 2020, Vol. 40
2020, Vol. 40文章信息
- 李坤, 邢小艺, 李逸伦, 李晓鹏, 李如辰, 范舒欣, 董丽
- LI Kun, XING Xiaoyi, LI Yilun, LI Xiaopeng, LI Ruchen, FAN Shuxin, DONG Li
- 石林风景区不同石漠化人工修复方式对木本植物群落组成及种群生态位的影响
- Effect of different artificial restoration methods of Karst rocky desertification on community composition and niche characteristics of woody populations in Shilin scenic area
- 生态学报. 2020, 40(13): 4641-4650
- Acta Ecologica Sinica. 2020, 40(13): 4641-4650
- http://dx.doi.org/10.5846/stxb201911052324
-
文章历史
- 收稿日期: 2019-11-05
- 修订日期: 2020-03-29





2. 城乡生态环境北京实验室, 北京 100083;
3. 国家花卉工程技术研究中心, 北京 100083
2. Beijing Laboratory of Urban and Rural Ecological Environment, Beijing 100083, China;
3. National Engineering Research Center for Floriculture, Beijing 100083, China
中国西南地区是世界三大喀斯特集中分布区之一, 受地球强烈的地质运动和碳酸盐岩溶蚀性强等因素的综合影响, 喀斯特生态系统稳定性和抗干扰能力差, 与黄土、沙漠和寒漠并列为中国四大生态环境脆弱区[1-2]。自19世纪以来, 我国西南喀斯特地区由于大范围受到人类不合理经济活动的干扰, 导致土壤侵蚀、森林退化与生物多样性急剧下降, 最终演变为石漠化[3]。20世纪70年代以来, 随着我国政府对石漠化问题的重视, 在大量研究的基础上采取了多种生态治理措施。在轻度石漠化地区, 通过封山育林保护植被, 依靠自然恢复促进正向演替;在中度石漠化地区, 通过人工结合自然恢复的方式促进正向演替;对于重度石漠化地区, 自然恢复艰难且漫长, 主要通过综合的方式进行治理[4], 包括生态移民、人工飞播和人工栽植等。石漠化植被恢复的科学研究主要集中在生态学和林学等领域。其中, 生态学的研究主要包括自然演替进程及演替动力[5]等;在林学方面, 许多学者对先锋树种, 造林技术与模式[6]进行研究, 从人工林的多样性[7]、土壤理化性质及微生物特征[8]、水土保持能力[9]等方面进行生态功能评价, 也有研究对人工林的经济成本及效益[10]进行探讨。大量研究表明, 在人为干扰严重的石漠化地区, 与自然恢复相比, 人工修复能较快构建植被, 从而相对较快地发挥生态和经济效益。但是不容忽视的是, 迄今石漠化地区人工修复的实践中仍然存在很多问题, 例如大规模爆破岩石和大范围人工覆土, 导致极高的人工修复成本;大量应用桉树等外来植物或单一树种造林, 导致群落结构失衡[11], 对当地生物多样性造成严重威胁, 形成大规模的低效人工林[12]。
国内外大量关于生态修复的研究表明, 采用近自然的理念模拟地带性植被特征是最可持续的修复方式[13]。地带性植被是自然选择的结果, 具有独特的种群组成, 稳定的层次结构和季相特征, 蕴含着丰富的乡土树种, 是石漠化生态修复的重要参考系。但是, 上述石漠化修复的问题都表明迄今缺乏对喀斯特地区地带性自然植被的深入研究。
我国云南昆明石林是西南地区重要的喀斯特研究区, 在19世纪末出现过严重的石漠化现象。1931年成立石林地质公园以来, 当地针对其不同的石漠化程度进行了自然恢复与人工修复。其中, 自然恢复主要为封山育林, 人工修复主要包括栽植园林植被与飞播云南松种子[14]。经过多年治理, 石漠化土地逐渐恢复, 石林风景区发展为喀斯特地貌、喀斯特森林与独特民族文化相融合的良好生态系统, 并于2002年入选联合国世界自然遗产, 也成为喀斯特石漠化修复的典范[15]。那么, 石林风景区石漠化区域采取的两种人工修复方式建成的植被历经几十年的演替后, 其特征是什么?有何差异?是否符合风景区地带性植被营建的目标?是否能够为当前以及未来石漠化修复提供理论依据及实践指导?这些问题并没有得到业界的重视。
生态位是种群生态学理论研究的核心。生态位特征定量地反映了物种之间、物种与环境之间的相互关系, 能够揭示群落中物种组成及其在生态系统中的功能地位[16]。生态位理论在解释群落构建与生物多样性维持机制[17]、预测群落演替方向[18]和评估植被恢复效果中具有重要意义。
本研究基于生态位理论, 以石林风景区两类人工修复植被为研究对象, 以地带性自然植被为参考, 以期回答以下科学问题:(1)栽植植被(PV)、飞播植被(ASV)、地带性自然植被(ZNV)3种类型植被的木本植物群落组成及数量特征是否存在差异;(2)3种植被类型中优势木本植物种群的生态位是否相同以及生态位特征的差异大小, 从而科学评价修复后植物种群对资源的利用程度和对环境的适应能力, 为喀斯特风景区或相似喀斯特石漠化地区植物景观生态恢复树种规划及群落构建提供理论指导与科学依据。
1 研究地区与研究方法 1.1 研究区概况云南石林风景区位于云南省昆明市石林彝族自治县境内(24°30′—25°03′N, 103°10′—104°40′ E), 占地面积350 km2, 核心景区面积为120 km2, 主体海拔1780—1950 m, 有明显的雨季和旱季之分, 每年5—10月为雨季, 11月至次年4月为旱季, 年降雨量约960 mm, 年平均温度15.6℃, 属亚热带高原干湿季风气候区[14]。土壤类型主要为发育面积广大的山原红壤和石灰土等。地带性自然植被(ZNV)主要为半湿润常绿阔叶林。从17世纪末至19世纪末, 随着铜矿采炼、薪炭林砍伐和过度放牧[14], 石林风景区地带性植被逐渐退化, 生物多样性急剧下降, 岩石大量出露, 最终变成“地瘠民贫”的石漠化荒地, 仅在当地民族村寨保护的密枝山保留了少量原生性强的半湿润常绿阔叶林[19]。
本文的研究样地包括石林风景区人工栽植及人工飞播两种修复方式下的代表性植被样地, 分别为:(1)1950—1982年在游览区外围人工飞播云南松(Pinus yunnanensis)[14], 修复面积约为10 hm2, 人工维护管理强度较低, 林下自然演替, 目前石林地区的云南松林龄为35—45年[11, 20], 本研究中将这种修复方式下的植被类型定义为飞播植被(ASV);(2)1982年核心游览区内采用人工植物群落构建的修复方式[14], 修复面积约为19 hm2, 应用的植物种类主要包括黄连木(Pistacia chinensis)和清香木(Pistacia weinmannifolia)等, 主要群落结构为乔灌结构与乔灌草结构, 植被生长年限为35年左右, 人工维护管理强度较低, 研究中定义该植被类型为栽植植被(PV)。
本文的对照样地为地带性自然植被(ZNV), 为石林风景区核心景区之外保存较为完整的半湿润常绿阔叶林——月湖彝族村落的“密枝山”森林。
1.2 样地及样方设置本研究采用典型样地取样法, 各建立10个20 m×20 m的乔木样方, 共计30个样方。每个乔木样方划分成4个10 m×10 m的灌木样方, 记录乔木样方内所有胸径(DBH)≥ 3 cm乔木的物种名称、株数、高度、胸径、冠幅、相对坐标等, 记录灌木样方内所有灌木(包括胸径<3 cm的乔木幼树), 植物信息记录指标同上, 同时记录经纬度、海拔、坡度、坡向和坡位等指标。本研究所选择的调查样地类型如下(表 1):
| 类型 Type |
样地 Site |
经纬度 Longitude and latitude |
海拔 Altitude/m |
坡度 Slope/° |
坡向 Aspect |
基岩类型 Rock type |
岩溶特征 Karst type |
土石比例 Ratio |
恢复时间 Time |
| 地带性自然植被ZNV | 月湖村“密枝林” | 103°25′48″E24°54′01″N | 1900 | 10—15 | — | 灰岩 | 石芽, 少量石柱 | 2:3 | — |
| 栽植植被PV | 特级保护区, 大小石林及李子园箐附近 | 103°20′52″E24°48′17″N | 1750—1790 | 10—20 | — | 灰岩 | 石柱, 石芽发育强烈 | 2:3 | 35年左右 |
| 飞播植被ASV | 一级保护区, 风景区中南部绿方塘附近 | 103°19′34″E24°49′50″N | 1800—1840 | 15—30 | 东南坡 | 灰岩 | 石芽, 少量石柱 | 3:1 | 35年 |
| ZNV:地带性自然植被, Zonal natural vegetation;PV:栽植植被, Planted vegetation;ASV:飞播植被, Air seeding vegetation | |||||||||
乔木层重要值:
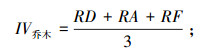
|
灌木层重要值:
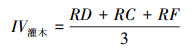
|
式中, RD为相对密度, RA为相对优势度, RC为相对盖度, RF为相对频度。
1.3.2 丰富度Patrick丰富度指数:S=所在样方的物种数
1.3.3 α多样性Shannon-Wiener多样性指数:H=-∑PilnPi;Pielou均匀度指数:J=H/lnS
式中, Pi为物种i的重要值。
1.3.4 β多样性Jaccard指数:C=j/(a+b-j)
式中, j为两个样地中共有的物种数, a为样地A中的物种数, b为样地B中的物种数。
1.3.5 生态位Levins生态位宽度指数: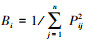
式中, pij为物种i在第j个资源状态下的重要值。
Pianka重叠指数: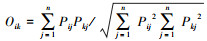
式中, Pij和Pkj分别为物种i与物种k在样方j的重要值。
样方整理和数据计算采用Excel 2016和R 3.5.1, 多样性指数的计算采用R 3.5.1的Vegan包进行, 多样性指数的均值比较运用agricolae包, 采用Duncan检验法(α=0.05)进行;生态位宽度和生态位重叠采用spaa包进行;统计图由Excel和R 3.5.1绘制。植物物种信息的整理参考《云南植物志》和《中国植物志》。其中, 乡土植物为自然分布于云南地区的物种, 外来植物为人类有意或无意引进的云南范围以外的物种, 分为国外外来与国内外来植物, 通过查询中国植物志(http://frps.iplant.cn/)确定。
2 研究结果 2.1 木本植物物种构成特征本研究中, 在地带性自然植被、栽植植被和飞播植被3种植被类型中分别记录到木本植物92种(45科80属)、138种(50科101属)与44种(26科37属)。从生活型来看, 地带性植被中木本植物种数所占比例呈现为灌木(48.91%)>乔木(42.39%)>木质藤本(7.61%)>竹类(1.09%)(图 1), 而栽植植被与飞播植被两种人工修复植被中木本植物种数所占比例呈现为乔木(45.65%, 54.55%)>灌木(44.93%, 43.18%)>木质藤本(7.97%, 2.27%)>竹类(1.45%, 0)。3种植被类型中, 落叶植物种数所占比例相近, 约为55%, 高于常绿植物所占比例(约为45%), 符合半湿润常绿阔叶林的特征。3种植被类型中所含植物物种数最多的科都是蔷薇科, 其次是豆科。此外, 栽植植被中木犀科的物种种类较多(6种), 但是在地带性自然植被和飞播植被中都仅为云南木犀榄(Olea yuennanensis)一种。

|
| 图 1 3种植被类型中木本植物物种构成特征 Fig. 1 Species characteristics of woody plants in three vegetation types |
从物种来源上看, 均以乡土树种为主, 例如清香木等, 外来植物所占比例很低(<5.10%), 主要有蓝桉(Eucalyptus globulus)、马缨丹(Lantana camara)等。
2.2 木本植物群落多样性特征 2.2.1 α多样性特征整体上, 乔木层与灌木层中, 3种植被类型的Patrick丰富度指数与Shannon-Wiener多样性指数排序均呈现为地带性自然植被>栽植植被>飞播植被(P<0.01);栽植植被与飞播植被之间的Pielou均匀度指数差异不显著, 但是均显著高于地带性自然植被(P<0.001)(图 2)。

|
| 图 2 3种不同植被类型下木本植物群落多样性比较 Fig. 2 Comparison of woody plants community diversity in three vegetation types 不同小写字母表示不同修复方式下差异显著(P<0.05) |
乔木层中, 栽植植被的Patrick丰富度指数波动范围为2.00—9.00, Shannon-Wiener多样性指数波动范围为1.43—7.84, 接近地带自然植被的Patrick丰富度指数(3.00—11.00)与Shannon-Wiener多样性指数(1.66—8.13)的分布范围。而飞播植被Patrick丰富度指数与Shannon-Wiener多样性指数的波动范围最小, 分别为1.00—5.00和1.00—4.24, 物种的多样性显著低于栽植植被和地带性自然植被(P<0.001)。
灌木层中, 栽植植被的Patrick丰富度指数与Shannon-Wiener多样性指数变化范围最大, 为2.00—11.00和1.67—9.51, 飞播植被的Patrick丰富度指数与Shannon-Wiener多样性指数变化范围最小, 为1.00—7.00和1.00—6.00。飞播云南松的植被类型中灌木层物种丰富度与多样性显著低于栽植植被和地带性自然植被(P<0.001)。
2.2.2 β多样性特征从3种植被类型的β多样性指数来看, 栽植植被与地带性自然植被中Jaccard指数为0.14, 共有种29种, 包括清香木等(表 2);飞播植被与地带性自然植被中Jaccard指数为0.12, 共有种14种, 包括滇润楠(Machilus yunnanensis)等;栽植植被与飞播植被中Jaccard指数为0.23, 共有种34种, 包括云南松等。3种植被类型中共有种有12种, 包括清香木、毛叶合欢(Albizia mollis)和黄连木等。
| 类型 Type |
木本植物种类 Species of woody plants |
种数 Species number |
| 3种植被的共有种 Common species in three vegetation types |
毛叶合欢(Albizia mollis)、黄连木(Pistacia chinensis)、滇润楠(Machilus yunnanensis)、清香木(P.weinmanniifolia)、团花新木姜子(Neolitsea homilantha)等 | 12 |
| 栽植植被与地带性自然植被的共有种 Common species between PV and ZNV |
华山松(Pinus armandii)、滇青冈(Cyclobalanopsis glaucoides)、云南木犀榄(Olea tsoongii)、竹叶花椒(Zanthoxylum armatum)、西南栒子(Cotoneaster franchetii)等 | 29 |
| 飞播植被与地带性自然植被的共有种 Common species between ASV and ZNV |
团花新木姜子(Neolitsea homilantha)、毡毛栒子(C.pannosus)等 | 14 |
| 栽植植被与飞播植被的共有种 Common species between PV and ASV |
云南松(P. yunnanensis)、华南五针松(P. kwangtungensis)、西藏柏木(Cupressus torulosa)、川滇无患子(Sapindus delavayi)、枫香树(Liquidambar formosana)等 | 34 |
| 3种植被中的共有种未在两种植被的共有种中重复列出 | ||
乔木层中, 地带性自然植被、栽植植被与飞播植被中生态位宽度最大的植物依次为云南木犀榄、黄连木和云南松。在生态位宽度前10位的物种中, 3种植被类型中的共有种3种, 为清香木、黄连木和滇润楠, 清香木和滇润楠在栽植植被中的生态位宽度更接近地带性自然植被中的生态位宽度(图 3)。栽植植被和地带性自然植被共有种3种;飞播植被和地带性自然植被共有种4种, 包括团花新木姜子(Neolitsea homilantha)等;栽植植被和飞播植被中共有种5种, 包括干香柏(Cupressus duclouxiana)等。

|
| 图 3 3种植被类型中木本植物生态位宽度前10位 Fig. 3 Niche width of top 10 woody plant species in three vegetation types |
灌木层中, 地带性自然植被、栽植植被与飞播植被中生态位宽度最大的植物依次为铁仔(Myrsine africana)、清香木和白刺花(Sophora davidii)。在生态位宽度前10位的物种中, 3种植被类型中的共有种3种, 包括铁仔、清香木和沙针(Osyris quadripartita), 其中, 清香木生态位宽度依然很高(图 3);栽植植被和地带性植被共有种3种;飞播植被和地带性自然植被共有种4种, 包括多脉猫乳(Rhamnella martini)等;栽植植被和飞播植被中的共有种4种, 包括白刺花等。
2.3.2 生态位重叠度在3种植被类型中选择重要值排名前25位的木本植物, 组成300个种对关系进行生态位重叠分析。生态位重叠值为1, 完全重叠的种对在3种植被类型中均尚未发现;生态位重叠值等于0, 即不发生重叠的种对, 栽植植被与飞播植被中分别为154对(51.33%)与178对(59.33%), 远高于地带性自然植被中不发生重叠的种对数量(22对, 17.33%);生态位重叠值大于或等于0.5的种对, 在栽植植被中仅为2对(0.67%), 包括蓝桉×火棘(0.53), 西南栒子(Cotoneaster franchetii)×山玉兰(Magnolia delavayi)(0.57), 低于飞播植被(22对, 7.33%)与地带性自然植被(22对, 7.33%)中高生态位重叠值的种对数(图 4)。

|
| 图 4 3种不同群落类型下木本植物生态位重叠区间 Fig. 4 Niche overlap section of woody plants in three vegetation types 横坐标中0代表生态位重叠值为0;0—0.5代表生态位重叠值大于0小于0.5;0.5—1代表生态位重叠值大于等于0.5小于1 |
具有较高生态位宽度的木本植物之间具有相对较高的生态位重叠。飞播植被的木本植物中云南松与多脉猫乳、车桑子(Dodonaea viscosa)等乡土植物的生态位重叠度大于0.5, 存在竞争关系;云南松与外来植物马缨丹的生态位重叠度为0.43, 可能存在竞争关系。栽植植被的木本植物中清香木与云南松、黄连木、白刺花、沙针的生态位重叠值大于0.35, 可能存在竞争关系。相同的植物种群之间在不同的修复植被类型中生态位重叠指数差异较大。飞播植被中, 云南松与其他木本植物的生态位重叠大于栽植植被中云南松与其他木本植物的生态位重叠。
3 讨论 3.1 人工修复植被与地带性自然植被木本植物群落多样性特征对比石林风景区核心景区0.4 hm2栽植植被样地中木本植物种数共计138种(50科101属), 高于兰斯安[21]在广西木论喀斯特自然保护区0.64 hm2的地带性植被样地内统计到的木本植物种数(120种48科90属), 也高于石林地区地带性自然植被中木本植物种数(92种45科80属)。但是, 栽植植被中乔木层α多样性显著低于地带性自然植被的α多样性。栽植植被中植物种类与植物配置的数量、结构、密度是影响其多样性的关键。栽植植被在恢复初期应用的整体植物种类丰富, 但是不同景观节点上植物配置差异较大, 例如既有全部为西南栒子的群植方式, 又有以西南栒子为骨干树种, 搭配黄连木、高盆樱桃(Cerasus cerasoides)等的丛植方式, 植物种类、数量与栽植方式对群落α多样性有影响;而地带性自然植被是长期自然演替的结果, 群落中不同种群占据各自的生态位, 数量分配更均匀。同时, 由于地带性自然植被受到的人为干扰较少, 植物物种在高度与冠幅上比栽植植被更具优势, 重要值更高。因此, 地带性自然植被的α多样性更高。栽植植被与地带性自然植被的灌木层α多样性无显著差异, 但是植物种类差异较大。栽植植被中的灌木多喜阳, 例如毛枝绣线菊(Spiraea martini)和白刺花等, 而地带性自然植被中由于郁闭度较高, 优势灌木多耐阴, 例如针齿铁仔(Mysine semiserrata)等。两种植被类型之间的β多样性指数表明木本植物物种相似性较低, 也证明人工栽植选择的植物种类与地带性自然植被的植物种类仍然存在较大差异, 地带性自然植被中优势度较高的物种, 例如裂果漆(Rhus griffithii)和大叶桂樱(Laurocerasus zippeliana)等在栽植植被中均未应用。
Cindy Q等[11]与Wang等[22]研究表明飞播植被中木本植物α多样性显著低于地带性自然植被, 本研究也发现相同的结果。Cindy于2010年在石林约0.6 hm2的云南松飞播植被样地中发现木本植物24种[7], 但是在本研究中0.4 hm2相似样地中发现木本植物43种, 表明在修复初期飞播云南松种子, 提供了乔木层的先锋树种, 在一定程度上改善了土壤等环境条件, 促进了已退化的半湿润常绿阔叶林正向自然演替。目前飞播植被中除云南松和干香柏以外的乔木多以DBH<3 cm的幼树和幼苗形式存在, 未来应当对飞播植被的群落多样性进行进一步跟踪研究。
3.2 人工修复植被与地带性自然植被木本植物生态位特征对比经典生态位理论认为物种间的生态位分化, 是物种共存的机制之一, 也是群落组成变化和群落演替的主要动力[16]。生态位宽度在一定程度上表现了物种对环境的适应能力, 通常生态位宽度较高的物种在群落中的地位更高。因此, 生态位宽度可以作为筛选修复植物和预测未来植被变化[23]的重要指标之一。
本研究中清香木在3种植被类型中的乔木层和灌木层中生态位宽度排名都在前三位之内, 证明清香木在石林风景区石漠化环境中分布广泛, 对植被构建起到重要作用。清香木在栽植植被中的生态位宽度与地带性自然植被中的生态位宽度接近, 可以推测清香木在两种环境中的地位一致, 对资源的利用能力接近。尽管3种植被类型所在样地的岩石裸露率相似, 但是研究发现栽植植被与地带性自然植被样地中起到“避难所”作用的溶痕生境(溶坑,溶槽等)更加丰富, 清香木的实生苗与萌生苗多数储存在溶痕生境中, 利于其进行快速更新[24], 充分利用环境资源, 拓展其生态位宽度。
生态位重叠是衡量种间生态相似性以及竞争性的重要指标。同时, 生态位重叠与物种的生态学特性以及竞争强弱有密切关系[25]。一般来说, 重要值越大, 生态位的宽度也越大, 分布的范围较广, 因而与其他物种间的生态位重叠值较大[26]。但是, 在栽植植被中, 生态位宽度与生态位重叠不存在类似关联, 生态位较宽的植物物种间的生态位重叠值普遍低于0.5, 这与石林地带性自然植被中的生态位重叠特征接近, 也与黄甫昭[27]对广西弄岗喀斯特季雨林顶级群落的生态位重叠研究结果相似, 这可能与喀斯特地区石芽出露的特殊地质条件造成物种分布的不连续性有关, 同时也表明栽植植被主要优势种群之间竞争不激烈, 植被较为稳定。
生态位重叠为零则表明种对的生态习性和对生态因子的要求完全不同, 种对之间不存在竞争关系。栽植植被及飞播植被中木本植物种群生态位重叠为0的种对比例远高于地带性自然植被中生态位重叠值为0的种对比例, 一方面说明人工修复植被中种群生态位分化明显, 生态习性不同, 这可能与石漠化地区浅薄土层与岩石镶嵌分布而形成的生境异质性有关[28];另一方面可能是人工修复中综合考虑地质景观特征而进行植物景观设计, 在特定的区域形成了特定的植物组合。
3.3 石漠化人工修复方式对石漠化地区植被修复启示当前石漠化生态修复的目标已从遏制石漠化面积扩张, 增加植被覆盖度转向提升生态系统服务, 增强石漠化生态修复的可持续性[29], 包括生态效果与景观效果的可持续性。在石漠化植被修复的初期, 应以提升生态效果为目标, 选择环境条件一致或相似的地带性自然植被为参考, 以乡土植物中生态位宽度大的物种为先锋种, 生态位重叠相对较小的物种作为伴生种[27]进行栽植, 考虑各种群在形态、结构和生理等方面互相弥补的特征[30]构建植物群落, 保证群落多样性指数维持在合理的范围之内, 充分利用环境资源。例如, 在云南地区可采用川滇无患子+清香木—白刺花, 黄连木+清香木—沙针+铁仔等群落配置方式, 营造近自然群落。当植被盖度达到一定程度, 可以进一步调整群落结构。本研究以栽植植被中高观赏价值的木本植物群落的物种组成和群落结构为参考, 提出基于提升生物多样性, 突出喀斯特地区景观效果的植被修复建议:从岩石形态考虑, 在高大的石柱单体周围配置常绿或者色彩变化丰富的木本植物, 例如清香木与黄连木等, 在低矮的溶痕生境中选择高度适中与形态多样的植物种类;结合乡土植物物候变化规律, 通过花色, 叶色和果色的季相变化来营造植物景观的时空变化之美, 在保证生态功能的基础上提升景观价值, 使得喀斯特石漠化治理地区先变“绿”, 再变“美”[31]。
| [1] |
兰安军, 张百平, 熊康宁, 安裕伦. 黔西南脆弱喀斯特生态环境空间格局分析. 地理研究, 2003, 22(6): 733-741. DOI:10.3321/j.issn:1000-0585.2003.06.008 |
| [2] |
宋同清, 彭晚霞, 曾馥平, 王克林, 覃文更, 谭卫宁, 刘璐, 杜虎, 鹿士杨. 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报, 2010, 34(3): 298-308. DOI:10.3773/j.issn.1005-264x.2010.03.007 |
| [3] |
曾馥平, 彭晚霞, 宋同清, 王克林, 吴海勇, 宋希娟, 曾昭霞. 桂西北喀斯特人为干扰区植被自然恢复22年后群落特征. 生态学报, 2007, 27(12): 5110-5119. DOI:10.3321/j.issn:1000-0933.2007.12.020 |
| [4] |
宋同清, 彭晚霞, 曾馥平, 王克林, 曹洪麟, 李先琨, 覃文更, 谭卫宁, 刘璐. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征. 生物多样性, 2010, 18(4): 355-364. |
| [5] |
文丽, 宋同清, 杜虎, 王克林, 彭晚霞, 曾馥平, 曾昭霞, 何铁光. 中国西南喀斯特植物群落演替特征及驱动机制. 生态学报, 2015, 35(17): 5822-5833. |
| [6] |
杜文鹏, 闫慧敏, 甄霖, 胡云锋. 西南岩溶地区石漠化综合治理研究. 生态学报, 2019, 39(16): 5798-5808. |
| [7] |
Tang C Q, Li Y H, Zhang Z Y. Species diversity patterns in natural secondary plant communities and man-made forests in a subtropical mountainous Karst Area, Yunnan, SW China. Mountain Research and Development, 2010, 30(3): 244-251. |
| [8] |
鹿士杨, 彭晚霞, 宋同清, 曾馥平, 杜虎, 王克林. 喀斯特峰丛洼地不同退耕还林还草模式的土壤微生物特性. 生态学报, 2012, 32(8): 2390-2399. |
| [9] |
Zhang M Y, Wang K L, Liu H Y, Zhang C H, Yue Y M, Qi X K. Effect of ecological engineering projects on ecosystem services in a karst region:a case study of northwest Guangxi, China. Journal of Cleaner Production, 2018, 183: 831-842. DOI:10.1016/j.jclepro.2018.02.102 |
| [10] |
苏维词, 朱文孝, 熊康宁. 贵州喀斯特山区的石漠化及其生态经济治理模式. 中国岩溶, 2002, 21(1): 19-24. DOI:10.3969/j.issn.1001-4810.2002.01.005 |
| [11] |
Tang C Q, Hou X L, Gao K, Xia T Y, Duan C Q, Fu D G. Man-made versus natural forests in mid-Yunnan, Southwestern China:plant diversity and initial data on water and soil conservation. Mountain Research and Development, 2007, 27(3): 242-249. |
| [12] |
王克林, 岳跃民, 陈洪松, 吴协保, 肖峻, 祁向坤, 张伟, 杜虎. 喀斯特石漠化综合治理及其区域恢复效应. 生态学报, 2019, 39(20): 7432-7440. |
| [13] |
Miyawaki A. Creative ecology:restoration of native forests by native trees. Plant Biotechnology, 1999, 16(1): 15-25. |
| [14] |
李玉辉, 冯正清, 俞筱押, 马遵平. 云南石林公园植被重大变化与意义. 中国岩溶, 2005, 24(3): 212-219. DOI:10.3969/j.issn.1001-4810.2005.03.009 |
| [15] |
李玉辉. 从中国南方喀斯特, 看中国世界自然遗产地如何服务国家新战略. 遗产与保护研究, 2016, 1(1): 95-100. DOI:10.3969/j.issn.2096-0913.2016.01.023 |
| [16] |
Tilman D. Causes, consequences and ethics of biodiversity. Nature, 2000, 405(6783): 208-211. |
| [17] |
Smith A B, Godsoe W, Rodríguez-Sánchez F, Wang H H, Warren D. Niche estimation above and below the species level. Trends in Ecology & Evolution, 2019, 34(3): 260-273. |
| [18] |
Vannette R L, Fukami T. Historical contingency in species interactions:towards niche-based predictions. Ecology Letters, 2014, 17(1): 115-124. |
| [19] |
沈有信, 刘文耀, 李玉辉, 崔建武. 滇中喀斯特山地半湿润常绿阔叶林的群落生态学研究. 广西植物, 2005, 25(4): 321-326. DOI:10.3969/j.issn.1000-3142.2005.04.007 |
| [20] |
俞筱押, 李玉辉, 杨光荣. 石林地质公园不同群落类型植物果实组成与种子散布特征. 植物生态学报, 2018, 42(6): 663-671. |
| [21] |
兰斯安, 宋敏, 曾馥平, 宋同清, 彭晚霞, 杜虎, 韩畅, 陈莉, 覃文更. 木论喀斯特森林木本植物多样性垂直格局. 生态学报, 2016, 36(22): 7374-7383. |
| [22] |
Wang Z H, Duan C Q, Chen M, Wang K Q. The ecological-hydrological characteristics of the three manmade forest communities in the central Yunnan province//Proceedings of the 12th International Soil Conservation Organization Conference. Beijing, China: Tsinghua University, 2002: 541-550.
|
| [23] |
Gelviz-Gelvez S M, Pavón N P, Illoldi-Rangel P, Ballesteros-Barrera C. Ecological niche modeling under climate change to select shrubs for ecological restoration in Central Mexico. Ecological Engineering, 2015, 74: 302-309. DOI:10.1016/j.ecoleng.2014.09.082 |
| [24] |
俞筱押, 李玉辉. 滇石林喀斯特植物群落不同演替阶段的溶痕生境中木本植物的更新特征. 植物生态学报, 2010, 34(8): 889-897. DOI:10.3773/j.issn.1005-264x.2010.08.001 |
| [25] |
Tilman G D. Plant dominance along an experimental nutrient gradient. Ecology, 1984, 65(5): 1445-1453. |
| [26] |
刘润红, 常斌, 荣春艳, 姜勇, 杨瑞岸, 刘星童, 曾惠帆, 傅桂焕. 漓江河岸带枫杨群落主要木本植物种群生态位. 应用生态学报, 2018, 29(12): 3917-3926. |
| [27] |
黄甫昭, 李冬兴, 王斌, 丁涛, 向悟生, 文淑均, 郭屹立, 李先琨. 喀斯特季节性雨林优势种群生态位特征及其对石漠化地区植被修复的启示. 广西科学, 2018, 25(5): 599-610. |
| [28] |
倪隆康, 顾大形, 何文, 黄玉清, 陈中义. 岩溶区植物生态适应性研究进展. 生态学杂志, 2019, 38(7): 2210-2217. |
| [29] |
Liao C J, Yue Y M, Wang K L, Fensholt R, Tong X W, Brandt M. Ecological restoration enhances ecosystem health in the karst regions of southwest China. Ecological Indicators, 2018, 90: 416-425. DOI:10.1016/j.ecolind.2018.03.036 |
| [30] |
陈林, 辛佳宁, 苏莹, 李月飞, 宋乃平, 王磊, 杨新国, 卞莹莹, 田娜. 异质生境对荒漠草原植物群落组成和种群生态位的影响. 生态学报, 2019, 39(17): 6187-6205. |
| [31] |
李坤, 董丽, 陈强, 陈德生. 云南石漠化地区植被生态修复群落特征研究. 中国园林, 2017, 33(11): 41-46. DOI:10.3969/j.issn.1000-6664.2017.11.008 |