至2019年7月每个月定期采集冠层结构和光环境数据, 平均后用于分析处理。1)冠层结构指标的获取:拍摄时间选在阴天或者无风的8:00—10:00, 以保证光照条件一致和使直射光造成的眩光最小化[30], 在3个林龄梯度天然常绿阔叶林固定样地固定样方中心处用Nikon D700数码相机配合鱼眼镜头获取半球面林冠影像。拍摄时具体操作为:用指南针确定正南方并面朝指针所指方向, 在确保相机的视域内无人的情况下, 镜头朝上进行拍摄;每个拍摄点每次拍摄3张照片, 同一样点重复取样时保持原位, 每个样地获得48张照片, 共获得3个样地144张照片。2)光环境指标的获取:每个月尽量选择一个典型晴天和一个典型阴天分别对3个林龄梯度常绿阔叶林群落样地进行测量, 根据Fielder和Comeau的文献[31]中所述方法, 将商用的PAR探头(NHGH09光合有效辐射传感器, 中国武汉能慧科技有限公司)连接到万用表上测量冠层下方光合有效辐射数据;采用Spectro-Sense2传感器连接光谱探头获得所需要的红光(R)、远红光(FR)、宽带红光(Rw)和宽带蓝光(Bw)的数据。测量时将PAR探头和光谱探头固定在同一支架上, 将支架升高到2m, 固定仪器与地面垂直, 避开光斑和树叶遮挡进行测量。由于光谱探头所测的光照波长仅代表PAR的一小部分, 在光照强度水平较低的时候会低于传感器测量下限[32], 因此实测时间为每天的9:00—16:00。老龄林冠层上方光照数据来自林内高于林冠层10m的气象梯度塔, 幼龄林和中龄林在林外空地测量作为对照。

|
叶片采集:于2018年9月采样, 在每个样地中选择个体较大且长势良好的挂牌标记幼树植株。
室内分析:采集回来的枝条平铺在地面上, 旁边放一张A4纸作为对照, 用照相机拍照并做好编号。选取枝条上15片左右完全展开、无病虫害、成熟且完整的叶片, 用Epson扫描仪扫描植物叶片后分别放入信封袋内保存。使用Digimizer version 4.3.0软件分析得到的扫描图像, 得到枝条面积、遮挡面积和叶面积。将扫描后的叶片置于85℃的烘箱中烘干至恒重, 然后使用分析天平称量干重, 换算得到比叶面积(SLA)(cm2/g)。最后将叶片送中国林业科学研究院热带林业研究所分析室检测叶片氮含量(LNC)(g/kg)。(采用凯氏定氮法[33]测定全氮含量)
1) 比叶面积=单片叶面积/单片干重,
2) 重叠率=1-枝条面积/叶片总面积,
3) 空隙率=1-枝条面积/遮挡面积。
2.5 野外光响应曲线及指标的测定2019年4月至5月, 晴天天气下在8:00—11:30间分别在3个样地中测定。根据植物对光照的需求不同, 选择3个林分共有树种(演替前期种-少花桂(Cinnamomum pauciflorum), 演替后期种-锥(Castanopsis chinensis), 演替中期种-木荷(Schima superba)[34-36]), 每个种选择3株, 每株选择中等大小、叶色正常、完全舒展并且成熟的3片叶子测定。使用Li-6400便携式光和系统(Li-Cor, lnc, 美国)测定叶片的净光合速率(Pn), 使用开放气路, 空气流速为0.5 L/min, 温度为25℃, 相对湿度为50%, CO2浓度为360μmol/mol。测定梯度设定为1800、1200、1000、800、600、400、300、200、150、100、90、80、70、60、50、40、30、20、0μmol m-2 s-1, 测定时每一光强下停留3 min。测定前在600—1200μmol m-2 s-1光强(依植物种类及生长光强而定)下诱导30 min, 以光合有效辐射(PAR)为横轴, Pn为纵轴绘出光合作用光响应曲线(Pn-PAR曲线), 依据叶子飘[37]的方法拟合Pn-PAR的曲线方程:
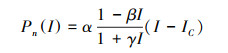
|
式中, Pn(I)为净光合速率;I为光合有效辐射;Ic为光补偿点;α、β、γ是独立于I的系数。
暗呼吸速率(Rd):
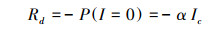
|
光饱和点(Im):
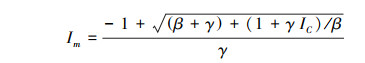
|
最大净光合速率P(Im):
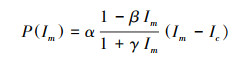
|
所拍摄的林冠照片采用Gap Light Analyzer version 2.0软件进行处理, 分析时将每张照片所在样方的经纬度坐标及海拔高程作为样地参数输入设置。冠层影像分析后获得一系列的冠层结构和林下光照参数, 包括林冠开度(Canopy Openness, CO)和叶面积指数(Leaf Area Index, LAI)。
利用Microsoft Excel 2016进行数据初处理, 采用IBM SPSS Statistics 20.0软件单因素方差分析(one-way ANOVA)和多重比较。对于方差不齐的数据, 采用Welch′s ANOVA分析, 结果比较选用Games-Howell。利用光合助手version 4.1.1软件[37]计算光合特征参数, originPro 9.0软件作图。
3 结果与分析 3.1 不同林龄群落特征调查结果表明, 不同林龄群落物种组成表现出一定差异。3个样地共出现木本植物83种, 分属31科50属, 株密度调查结果(表 1)显示, 幼龄林和老龄林密度没有显著差异, 但是中龄林株密度显著低于二者(P < 0.05)。根据树种的多度、频度和显著度, 计算得到各主要物种的重要值(表 2), 进而确定群落建群种均为壳斗科锥属物种, 分别为黧蒴锥(Castanopsis fissa)、锥(Castanopsis chinensis)和罗浮锥(Castanopsis fabri)。幼龄林乔木层植物20科23属52种, 灌木层植物21科33属62种, 中龄林群落乔木层植物17科28属42种, 灌木层植物21科35属51种, 老龄林群落乔木层植物18科26属54种, 灌木层植物22科35属67种。同一林龄乔木层和灌木层优势种相似性较高。
| 种名Species | 乔木层Tree layer | 灌木层Shrub layer | |||||
| YF | MF | OF | YF | MF | OF | ||
| 黧蒴锥Castanopsis fissa | 18.98 | 7.12 | 19.11 | 7.02 | |||
| 杉木Cunninghamia lanceolata | 9.86 | 9.98 | |||||
| 罗浮柿Diospyros morrisiana | 9.30 | 9.43 | |||||
| 多花杜鹃Rhododendron cavaleriei | 5.03 | 3.99 | 5.12 | ||||
| 米槠Castanopsis carlesii | 4.39 | 4.84 | 4.47 | 4.66 | 3.79 | ||
| 鹿角锥Castanopsis lamontii | 3.84 | ||||||
| 华润楠Machilus chinensis | 3.85 | ||||||
| 广东润楠Machilus kwangtungensis | 3.38 | 3.45 | |||||
| 锥Castanopsis chinensis | 23.18 | 22.54 | |||||
| 红豆树Ormosia hosiei | 9.09 | 9.32 | |||||
| 阴香Cinnamomum burmanni | 7.76 | 7.50 | |||||
| 狗骨柴Diplospora dubia | 6.86 | 6.63 | |||||
| 木荷Schima superba | 6.46 | 6.03 | 6.22 | 5.88 | |||
| 山橿Lindera reflexa | 6.26 | 6.75 | 6.05 | 6.56 | |||
| 黄杞Engelhardtia roxburghiana | 5.82 | ||||||
| 鸭公树Neolitsea chuii | 4.77 | 4.60 | |||||
| 罗浮锥Castanopsis fabri | 15.99 | 15.88 | |||||
| 鼠刺Itea chinensis | 4.10 | ||||||
| 省略重要值小于10%的物种 | |||||||
研究结果如表 3所示, 单因素方差分析表明, 沿林龄梯度, 林冠开度和叶面积指数在不同林龄阶段均有显著差异(P < 0.05)。中龄林林冠开度最低, 叶面积指数最高。幼龄林和老龄林冠层结构指标之间均没有显著的差别。
| 林分Forest type | 林冠开度Canopy openness/% | 叶面积指数Leaf area index |
| YF | 6.3276±0.164a | 3.4742±0.070b |
| MF | 5.7496±0.153b | 3.7091±0.063a |
| OF | 6.2265±0.276a | 3.3968±0.048b |
| P | 0.021 | 0.001 |
| 不同小写字母代表差异显著P < 0.05;每个值代表平均值±标准误 | ||
由图 1可以看出, 3个不同林龄群落间的透光率、R/FR、Bw/Rw的差异性均极显著(P < 0.001)。其中透光率表现为随着林龄的增加而减少, R/FR为中龄林中最大, 幼龄林次之, 老龄林中最小;Bw/Rw为中龄林中最小, 幼龄林次之, 老龄林中最大。

|
| 图 1 林龄梯度下的透光率和光质(R/FR Bw/Rw) Fig. 1 Light transmittance and R/FR Bw/Rw in age gradient forests R/FR, 红光/远红光red/far red; Bw/Rw, 宽带红光/宽带蓝光broadband blue/broadband red |
透光率和光质比之间的非线性拟合分析表明, R/FR与透光率呈极显著非线性正相关(图 2), Bw/Rw与透光率呈极显著非线性负相关(图 3)。其中在阴天, R/FR与透光率在幼龄林(R2=0.155, P < 0.001)、中龄林(R2=0.106, P < 0.001)和老龄林(R2=0.169, P < 0.001)呈极显著正相关;Bw/Rw与透光率在幼龄林(R2=0.23, P < 0.001)、中龄林(R2=0.117, P < 0.001)和老龄林(R2=0.276, P < 0.001)呈极显著负相关。在晴天, R/FR与透光率在幼龄林(R2=0.002, P < 0.001)、中龄林(R2=0.051, P < 0.001)和老龄林(R2=0.194, P < 0.001)中呈极显著正相关;Bw/Rw与透光率在幼龄林(R2=0.111, P < 0.001)、中龄林(R2=0.197, P < 0.001)和老龄林(R2=0.395, P < 0.001)中呈极显著负相关。其拟合关系R2值均较低, 说明透光率对R/FR和Bw/Rw的解释力度均较低。

|
| 图 2 林龄梯度下透光率与R/FR之间的关系 Fig. 2 The relationship between light transmittance and R/FR in age gradient forests R/FR, 红光/远红光red/far red; |

|
| 图 3 林龄梯度下透光率与Bw/Rw之间的关系 Fig. 3 The relationship between transmittance and Bw/Rw in age gradient forests Bw/Rw, 宽带红光/宽带蓝光broadband blue/broadband red |
不同光环境林下幼树性状的方差分析结果表明, 重叠率在不同光环境下差异显著(P < 0.05), LNC在不同光环境下差异极显著(P < 0.001), 空隙率、SLA和高径比在不同光环境下无显著差异(图 4), 进一步对功能性状进行多重比较, 结果表明:幼龄林重叠率显著高于老龄林, 中龄林重叠率和幼龄林、老龄林相比没有显著差别;幼龄林下幼树LNC最小(12.472±0.277), 显著低于中林龄和老龄林。

|
| 图 4 同光环境下林下幼树功能性状差异性 Fig. 4 Differences of understory saplings functional traits under different light environments YF, 幼龄林young forest; MF, 中龄林middle-aged forest; OF, 老龄林old-growth forest |
如图 5所示, 3个树种在不同光环境下净光合速率(Pn)随光合有效辐射(PAR)的变化趋势基本一致, 均表现为:低PAR时Pn迅速增加, 达到一定的PAR后, Pn增幅逐渐趋于平缓。采用叶子飘模型[37]对3个树种的光响应曲线进行了模拟及参数计算, 结果显示, 各方程的决定系数均在0.95以上。说明该模型适合于不同光环境条件下的各树种, 较好地反映了叶片光合对光的响应过程。

|
| 图 5 不同光环境下少花桂、锥和木荷光响应曲线 Fig. 5 The response curves of net photosynthetic rate to light intensity of Cinnamomum pauciflorum、Castanopsis chinensis and Schima superba in different light environments |
3个树种在不同光环境下Pn-PAR曲线的高低呈不同的变化规律。少花桂Pn-PAR曲线的高低在不同光环境下显示出较大的差异, 在同等PAR时的Pn值表现为幼龄林>中龄林>老龄林, 其中, Pn值在中龄林和老龄林中差异不明显;锥Pn-PAR曲线的高低在不同光环境下显示出较大的差异, 在同等PAR时的Pn值表现为老龄林>中龄林>幼龄林;木荷Pn-PAR曲线在不同光环境下无明显差异。3个树种的暗呼吸速率(Rd)和光饱和点(Im)在不同光环境下无显著差异, 少花桂和锥的最大净光合速率(Pnmax)在不同光环境下有显著差异(P < 0.05)。少花桂在幼龄林林下的Pnmax值最大, 为5.958±1.001, 而锥在老龄林林下Pnmax最大为:5.924±0.796, 木荷的Pnmax在不同光环境下无显著差异。
4 讨论 4.1 冠层结构与林下光环境的关系通过对天然常绿阔叶林林龄梯度下冠层结构和林下光环境特征进行研究可知, 随着林龄的增加, 林冠开度表现为先减少后增加, 叶面积指数表现为先增加后减少。透光率随着林龄增长逐渐降低, R/FR呈现出总体降低的趋势, Bw/Rw则呈现增加趋势。因此在林冠上方光环境相似的情况下, 天然常绿阔叶林冠层结构对林下光环境的影响较大, 在一定程度上决定着林下光环境特征。有研究发现:随着林龄的增加, 林冠开度基本呈减小的趋势, 叶面积指数呈增加的趋势[38-39]。但本研究结果表明, 中龄林林冠开度最小, 林冠较为郁闭, 其林冠开度值为5.75%, 叶面积指数最大为3.71, 对光的截获能力大, 因此其林下光照较弱, 透光率较低, 而到了老龄林阶段林冠开度增加, 叶面积指数减少。这可能与我们选择的老龄林到了过熟龄阶段有关, 黄柳菁等[40]对广东不同林龄乔木层叶面积指数的研究中也指出, 中龄林的乔木层叶面积指数显著高于幼龄林和过熟林。
相关研究表明, 森林冠层叶面积指数的增大能引起林分透光率的下降[41-42], 本研究结果显示透光率与叶面积指数的变化规律并不一致, 老龄林冠层叶面积指数低于中龄林, 但是透光率低于中龄林, 其值最低为0.328, 这可能是因为老龄林附近有地形遮挡导致白天光照强度较低, 当光照强度较低时, 林冠透光率会明显下降[43]。林下光质的变化在不同的林龄间差异显著(图 1)。
通常来说, 与冠层上方的光合有效辐射相比, 入射到冠层下方太阳辐射光谱组成中的蓝色部分占比相对较低, 同时红色(650—700nm)部分轻微增加, 远红光(700—750nm)明显增加[44]。并且林下光照强时富含红光, 光照弱时会富含更多的远红光[45]。我们观测到老龄林中R/FR比值明显低于幼龄林, 是因为老龄林林冠比幼龄林更加郁闭, 冠层对红光的吸收相对于幼龄林较高[20, 32]且透射更多远红光。老龄林中Bw/Rw比值明显高于幼龄林, 是由于老龄林林下光照强度较低, 当透光率减少时, 冠层内红光相较蓝光下降更快[46]。中龄林光质的异常情况可能是中龄林样地株密度(0.18株/m2)显著低于幼龄林和老龄林所致, 高的株密度能够降低林下R/FR[47], 因此, 老龄林和幼龄林中的R/FR比中龄林低。综上所述, 天然林冠层结构在很大程度上对林下光强和光质有影响, 叶面积指数高、林冠开度低的林分林下光照强度越弱, R/FR越低, Bw/Rw越高。
在这项研究中, 透光率与R/FR之间极显著正相关(P < 0.001), 这与Leuchner和Menzel等[20]对德国南部温带云杉-山毛榉混交林透光率和R/FR的研究一致。随着透光率的增加红光占比增多, 导致了较高的R/FR值。透光率和R/FR之间的关系非常复杂, 并非简单的线性相关。Leuchner选择了矩形双曲线函数来描述温带混交林透光率和R/FR之间的关系, 且透光率对R/FR的解释度在80%左右;Caper和Chazdon[32]提出了一种通过测量热带森林中的R/FR迅速评估透光率的方法, 所获得的透光率与R/FR之间的关系是对数的, 且阴天情况下相关系数高达97%。本研究结果中透光率与R/FR之间的关系也符合对数函数关系, 但是无论是晴天还是阴天相关系数都较弱, 其原因可能是天然林林分组成复杂导致林下光照变化复杂。本研究结果表明透光率与Bw/Rw之间极显著负相关(P < 0.001)。国外学者对挪威云杉冠层林下光环境研究显示晴天条件下随着太阳光照强度减弱, 林下蓝光占比相较于红光增加明显[46], 从而引起了Bw/Rw比值的增加。本文的研究结果显示无论晴天还是阴天, 亚热带天然常绿阔叶林透光率的减弱都会引起Bw/Rw的增加。
综合来看, 冠层叶面积指数随着林龄的增加, 在一定阶段范围内也会随之增加, 林冠开度与之相反。林下光环境与冠层结构也紧密相连。冠层郁闭程度加深, 透光率就会逐渐减弱;同时林下红光和蓝光占比减少, 远红光占比增多, 导致R/FR减少, 蓝光占比较红光略高, 导致Bw/Rw增加, 天然林透光率与光质之间存在显著相关关系, 但是相关程度一般。
4.2 林下幼树对不同光环境的响应林下幼树对林龄梯度下光强和光质的响应存在差异, 主要通过叶片功能性状特征及光合生理特性的不同来实现。
老龄林林下幼树叶片重叠率显著低于幼龄林, 中龄林和老龄林林下幼树LNC显著高于幼龄林。幼龄林林下光照资源充足, 林下幼树通过增加叶片重叠率来充分利用光资源, 积累更多的光合产物, 老龄林林下幼树由于光照强度降低, 光资源不足, 其叶片重叠率也因此降低。氮是光合结构蛋白质的重要组分, LNC很大程度上决定了植物的光合能力[48]。老龄林林下幼树通过增加单位干质量的氮含量来提高净光合速率[49], 使其具有较强的养分保持能力和耐荫能力。而在幼龄林高光强的环境下, 林下幼树可能倾向于将较多的氮投资于保护构造上以防止高温损伤和过度失水[50], 充足的光照使林下幼树不必将过多的氮投资给光合器官。研究表明[45], 在演替阶段早期, 森林冠层郁闭度较小, 光照充足, 幼龄林林下幼树具有阳生植物的特性, 生长需要光照条件较好的生境。相应地, 演替阶段后期植物具有阴生植物的特性, 能够耐受荫蔽的生境。植物的这种特征可能对其光合作用产生一定的影响。我们通过对不同林龄梯度下共有树种(少花桂、锥和木荷)光响应曲线进行研究发现, 它们在不同光环境下的最大净光合速率有显著差异, 主要受到光照强度变化的影响。演替前期种少花桂在光照充足的环境中具有较高的光合能力, 较耐荫的后期演替种锥在光照强度较弱的环境中具有较高的光合能力, 演替中期种木荷在不同的光照条件下光合能力相近。
植物对光环境的响应不仅仅体现在叶片功能性状方面, 在整体形态方面也有响应:林下弱光环境下, 红光和蓝光的减少, 远红光的增加都在诱导植物的整体形态发生反应[51]。R/FR的减少会促使植物伸长生长, 而本研究中植物的高径比在林龄梯度下差异不显著, 可能是因为天然林生境复杂, 植物高径比会受到其他因素(如凋落物和种子特性等[52])的影响。
总之, 在天然常绿阔叶林不林龄梯度光环境下, 林下幼树叶片重叠率和LNC随着光照强度的减弱, 重叠率减少, LNC增加。林下幼树功能性状的变化主要受到光照强度的影响。此外, 林下共有种在光合响应方面表现出了差异, 这说明植物的光合生理特征同时受光环境和植物物种自身特性的影响。
5 结论通过对南岭区域天然常绿阔叶林林龄梯度下冠层结构、林下光环境和林下幼树功能性状的研究, 我们发现随着林龄的增加, 整体趋势呈现出叶面积指数随之增加, 林冠开度随之减少;林龄梯度下光照强度和光质特征有一定的差异, 光照强度的减少使得林下光质占比发生了变化, 红光和蓝光减少, 远红光增多, R/FR值降低, Bw/Rw增加。透光率和光质比之间有显著相关关系, 但是透光率对光质的解释程度很低, 可能是因为我们观测的是天然林而非人工林或者成分简单的混交林。光照强度的减弱使得林下幼树重叠率减少, LNC增多。R/FR和Bw/Rw的变化对林下幼树的高径比并无显著影响。
综上所述, 不同的林龄阶段, 冠层结构和林下光环境具有差异性, 这在一定程度上影响着林下幼树的生长和更新。通过对林分冠层结构和林下光环境的研究, 有助于我们解释林下幼树功能性状指标的差异性, 从而为探讨森林幼苗更新机制、推测幼树的适应策略及未来的发展方向提供理论依据。对了解森林的结构及生物多样性的维持也有着重要的作用, 同时为森林保护、森林生态恢复和合理经营以及保持森林生态系统的可持续发展提供可靠的理论依据。
| [1] |
李德志, 臧润国. 森林冠层结构与功能及其时空变化研究进展. 世界林业研究, 2004, 17(3): 12-16. |
| [2] |
胡启鹏, 郭志华, 李春燕, 马履一. 不同光环境下亚热带常绿阔叶树种和落叶阔叶树种幼苗的叶形态和光合生理特征. 生态学报, 2008, 28(7): 3262-3270. |
| [3] |
Chazdon R L. Sunflecks and their importance to forest understorey plants. Advances in Ecological Research, 1988, 18: 1-63. |
| [4] |
de Freitas C R, Enright N J. Microclimatic differences between and within canopy gaps in a temperate rainforest. International Journal of Biometeorology, 1995, 38(4): 188-193. |
| [5] |
Press M C, Brow N D, Barker M G, Zipperlen S W. Photosynthetic responses to light in tropical rain forest tree seedlings//Swaine M D, ed. The Ecology of Tropical Forest Tree Seedlings. Paris: UNESCO Paris and The Parthenon Publishing Group, 1996: 41-58.
|
| [6] |
Downs R J. Light and Plants:A Series of Experiments Demonstrating Light Effects on Seed Germination, Plant Growth, and Plant Development. Washington: Agricultural Research Service, 1961.
|
| [7] |
Lieffers V J, Messier C, Stadt K J, Gendron F, Comeau P G. Predicting and managing light in the understory of boreal forests. Canadian Journal of Forest Research, 1999, 29(6): 796-811. |
| [8] |
Montgomery R A, Chazdon R L. Forest structure, canopy architecture, and light transmittance in tropical wet forests. Ecology, 2001, 82(10): 2707-2718. |
| [9] |
Mejía-Domínguez N R, Meave J A, Díaz-Ávalos C. Spatial structure of the abiotic environment and its association with sapling community structure and dynamics in a cloud forest. International Journal of Biometeorology, 2012, 56(2): 305-318. |
| [10] |
区余端, 苏志尧. 粤北山地常绿阔叶林自然干扰后冠层结构与林下光照动态. 生态学报, 2012, 32(18): 5637-5645. |
| [11] |
杨建宇.古田山常绿阔叶林冠层结构及其光环境特征[D].齐齐哈尔: 齐齐哈尔大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10232-1015622905.htm
|
| [12] |
张彦雷, 康峰峰, 韩海荣, 尹学哲, 程小琴, 李勇, 周彬, 刘可, 孙敬军. 太岳山油松人工林冠下光环境特征与冠层结构. 南京林业大学学报:自然科学版, 2014, 38(2): 169-174. |
| [13] |
Zhu J J, Matsuzaki T, Lee F Q, Gonda Y. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. Forest Ecology and Management, 2003, 182(1/3): 339-354. |
| [14] |
崔佳玉, 曾焕忱, 王永强, 张毅, 胡益珩, 苏志尧. 银瓶山自然保护区阔叶林冠层结构与辐射消减效应. 西北林学院学报, 2015, 30(4): 45-49. |
| [15] |
黄练忠, 杨进良, 徐庆华, 张星元, 阚蕾, 张璐. 城市森林群落林冠结构与林下光环境的关系. 中南林业科技大学学报, 2019, 39(9): 53-58. |
| [16] |
Wassink E C, Stolwijk J A J. Effects of light quality on plant growth. Annual Review of Plant Physiology, 1956, 7(1): 373-400. |
| [17] |
Dengel S, Grace J, MacArthur A. Transmissivity of solar radiation within a Picea sitchensis stand under various sky conditions. Biogeosciences, 2015, 12(14): 4195-4207. |
| [18] |
Perot T, Mårell A, Korboulewsky N, Seigner V, Balandier P. Modeling and predicting solar radiation transmittance in mixed forests at a within-stand scale from tree species basal area. Forest Ecology and Management, 2017, 390: 127-136. |
| [19] |
Bellow J G. The overstory book: cultivating connections with trees. Elevitch, C., Wilkinson, K. (Editors). Agroforestry Systems, 2002, 54(3): 245-247.
|
| [20] |
Leuchner M, Menzel A, Werner H. Quantifying the relationship between light quality and light availability at different phenological stages within a mature mixed forest. Agricultural and Forest Meteorology, 2007, 142(1): 35-44. |
| [21] |
Hertel C, Leuchner M, Menzel A. Vertical variability of spectral ratios in a mature mixed forest stand. Agricultural and Forest Meteorology, 2011, 151(8): 1096-1105. |
| [22] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学:生命科学, 2015, 45(4): 325-339. |
| [23] |
胡梦瑶, 张林, 罗天祥, 沈维. 西藏紫花针茅叶功能性状沿降水梯度的变化. 植物生态学报, 2012, 36(2): 136-143. |
| [24] |
王玉平, 陶建平, 刘晋仙, 何泽. 不同光环境下6种常绿阔叶林树种苗期的叶片功能性状. 林业科学, 2012, 48(11): 23-29. |
| [25] |
蒲高斌, 刘世琦, 刘磊, 任丽华. 不同光质对番茄幼苗生长和生理特性的影响. 园艺学报, 2005, 32(3): 420-425. |
| [26] |
崔瑾, 马志虎, 徐志刚, 张欢, 常涛涛, 刘海俊. 不同光质补光对黄瓜、辣椒和番茄幼苗生长及生理特性的影响. 园艺学报, 2009, 36(5): 663-670. |
| [27] |
Johkan M, Shoji K, Goto F, Hashida S N, Yoshihara T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience, 2010, 45(12): 1809-1814. |
| [28] |
Yorio N C, Goins G D, Kagie H R, Wheeler R M, Sager J C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. HortScience, 2001, 36(2): 380-383. |
| [29] |
Baraldi R, Rossi F, Facini O, Fasolo F, Rotondi A, Magli M, Nerozzi F. Light environment, growth and morphogenesis in a peach tree canopy. Physiologia Plantarum, 1994, 91(2): 339-345. |
| [30] |
Beaudet M, Messier C. Variation in canopy openness and light transmission following selection cutting in northern hardwood stands:an assessment based on hemispherical photographs. Agricultural and Forest Meteorology, 2002, 110(3): 217-228. |
| [31] |
Fielder P, Comeau P. Construction and Testing of an Inexpensive PAR Sensor. British Columbia: Ministry of Forests Research Program, 2000.
|
| [32] |
Capers R S, Chazdon R L. Rapid assessment of understory light availability in a wet tropical forest. Agricultural and Forest Meteorology, 2004, 123(3/4): 177-185. |
| [33] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000.
|
| [34] |
吴能表, 谈锋, 肖文娟, 王小佳. 光强因子对少花桂幼苗形态和生理指标及精油含量的影响. 生态学报, 2005, 25(5): 1159-1164. |
| [35] |
邵宜晶, 俞梦笑, 江军, 曹楠楠, 褚国伟, 闫俊华. 鼎湖山3种演替阶段森林土壤C、N、P现状及动态. 热带亚热带植物学报, 2017, 25(6): 523-530. |
| [36] |
李昊民, 李宁云, 喻庆国, 杨宇明. 天童山常绿阔叶林优势树种光合生理特性在其群落演替中的生态意义研究. 西部林业科学, 2011, 40(2): 13-20. |
| [37] |
Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa. Photosynthetica, 2007, 45(4): 637-640. |
| [38] |
丁圣彦, 卢训令, 李昊民. 天童国家森林公园常绿阔叶林不同演替阶段群落光环境特征比较. 生态学报, 2005, 25(11): 2862-2867. |
| [39] |
黄慧敏, 董蓉, 何丹妮, 向运蓉, 张小晶, 陈娟, 陶建平. 冠层结构和光环境的时空变化对紫耳箭竹种群特征的影响. 应用生态学报, 2018, 29(7): 2129-2138. |
| [40] |
黄柳菁, 林欣, 刘兴诏, 庄长伟, 肖荣波. 广东不同林龄乔木生物量及物种多样性与叶面积指数的关系. 西南林业大学学报, 2017, 37(6): 91-98. |
| [41] |
班宏娜.樟子松人工林树冠层光分布规律及对生长影响的研究[D].哈尔滨: 东北林业大学, 2010. http://d.wanfangdata.com.cn/Thesis/Y2051107
|
| [42] |
贾小容, 苏志尧, 区余端, 解丹丹. 三种人工林分的冠层结构参数与林下光照条件. 广西植物, 2011, 31(4): 473-478. |
| [43] |
Leuchner M, Hertel C, Menzel A. Spatial variability of photosynthetically active radiation in European beech and Norway spruce. Agricultural and Forest Meteorology, 2011, 151(9): 1226-1232. |
| [44] |
Combes D, Sinoquet H, Varlet-Grancher C. Preliminary measurement and simulation of the spatial distribution of the Morphogenetically Active Radiation (MAR) within an isolated tree canopy. Annals of Forest Science, 2000, 57(5): 497-511. |
| [45] |
李庆康, 马克平. 植物群落演替过程中植物生理生态学特性及其主要环境因子的变化. 植物生态学报, 2002, 26(S1): 9-19. |
| [46] |
Navrátil M, Špunda V, Marková I, Janouš D. Spectral composition of photosynthetically active radiation penetrating into a Norway spruce canopy:the opposite dynamics of the blue/red spectral ratio during clear and overcast days. Trees, 2007, 21(3): 311-320. |
| [47] |
Casal J J, Sanchez R A, Deregibus V A. The effect of plant density on tillering:the involvement of R/FR ratio and the proportion of radiation intercepted per plant. Environmental and Experimental Botany, 1986, 26(4): 365-371. |
| [48] |
张继光, 傅声雷, 温达志, 张玲玲. 南亚热带人工林16种木本植物重要叶特征参数的相互关系. 热带亚热带植物学报, 2009, 17(4): 395-400. |
| [49] |
赵平, 孙谷畴, 彭少麟. 植物氮素营养的生理生态学研究. 生态科学, 1998, 17(2): 37-42. |
| [50] |
张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844-852. |
| [51] |
Ammer C. Growth and biomass partitioning of Fagus sylvatica L. and Quercus robur L. seedlings in response to shading and small changes in the R/FR-ratio of radiation. Annals of Forest Science, 2003, 30(2): 163-171. |
| [52] |
郑维娜, 王孝安, 郭华, 王丹, 刘鹏, 刘史力. 微生境对辽东栎幼苗生长的影响. 干旱区研究, 2013, 30(6): 1049-1055. |