 2020, Vol. 40
2020, Vol. 40文章信息
- 邓健, 种玉洁, 贾小敏, 焦子怡, 任成杰, 韩新辉, 杨改河
- DENG Jian, CHONG Yujie, JIA Xiaomin, JIAO Ziyi, REN Chengjie, HAN Xinhui, YANG Gaihe
- 黄土高原子午岭林区典型树种叶片N、P再吸收特征
- Leaf N and P resorption characteristics of typical tree species in the Ziwuling forest area on Loess Plateau
- 生态学报. 2020, 40(11): 3698-3705
- Acta Ecologica Sinica. 2020, 40(11): 3698-3705
- http://dx.doi.org/10.5846/stxb201906201294
-
文章历史
- 收稿日期: 2019-06-20
- 网络出版日期: 2020-03-31





2. 陕西省红枣重点实验室(延安大学), 延安 716000;
3. 西北农林科技大学农学院, 杨凌 712100
2. Shaanxi Key Laboratory of Chinese Jujube, Yan'an University, Yan'an 716000, China;
3. College of Agronomy, Northwest Agriculture and Forestry University, Yangling 712100, China
植物养分再吸收(Nutrient resorption)是指植物枝叶在枯落前将其中的营养成分转移到生长组织中的过程[1]。这一过程有效延长了养分在植物体内的留存时间, 不仅降低了植物对土壤养分的依赖程度, 有助于维持植物体内化学计量平衡;而且减少了生态系统凋落物分解后的养分淋溶, 减缓了生态系统养分损失[2-3]。在众多器官中, 叶片是植物的重要光合器官和营养器官, 且具有生长迅速, 周转快的特点, 植物叶片的养分再吸收被认为是植物适应环境养分限制的重要机制, 也是增强竞争力、提高养分利用效率和生产力的重要策略[3-4]。因此, 研究不同类型植物叶片养分再吸收特征有助于揭示植物对环境的响应特征[5]。
氮(N)和磷(P)是植物生长代谢的必需元素, 也是陆地生态系统主要限制元素[6], 而植物叶片N、P再吸收率(Nitrogen resorption efficiency, NRE和Phosphorus resorption efficiency, PRE)能很好地揭示植物对N或P限制的适应性, 因此受到研究者的广泛关注[3-7]。Vergutz等估计全球陆地植物大约有62.1%的N和64.9%的P通过再吸收作用回到植物活体组织中[8], 但不同演替阶段、不同种类[3-4]植物的N、P再吸收率差异较大[9-10], 这有可能是由于植物本身遗传特性[11-12]或土壤养分等环境因子的影响[13-14], 例如Han等[15]提出的“相对再吸收假说”认为当植物生长过程受到某种元素限制时会对该元素具有较高的再吸收效率。基于生态化学计量学理论的植物和土壤C:N:P养分计量比能直观反映养分的相对含量和限制状况, 如叶片N:P常被用来指示植物受到N(N:P < 14)或P限制(N:P>16)[16]。因此, 研究不同植被类型和演替阶段植物叶片N、P再吸收特征及其与养分化学计量比的关系, 有助于更好地揭示植物对养分限制环境的适应性, 对于阐明特定区域植被群落演替机理也具有重要意义[17]。
子午岭林区是黄土高原保存较好的连片天然次生林区, 辽东栎(Quercus wutaishansea Mary)林和油松(Pinus tabuliformis Carr.)林为该区演替过程的亚顶级群落, 白桦(Betula platyphylla Suk.)和山杨(Populus davidiana Dode)林是该区演替的先锋乔木群落[18];另外区域内还分布较大面积的人工刺槐(Robinia pseudoacacia L.)林和人工侧柏(Platycla dusorientalis (L.) Franco)林[9]。此前学者针对子午岭林区植被演替和物种多样性[19]、土壤养分、酶活性和植物养分[20]等方面进行了研究, 同时指出该区不同植被生长可能受到N或P养分限制[9, 21], 然而面对养分限制条件, 不同类型和演替阶段树种的养分再吸收特征仍然不够明确。因此, 本研究选取子午岭林区典型植被树种, 测定其叶片和林下土壤养分含量, 计算生态化学计量学指标, 重点分析不同植被类型和演替阶段树种叶片N、P养分再吸收特征及其与养分指标的关系, 以期揭示子午岭林区典型树种对养分限制环境的适应性, 研究可为黄土高原地区植被恢复树种选择提供理论依据。
1 材料和方法 1.1 研究区概况研究区域位于陕西省富县牛武镇附近(109°36′13″E, 36°05′24″N), 为典型的黄土丘陵沟壑区, 属暖温带半湿润气候, 海拔1115—1342 m, 年均降水量550.3 mm, 年均温9.8℃, 无霜期175 d, 土壤为原生或次生黄土。植被以天然次生林为主, 主要树种有山杨、白桦、辽东栎、油松等;少量人工林分布于缓坡或沟谷, 以刺槐、侧柏为主。
1.2 试验设计和采样本研究选取立地条件相近的天然白桦和山杨林、辽东栎和油松林、人工刺槐和侧柏林为研究对象, 分别代表演替初期、演替后期和人工植被林地, 每种林地选择3块样地, 每个样地内随机设置3个20 m×20 m的样方进行采样和调查。为保证气候条件的一致性, 不同样地间距在5 km范围内。样地基本信息见表 1。
| 样地 Sample land |
海拔 Elevation/m |
坡度 Slope/(°) |
坡向 Aspect/(°) |
容重 Bulk density/ (g/cm3) |
平均胸径 Diameter at breast height/cm |
平均树高 Average tree height/m |
林下主要植物 Mainly undergrowthplans |
| 刺槐 Robinia pseudoacacia L. |
1195 | 27 | 40 | 1.15 | 28.64 | 7.65 | 蛇莓Duchesnea indica (Andr.) Focke 刺儿菜 Cirsium setosum (Willd.) MB. |
| 侧柏 Platycladus orientalis (L.) Franco |
1241 | 30 | 45 | 1.14 | 22.36 | 7.03 | 陕西荚蒾Viburnum schensianum Maxim. 翻白草 Potentilla discolorBge. |
| 白桦 Betula platyphylla Suk |
1194 | 22 | 50 | 1.06 | 38.55 | 8.36 | 毛樱桃 Cerasus tomentosa (Thunb.) Wall. 大披针薹草Carex lanceolate Boott |
| 山杨 Populus davidiana Dode |
1173 | 16 | 284 | 1.04 | 36.85 | 9.86 | 冻绿RhamnusutilisDecne. 大披针薹草Carexlanceolate Boott |
| 油松 Pinus tabuliformis Carr |
1201 | 35 | 310 | 1.01 | 35.64 | 8.65 | 毛樱桃Cerasus tomentosa (Thunb.) Wall. 大披针薹草Carexlanceolate Boott |
| 辽东栎 Quercus wutaishansea Mary |
1267 | 32 | 60 | 1.01 | 39.67 | 7.66 | 麻叶绣线菊Spiraea cantoniensis Lour. 山罗花Melampyrum roseum Maxim. |
2016年7月中旬在每个植被样方内随机选择10株生长良好的建群种植株, 测量其胸径并标记, 在每株植物东南西北四个方位摘取树冠上、中、下部外侧枝条上受光良好、深绿色、无病虫害的成熟叶片(侧柏和油松摘取当年新生叶片), 每株采集叶片约50 g, 同一样方内的叶片混匀后采用四分法取100 g左右装入牛皮纸袋带回实验室, 105℃杀青30 min, 然后60℃烘干至恒重, 用粉碎机粉碎后过100目筛保存用于养分测定;10月底在标记的植株上采集完全变黄但还未凋落的叶片, 采样方法与成熟叶方法一样。同时, 7月中旬每个样方内沿对角线选择5个采样点, 去除植物残体和表层枯落物后用直径为4 cm的土钻采集0—10 cm林下土壤, 将同一样方内5个样点的土样去除杂物、混匀、风干、研磨过100目筛后保存, 用于土壤养分测定。每个样方内随机选择5个1 m×1 m的小样方调查林下草本植被。
1.3 样品分析土壤和植物叶片有机碳含量均采用重铬酸钾-外加热容量法测定;土壤全氮采用浓硫酸-高氯酸加热消煮后用全自动凯氏定氮仪(KDY-9830)测定;叶片全氮采用浓硫酸-H2O2加热消煮后用全自动凯氏定氮仪测定;土壤和叶片全磷经上述消煮后用钼酸铵比色法测定;土壤样品采集时用环刀法测定土壤容重[22]。
1.4 数据处理和统计分析植物叶片养分再吸收率计算公式为[7]:
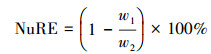
|
式中NuRE(Nutrients resorption efficiency)为叶片N或P的再吸收率;w1为凋落叶N或P含量(g/kg);w2为成熟叶N或P含量(g/kg)。
土壤和叶片养分生态化学计量比采用质量比;研究采用单因素方差分析(One-way ANOVA)检验各指标不同处理间的差异, 采用Duncan法进行多重比较, 显著性水平为P < 0.05;采用冗余分析(redundancy analysis, RDA)研究不同植物N、P再吸收率与养分指标、生态化学计量学指标间的关系;使用SAS(V9.0, SAS Institute Inc., North Carolina, USA)进行数据统计和计算, Origin 2016(OriginLab Corporation, Northampton MA, USA)用于作图, 图中误差线为标准误差。
2 结果 2.1 不同树木林地土壤C、N、P含量和计量学特征土壤C、N、P含量和计量比在不同林地之间具有显著差异, 其中C、N含量和C:N:P计量比均为刺槐林最低, 辽东栎林最高(P < 0.05), 总体表现为演替后期林地(辽东栎和油松)>演替前期林地(山杨和白桦)>人工林(侧柏和刺槐)(P < 0.05)。刺槐林土壤P含量显著高于其他林地, 侧柏和辽东栎林土壤P含量显著低于其他林地(P < 0.05)。
| 样地Sample land | C/(g/kg) | N/(g/kg) | P/(g/kg) | C:N | C:P | N:P |
| 刺槐 Robinia pseudoacacia L. | 9.62±2.08d | 0.89±0.07d | 0.60±0.04a | 12.41±1.81bc | 42.23±11.46c | 3.34±0.45c |
| 侧柏Platycladus orientalis (L.) Franco | 15.50±1.63c | 1.51±0.08c | 0.42±0.07c | 11.94±0.94c | 98.43±15.91b | 8.22±0.88b |
| 白桦Betula platyphylla Suk | 22.18±1.67b | 1.87±0.10b | 0.49±0.05b | 13.87±1.47b | 117.98±19.10b | 8.46±0.48b |
| 山杨Populus davidiana Dode | 21.91±1.10b | 1.84±0.10b | 0.49±0.08b | 13.96±1.11b | 118.48±17.16b | 8.51±1.14b |
| 油松Pinus tabuliformis Carr | 22.97±3.46b | 1.64±0.05bc | 0.51±0.02b | 16.39±2.02a | 116.18±13.86b | 7.17±0.43b |
| 辽东栎Quercus wutaishansea Mary | 28.95±1.05a | 2.05±0.24a | 0.44±0.04c | 16.69±2.21a | 171.44±16.75a | 10.5±1.08a |
| 同列不同小写字母表示不同树种间存在显著差异(P < 0.05) | ||||||
不同树种鲜叶和凋落叶C、N、P含量存在显著差异(P < 0.05)。其中白桦和油松鲜叶和凋落叶C含量均显著高于其他树种(P < 0.05)。刺槐鲜叶和凋落叶N含量最高, 白桦和辽东栎次之, 其余树种都较低(P < 0.05)。刺槐、侧柏和辽东栎鲜叶P含量显著高于其他三个树种(P < 0.05), 但凋落叶P含量为侧柏最高, 油松最低, 其他树种之间差异不显著。
不同树种鲜叶和凋落叶C:N:P计量学特征也存在显著差异(P < 0.05)。其中鲜叶C:N表现为侧柏、山杨和油松显著高于其他三种树种(P < 0.05);凋落叶C:N油松最高, 刺槐最低, 其他树种之间差异不显著。白桦、山杨、油松的鲜叶C:P显著高于刺槐、侧柏和辽东栎(P < 0.05);油松凋落叶C:P显著高于其他树种, 刺槐、侧柏两种人工林凋落叶C:P显著低于其他4种树种(P < 0.05)。刺槐鲜叶和凋落叶N:P均显著高于其他树种(P < 0.05), 但侧柏鲜叶和凋落叶N:P显著低于其他树种(P < 0.05), 白桦、山杨、油松和辽东栎之间叶片N:P差异不显著。

|
| 图 1 不同树种叶片C、N、P含量和计量比 Fig. 1 Leaf C, N, P contents and stoichiometry ratios in different forests 同一指标标注不同小写字母表示树种之间存在显著差异(P < 0.05) |
不同树种叶片NRE和PRE之间均具有显著的差异。其中辽东栎叶片NRE最高, 为43.34%, 刺槐(40.99%)和油松(36.78%)次之, 山杨最低, 仅为17.18%(P < 0.05)。叶片PRE表现为辽东栎最高(58.12%), 油松(46.00%)、刺槐(42.51%)和侧柏(37.14%)次之, 山杨(30.73%)和白桦(27.13%)最低的趋势(P < 0.05)。对不同演替阶段和类型的树种养分再吸收特征分析发现, 叶片NRE和PRE均表现为演替后期林地(辽东栎和油松)>人工林(侧柏和刺槐)>演替前期林地(山杨和白桦)(P < 0.05)。除白桦外, 其他树种均表现为叶片PRE>NRE。

|
| 图 2 不同树种叶片N和P再吸收率(NRE和PRE)特征 Fig. 2 Leaf N and P resorption efficiencies of different tree species NRE:氮再吸收率Nitrogen resorption efficiency;PRE:磷再吸收率Phosphorus resorption efficiency; 同一指标标注不同小写字母表示树种之间存在显著差异(P < 0.05) |
不同树种间NRE和PRE变化规律与土壤和植物叶片养分和计量比之间存在一定的关系。植物叶片C、N、P含量和计量比对叶片N、P再吸收率变化的综合解释率达到68.53%, 对NRE和PRE的解释率分别为56.96%和58.45%(图 3)。其中叶片TN、C:N和N:P与NRE之间存在显著相关性, 叶片TP、C:P和N:P与PRE之间存在显著相关性(P < 0.05)。土壤C、N、P含量和计量比对叶片N、P再吸收率变化的综合解释率为47.58%, 对NRE和PRE的解释率分别为43.04%和41.55%(图 3)。其中仅土壤C:N与叶片NRE之间存在显著相关性, 土壤C:N和C:P与叶片PRE之间存在显著相关性(P < 0.05)。叶片养分和计量比的综合解释率和指标单项解释率均高于土壤。

|
| 图 3 不同树种叶片N和P再吸收率与养分和计量比的冗余分析 Fig. 3 Redundancy analysis of leaf N, P resorption efficiencies and nutrients concentration and its stoichiometry ratios in different tree species TC:碳含量C concentration;TN:氮含量N concentration;TP:磷含量P concentration;下标Leaf或Soil分别表示该指标为叶片或土壤;VEP和VEN分别表示该指标对NRE或PRE的解释率;TVE表示叶片(下标leaf)或土壤(下标soil)指标对NRE(下标-N)或PRE(下标-P)的综合解释率;图中未标注解释率的因子说明其对NRE和PRE的解释不显著; * P < 0.05;** P < 0.01 |
土壤-植物系统之间的C、N、P养分循环和转化过程是陆地生态系统物质循环的基础, 但不同类型植被与土壤之间的关系存在差异[23-24]。与人工林相比, 相同条件下的天然林和次生林植被更容易促进土壤养分的积累[25-26]。本研究中除土壤P含量外, 土壤C、N和C:N:P计量比均表现为次生林大于人工林, 且演替后期林地(辽东栎和油松)大于演替前期林地(白桦和山杨), 进一步支持了以上结论。
植物成熟叶片养分含量及其计量比是反映植物生长状况的重要指标, 主要决定于植物物种本身特性和生长阶段[4, 27], 但也受土壤养分含量及其平衡关系的影响[28]。本研究中各树种成熟叶C、N、P含量变化范围分别为369.72—476.26 g/kg、11.62—28.94 g/kg和2.48—4.74 g/kg, 其中C、N含量接近全球陆地植物叶片C(464 g/kg)[29]和N(20.09 g/kg)[30]平均含量, 而P含量高于全球平均水平(1.77 g/kg)[31];不同树种成熟叶片养分含量存在显著差异, 其中叶片C、N、P含量最高的树种分别为白桦、刺槐和辽东栎。Güsewell[16]研究提出将叶片N:P>16、16>N:P>14和N:P < 14作为指示植物生长受P限制、N和P共同限制和N限制的阈值。根据这个阈值, 本研究中仅刺槐叶片N:P为17.72大于16, 白桦叶片N:P为14.83, 略高于14但小于16, 说明二者分别可能受到P限制或N、P共同限制;而其他树种叶片N:P均远小于14, 生长可能受到N限制。这与此前研究认为黄土高原地区刺槐林生长受到P限制[27]而油松、辽东栎、侧柏等受到N限制[9, 21]的结论一致, 说明子午岭人工和次生林树种均不同程度受到N或P养分的限制。
植物凋落叶是养分向土壤归还的重要途径, 通常植物叶片在凋落前会将一部分N、P养分转移到枝干从而造成了凋落叶N、P含量低于新鲜叶片N、P含量[3, 32], 本研究中各树种凋落叶N、P含量变化范围分别为7.38—17.07 g/kg和1.33—2.66 g/kg, 均低于成熟叶(P < 0.05), 符合这一规律。但凋落叶C含量为404.43—497.65 g/kg, 较成熟叶平均增加了5.29%, 主要因为C是构成植物叶片的结构性元素, 凋落叶片水分下降和其他养分再吸收会导致其C含量相对增加。由于凋落叶的分解主要依靠微生物进行, 而凋落物N、P丰富时有利于微生物(尤其是细菌)活动, 进而促进凋落物快速分解和养分释放[33], 通常将凋落物C:N < 40、C:P < 480认为是有利于分解的临界值[34-35], 本研究中仅刺槐凋落叶C:N(27.71)、侧柏凋落叶C:P(400.54)低于临界值, 其他树种凋落叶计量比均高于临界值, 说明子午岭地区次生林树种凋落叶分解受到N、P养分的限制, 有利于C在枯落物层的积蓄, 这与张海鑫等的研究结果一致[9]。
3.2 不同植被养分再吸收率与叶片和土壤养分的关系植物从土壤中吸收各类养分并分配到不同器官中积累或参与各种生命活动, 当叶片衰老凋落前又会对其中的N、P等进行再吸收, 从而延长养分在植物体内的留存时间[3]。本研究中不同植被叶片N、P再吸收率分别为17.18%—43.34%和27.13%—58.12%, 说明这些植被对P的再吸收能力高于N, 但均远低于Vergutz等综合分析得到的全球陆地森林生态系统叶片N、P再吸收率(分别为62.10%和64.90%)[8], 这可能是由于研究选取的树种差异或者研究区域环境条件的差异导致[4], 同时本研究计算再吸收率时未考虑叶片枯落后的质量损失矫正, 这也会使计算得到的再吸收率偏低[8]。不同类型植被N、P再吸收率均表现为演替后期植被>人工植被>演替前期植被(P < 0.05), 尤其是辽东栎对N和P的再吸收率均最高, 这与张海鑫等研究发现子午岭地区辽东栎对N的再吸收不显著的结果不一致, 可能是由于采样时间间隔不同导致[9]。虽然本研究中不同植物生长条件和年限基本一致, 但其对N和P的再吸收能力表现出较大差异, 这可能是由于植物本身遗传特性决定了其对养分的再吸收[11-12]。值得注意的是, 本研究中演替后期的辽东栎和油松对N、P的再吸收率显著高于演替前期的白桦和杨树, 由于不同演替阶段的群落形成是物种对资源竞争的结果, 我们推测更高的养分再吸收能力可能有助于油松、辽东栎在养分相对贫瘠的条件下生长, 进而在演替后期形成稳定的顶级群落, 但这一推测还需要进一步研究。
虽然Killingbeck认为养分再吸收能力是植物本身的固有特性[12], 但也有研究认为植物的养分再吸收率可能还受土壤养分条件、成熟叶片养分和凋落物养分的单独或共同影响[8, 11]。本研究中叶片的N、P含量和C:N:P计量比与N、P再吸收率表现出显著的相关性, 对不同植物N、P再吸收的解释率分别达到56.96%和58.45%;土壤养分仅C:N和C:P与植物N、P再吸收率有显著关系, 且总体解释率低于叶片, 这一结果反映出两个问题:(1)本研究中叶片养分对植物N、P再吸收率的影响大于土壤养分, 这符合Vergutz等[8]得到的成熟叶片养分对植物再吸收有显著影响的结论;(2)无论是叶片和还是土壤, 均表现为计量学指标与N、P再吸收率的关系比养分含量更加显著, 这可能是由于计量比能更好地反映植物或土壤养分的相对含量, 而Han等[15]提出的“相对再吸收假说”就认为植物对某种养分的再吸收率会受到该养分相对缺乏状况的显著影响, 例如指示植物的养分限制状况的叶片N:P与N、P养分再吸收率均具有显著相关关系。因此, 子午岭林区不同植物对N、P的再吸收可能是其适应N、P限制环境的重要策略。当然, 土壤-植物系统的养分互作和循环是一个复杂的过程, 本研究仅从叶片养分再吸收特征及其与环境养分条件关系的角度进行了分析, 但针对植物养分再吸收机理、不同器官养分再吸收能力等方面的研究并未涉及, 未来依托分子标记、同位素示踪等手段的深入研究能够更进一步揭示该过程的内在机理。
| [1] |
Aerts R. Nutrient resorption from senescing leaves of perennials:are there general patterns?. Journal of Ecology, 1996, 84(4): 597-608. DOI:10.2307/2261481 |
| [2] |
Lü X T, Reed S C, Yu Q, Han X G. Nutrient resorption helps drive intra-specific coupling of foliar nitrogen and phosphorus under nutrient-enriched conditions. Plant and Soil, 2016, 398(1/2): 111-120. |
| [3] |
陆姣云, 段兵红, 杨梅, 杨晗, 杨惠敏. 植物叶片氮磷养分重吸收规律及其调控机制研究进展. 草业学报, 2018, 27(4): 178-188. |
| [4] |
刘宏伟, 刘文丹, 王微, 柴捷, 陶建平. 重庆石灰岩地区主要木本植物叶片性状及养分再吸收特征. 生态学报, 2015, 35(12): 4071-4080. |
| [5] |
Liang D F, Zhang JJ, Zhang S T. Patterns of nitrogen resorption in functional groups in a Tibetan alpine meadow. Folia Geobotanica, 2015, 50(3): 267-274. DOI:10.1007/s12224-015-9217-9 |
| [6] |
Campo J. Shift from ecosystem P to N limitation at precipitation gradient in tropical dry forests at Yucatan, Mexico. Environmental Research Letters, 2016, 11(9): 095006. DOI:10.1088/1748-9326/11/9/095006 |
| [7] |
Zhang H, Wang J, Wang J, Guo Z, Wang G G, Zeng D, Wu T. Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. Forest Ecology and Management, 2018, 430: 445-450. DOI:10.1016/j.foreco.2018.08.037 |
| [8] |
Vergutz L, Manzoni S, Porporato A, Novais R F, Jackson R B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecological Monographs, 2012, 82(2): 205-220. DOI:10.1890/11-0416.1 |
| [9] |
张海鑫, 曾全超, 安韶山, 白雪娟, 王宝荣. 黄土高原子午岭林区主要林分生态化学计量学特征. 自然资源学报, 2017, 32(06): 1043-1052. |
| [10] |
申奥, 朱教君, 闫涛, 卢德亮, 杨凯. 辽东山区主要阔叶树种叶片养分含量和再吸收对落叶时间的影响. 植物生态学报, 2018, 42(5): 573-584. |
| [11] |
Luyssaert S, Staelens J, De Schrijver A. Does the commonly used estimator of nutrient resorption in tree foliage actually measure what it claims to?. Oecologia, 2005, 144(2): 177-186. DOI:10.1007/s00442-005-0085-5 |
| [12] |
Killingbeck K T. Nutrients in Senesced Leaves:Keys to the Search for Potential Resorption and Resorption Proficiency. Ecology, 1996, 77(6): 1716-1727. DOI:10.2307/2265777 |
| [13] |
宁秋蕊, 李守中, 姜良超, 陶晶晶, 陈涵睿, 刘聪, 杨贤宇. 植物叶片养分再吸收特征及其影响因子. 应用与环境生物学报, 2017, 23(05): 811-817. |
| [14] |
Yan T, Zhu J, Yang K. Leaf nitrogen and phosphorus resorption of woody species in response to climatic conditions and soil nutrients:a meta-analysis. Journal of Forestry Research, 2018, 29(4): 905-913. DOI:10.1007/s11676-017-0519-z |
| [15] |
Han W, Tang L, Chen Y, Fang J. Relationship between the relative limitation and Resorption efficiency of nitrogen vs phosphorus in woody plants. PLoS One, 2014, 9(4): e94515. DOI:10.1371/journal.pone.0094515 |
| [16] |
Güsewell S. N:P ratios in terrestrial plants:variation and functional significance. New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [17] |
Yan T, Lü X T, Zhu JJ, Yang K, Yu L Z, Gao T. Changes in nitrogen and phosphorus cycling suggest a transition to phosphorus limitation with the stand development of larch plantations. Plant and Soil, 2018, 422(1/2): 385-396. |
| [18] |
王凯博, 陈美玲, 秦娟, 刘勇, 安慧, 上官周平. 子午岭植被自然演替中植物多样性变化及其与土壤理化性质的关系. 西北植物学报, 2007, 27(10): 2089-2096. DOI:10.3321/j.issn:1000-4025.2007.10.025 |
| [19] |
范玮熠, 王孝安, 郭华, 王世雄, 段仁燕. 陕西子午岭森林群落的物种多样性研究. 陕西师范大学学报:自然科学版, 2014, 42(3): 59-66. |
| [20] |
张海鑫, 曾全超, 安韶山, 王宝荣, 白雪娟. 子午岭典型植被凋落叶-土壤养分与酶活性特征. 生态学报, 2018, 38(07): 2262-2270. |
| [21] |
汪宗飞, 郑粉莉. 黄土高原子午岭地区人工油松林碳氮磷生态化学计量特征. 生态学报, 2018, 38(19): 6870-6880. |
| [22] |
鲍士旦. 土壤农化分析. 北京: 中国农业出版社, 2007.
|
| [23] |
贺纪正, 陆雅海, 傅伯杰. 土壤生物学前沿. 北京: 科学出版社, 2015.
|
| [24] |
艾泽民.黄土丘陵区撂荒演替典型草本植物——土壤反馈研究[D].杨凌: 中国科学院大学(中国科学院教育部水土保持与生态环境研究中心), 2018.
|
| [25] |
张克胜, 尚晴, 刘彦春, 韩艳婷, 马远超, 管勇, 琚煜熙. 中国不同气候带人工林与天然林的土壤呼吸差异. 生态科学, 2017, 36(06): 49-56. |
| [26] |
陈小花, 杨青青, 余雪标, 陈宗铸, 杨琦, 雷金睿. 热带海岸防护林土壤活性有机碳分布特征:人工林与天然次生林的比较. 热带作物学报, 2017, 38(03): 494-499. DOI:10.3969/j.issn.1000-2561.2017.03.018 |
| [27] |
Schreeg L A, Santiago L S, Wright S J, Turner B L. Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology, 2016, 95(8): 2062-2068. |
| [28] |
Elser J J, Fagan W F, Denno R F, Dobberfuhl D R, Folarin A, Huberty A, Interlandi S, Kilham S S, McCauley E, Schulz K L, Siemann E H, Sterner R W. Nutritional constraints in terrestrial and freshwater food webs. Nature, 2000, 408(6812): 578-580. DOI:10.1038/35046058 |
| [29] |
Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006. DOI:10.1073/pnas.0403588101 |
| [30] |
Herbert D A, Williams M, Rastetter E B. A model analysis of N and P limitation on carbon accumulation in Amazonian secondary forest after alternate land-use abandonment. Biogeochemistry, 2003, 65(1): 121-150. DOI:10.1023/A:1026020210887 |
| [31] |
马任甜, 安韶山, 黄懿梅. 黄土高原不同林龄刺槐林碳、氮、磷化学计量特征. 应用生态学报, 2017, 28(09): 2787-2793. |
| [32] |
江大龙, 徐侠, 阮宏华. 植物养分重吸收及其影响研究进展. 南京林业大学学报:自然科学版, 2017, 41(1): 183-188. |
| [33] |
陆晓辉, 丁贵杰, 陆德辉. 人工调控措施下马尾松凋落叶化学质量变化及与分解速率的关系. 生态学报, 2017(07): 2325-2333. |
| [34] |
Parton W, Silver W L, Burke I C, GrassensL, Harmon M E, Currie W S, King J Y, Carol Adair E, Brandt L A, Hart S C, Fasth B. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science, 2007, 315(5810): 361-364. DOI:10.1126/science.1134853 |
| [35] |
李明军, 喻理飞, 杜明凤, 黄宗胜, 石建华. 不同林龄杉木人工林植物-凋落叶-土壤C、N、P化学计量特征及互作关系研究. 生态学报, 2018(21): 1-9. |