 2020, Vol. 40
2020, Vol. 40文章信息
- 王庆, 卢志兴, 赵婧文, 陈又清
- WANG Qing, LU Zhixing, ZHAO Jingwen, CHEN Youqing
- 互利关系和植物多样性对节肢动物群落不同营养级的影响路径和强度
- The impact path and intensity of mutualism and plant diversity on different trophic levels of arthropod community
- 生态学报. 2020, 40(1): 51-59
- Acta Ecologica Sinica. 2020, 40(1): 51-59
- http://dx.doi.org/10.5846/stxb201811062407
-
文章历史
- 收稿日期: 2018-11-06
- 网络出版日期: 2019-10-25

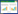




食物网中各营养级之间的相互作用和上行-下行效应在生态系统中具有重要的作用, 是群落动力学的基础[1-5]。在以往的营养级研究中, 研究者们多关注在生产者的上行效应和各营养级之间的捕食与竞争作用对食物网结构的影响, 但是越来越多的研究认为物种间的正相互作用可能与竞争、捕食同等重要[6-9], 而这种正相互作用对食物网中各营养级结构的影响的研究还较为稀少[10-11]。同时, 已有研究极少对这种影响的路径和强度进行量化。
蚂蚁与产蜜露的半翅目昆虫所产生的互利关系是正相互作用的经典模型[12-13], 以蚂蚁-紫胶虫Kerria spp.的互利关系为例, 紫胶虫在寄生过程中会分泌蜜露, 蚂蚁受到蜜露的吸引并前来取食, 蚂蚁在取食蜜露的同时会保护紫胶虫免受天敌的危害[14-15], 两者因此形成了“食物换保护(food-for-protection)”的互利关系[16-17]。蚂蚁不仅是互利关系的参与者, 还是捕食者, 而互利关系有利于强化蚂蚁作为捕食者的作用[18]。同时, 蚂蚁作为生态系统的工程师, 对生物群落结构和生态系统的功能有十分重要的影响[12, 18]。已有研究表明, 蚂蚁与紫胶虫的互利关系会对一些其他节肢动物产生影响, 如蜘蛛、蝗虫类、蝽类等[19-21]。但是这些研究大都只关注于互利关系对某一类群或某一营养级的影响, 它对整个节肢动物群落各营养级相互作用的路径和强度的影响还少有研究。
在陆地生态系统中, 对食物网中不同营养级之间的关系的研究多关注在上行效应, 即植物的多样性和生物量对较高营养级的影响, 大尺度的调查只关注了两个营养层级—生产者和消费者, 而对捕食者以及再往上的顶级捕食者的多度及丰富度的调查研究较少[22-24]。例如, Leles等[25]的研究发现消费者的丰富度会随着植物多样性的升高而增加, 同时捕食者(主要是蚂蚁)的捕食强度也会增强, 生产者的上行效应会传导到第三营养级, 但是对于植物多样性和蚂蚁-产蜜露昆虫的互利关系如何交互影响整个节肢动物群落, 目前还未有研究。
本研究主要探讨互利关系和植物多样性的下行效应和上行效应的对节肢动物群落中不同营养级的相互关系的影响。我们假设:受互利关系影响, 不同强度的捕食作用会产生不同强度的下行效应。同时, 在较小植物多样性变动(单一种植, 两种, 三种混植)的生态系统中, 植物的多样性会对消费者有上行效应, 但是这种上行效应不会传导到更高营养级或者对高营养级影响很弱。通过结构方程模型(SEM), 我们可以量化上行效应和下行效应的路径及其强弱。对于消费者和部分捕食者来说, 如果他们同时受到上行和下行的影响, 结构方程模型的路径分析结果可以说明他们是主要受哪种效应的影响。具体来说, 我们主要关注以下问题:(1)互利关系对其他营养级的下行效应是怎样的?(2)植物多样性-生产力的上行效应是怎样的, 是否会对捕食者阶层有影响?(3)消费者和捕食者是否会同时受到上行效应和下行效应的交互影响, 他们的相对强弱是如何的?
1 研究材料与方法 1.1 研究地概况本研究设置了以蚂蚁-紫胶虫互利关系为关键作用的生态系统模型。研究试验地位于云南省普洱市墨江县雅邑乡(23°14′N, 101°43′E), 海拔为1000—1056 m。该地区年均温17.8℃, 年平均降水量1315.4 mm, 属南亚热带半湿润山地季风气候。紫胶生产是雅邑乡的主要产业之一, 当地常用紫胶寄主植物为牛肋巴Dalbergia obtusifolia、南岭黄檀Dalbergia balansae、思茅黄檀Dalbergia szemaoensis、木豆Cajanus cajan、聚果榕Ficus racemosa等。
1.2 试验设计本研究选取牛肋巴、木豆和南岭黄檀3种树作为紫胶虫的主要寄主植物, 于2015年4月至5月营养袋育苗, 2015年8月将苗木转移到试验地供试。该试验地为3 hm2的钝叶黄檀人工紫胶林, 林地郁闭度为40%左右。参考Moreira等[26]的随机裂区试验设计方法, 本试验裂区的主区为蚂蚁-紫胶虫互利关系因素, 分为3个处理, 第1种为有紫胶虫但是无蚂蚁的无互利关系的处理(Lac insects without ants, LN), 该处理下蚂蚁无法访问、照顾紫胶虫;第2种为有紫胶虫有蚂蚁的有互利关系的处理(Lac insects with ants, LA), 该处理下蚂蚁可以自由访问、照顾紫胶虫;第三种为无紫胶虫但是有蚂蚁的自然对照处理(natural control, NT)。副区为寄主植物多样性因素, 也分为3个处理:第1种处理为3种寄主植物的单一树种, 作为植物多样性1的处理;第2种处理为3种寄主植物的中的任意两种寄主植物混合种植, 作为植物多样性2的处理;第3种处理为3个树种同时出现, 作为植物多样性3的处理。为实现三树种的不同组合, 3种树种组合还引入了当地另一种常用紫胶虫寄主植物聚果榕。苗木生长4个月后, 连同营养袋移入林地中空隙, 每个组合包括6株寄主植物, 分2行种植, 每行3株。相邻的植株之间的距离为15 cm, 组合之间的距离为1.5 m, 以保证组合内相邻的植物之间产生相互作用而组与组之间不会产生相互影响, 各组合在试验林地内随机分布, 每株植物出现在边缘的概率基本一致, 以减少边缘效应对试验结果的影响(图 1中未显示)。按照此方法等分试验区设计4个重复, 每个试验区之间相隔为8—10 m。育苗的营养土、试验地的坡度、坡向、土壤条件、水热等条件均基本一致。于2016年5月放养云南紫胶虫Kerria yunnanensis, 枝条上固定紫胶虫的比例约占枝条总长度的30%。每株植物的茎干上缠绕4 cm宽的透明胶带, 其中控制蚂蚁不能访问的寄主植物上沿胶带涂满粘虫胶, 粘虫胶每2周更换一次, 并清理杂草藤蔓以确保蚂蚁无法到达植株。实验后期收获所有植株并测量每株的生物量。

|
| 图 1 试验样地设计的图解 Fig. 1 Schematic of the experimental design 蚂蚁能否访问紫胶虫的互利关系处理为主区, 3种植物多样性处理为副区, 在试验区不同的树种组合为随机分布(图解中未显示) |
云南紫胶虫一年两代, 其中夏代是最重要的世代, 时间为4月底5月初至9月底10月初。在8—9月份, 是云南紫胶虫泌胶和分泌蜜露重要的时期, 也是节肢动物群落不同营养级物种丰富度和多度最高的时期, 这个时期调查涉及的营养级和类群较多, 比较理想。因此, 寄主植物上节肢动物调查选在云南紫胶虫夏代末期进行。我们于2016年8月和9月分两次调查寄主植物上的节肢动物, 每次调查持续7—10d。在天气晴朗, 植物上没有露水的情况下, 每天从9:00至22:00分3次进行调查。调查人员分为2组, 每组2人, 用手捡法、震落法和网扫法采集每株寄主植物上所有的节肢动物。通过参考相关分类文献并在李巧教授(西南林业大学)的帮助下, 将采集到的节肢动物样本鉴定到种或科, 并确定每个物种的营养级水平。以植物组合为单位, 将两次节肢动物调查的数据合并分析, 并将其划分为紫胶虫(互利关系的参与者, 提供蜜露资源)、蚂蚁(互利关系的参与者, 提供保护的捕食者)、消费者(紫胶虫寄主植物上蚂蚁不照顾的植食性害虫)和除蚂蚁外的捕食者(取食对象为紫胶虫寄主植物上蚂蚁不照顾的植食性害虫, 以下简称为捕食者)等不同的营养级。本试验共采集到蚂蚁28种, 计14767头, 其中, 粗纹举腹蚁Crematogaster macaoensis, 黑头酸臭蚁Tapinoma melanocephalum, 来氏大头蚁Pheidole lighti Wheeler这3种蚂蚁为主要的互利关系参与的种类, 占蚂蚁多度总数的90%以上。消费者共采集到102种1421头, 捕食者共采集到26种264头。每个营养级所有物种的总个体数即为该营养级的多度。
1.4 统计分析我们利用结构方程模型(SEM)来研究受互利关系影响的蚂蚁多度和寄主植物多样性对中层捕食者多度、消费者多度以及植物生物量的直接作用或间接作用, SEM的标准化回归系数可以量化某一个营养级变量对另一个营养级变量的影响程度, 即为营养级之间的路径系数。同一样地类型下不同路径系数之间的大小比较利用参数间差异临界比值来比较, 若该比值的绝对值大于1.96, 则在0.05水平上两系数之间具有显著差异, 不同样地类型下同样的路径系数则用多群组的参数间差异临界比值比较进行比较[27]。在初始模型(图 2)中, 我们假设任何两个营养级的多度之间都会产生相互作用, 包括食物网中的上行效应和下行效应, 这样的模型为饱和模型, 通过删除1条或者n条路径, 去除不显著的变量关系(P> 0.05的路径, 从最大P值开始), 或者基于不同变量之间的残差协方差来增加变量关系, 来修改我们的初始模型[27]。利用χ2(当P> 0.05时说明模型拟合较好), 拟合指数(CFI; 当CFI> 0.95时模型拟合较好)和近似的均方根误差(RMSEA; 当RMSEA < 0.05时模型拟合较好)评估整体模型[28]。植物生物量、消费者多度、捕食者多度和蚂蚁多度数据均经过对数转化, 以符合模型的参数假设。所有的数据分析均在SPSS 22.0和AMOS 22(IBM)软件下进行。

|
| 图 2 上行效应和下行效应的初始模型 Fig. 2 The initial model of the bottom-up effect and top-down effect 矩形框内为各观测变量, 圆形e1-e4为每个观测变量的残差, 下同 |
三种样地食物网的上行-下行效应的路径的结构方程模型如图 3所示。LA样地模型χ2= 4.512, df=6, P=0.608, CFI=1.000, RMSEA < 0.001。LN样地χ2= 5.747, df=6, P=0.452, CFI=1.000, RMSEA=0.002。NT样地χ2= 0.696, df=3, P=0.874, CFI=0.992, RMSEA < 0.001, 3个模型的拟合均良好, 能较好地表示出各营养级之间的路径系数。

|
| 图 3 互利关系与植物多样性对节肢动物群落营养级的上行—下行效应影响的路径分析 Fig. 3 Path analysis of bottom-up and top-down effects of mutualism relationship and plant diversity on the trophic level of arthropod community LA:有互利关系处理样地Lac insects without ants; LN:无互利关系处理样地Lac insects with ants; NT:自然对照样地Natural control; 图中不显著的路径为虚线, 显著的路径为实线, 箭头上的数字为标准化路径系数β, 变量的左上角为决定系数R2值 |
从表 1中可以看出, 在LA样地, 蚂蚁多度对捕食者多度和消费者多度均有显著影响, 其标准化路径系数β分别为-0.93(P=0.036)和-0.33(P=0.01), 捕食者多度对消费者多度也有显著影响(β=-0.57, P < 0.001)。在LN样地, 捕食者多度对消费者没有显著影响(β=-0.33, P=0.173)。在NT样地, 蚂蚁多度对捕食者多度(β=-0.36, P=0.024)和消费者多度(β=-0.36, P=0.005)均有显著影响, 而捕食者多度对消费者多度也有显著影响(β=-0.48, P < 0.001)。
| 上行-下行效应路径 Path of bottom-up and top-down effect |
LA | LN | NT | ||||||||
| β | C.R. | P | β | C.R. | P | β | C.R. | P | |||
| 植物生物量←植物多样性 Plant biomass←Plant diversity |
0.46 | 3.157 | 0.002 | 0.51 | 8.652 | 0.039 | 0.60 | 4.114 | 0.027 | ||
| 消费者多度←植物生物量 Consumer abundance←Plant biomass |
0.638 | 4.903 | < 0.001 | 0.745 | 5.923 | < 0.001 | 0.701 | 5.461 | < 0.001 | ||
| 捕食者多度←植物生物量 Predator abundance←Plant biomass |
0.423 | 3.476 | < 0.001 | -0.332 | -0.814 | 0.416 | 0.271 | 0.319 | 0.750 | ||
| 蚂蚁多度←植物生物量 Ant abundance←Plant biomass |
0.201 | 1.218 | 0.223 | 0.196 | 1.103 | 0.270 | |||||
| 捕食者多度←消费者多度 Predator abundance← Consumer abundance |
0.496 | 4.076 | < 0.001 | 0.616 | 4.622 | < 0.001 | 0.864 | 3.045 | < 0.001 | ||
| 蚂蚁多度←捕食者多度 Ant abundance←Predator abundance |
0.091 | 0.422 | 0.656 | 0.112 | 0.162 | 0.545 | |||||
| 蚂蚁多度←消费者多度 Ant abundance← Consumer abundance |
0.267 | 1.348 | 0.178 | 0.261 | 1.127 | 0.321 | |||||
| 捕食者多度←蚂蚁多度 Predator abundance← Ant abundance |
-0.928 | -2.099 | 0.036 | -0.357 | -2.264 | 0.024 | |||||
| 消费者多度←捕食者多度 Consumer abundance← Predator abundance |
-0.566 | -4.628 | < 0.001 | -0.331 | -1.363 | 0.173 | -0.487 | -3.775 | < 0.001 | ||
| 消费者多度←蚂蚁多度 Consumer abundance← Ant abundance |
-0.329 | -2.560 | 0.01 | -0.359 | -2.785 | 0.005 | |||||
| C.R.为临界比值, 若该值的绝对值大于1.96说明在0.05水平上两路径系数大小具有显著差异; LA:有互利关系处理样地Lac insects without ants; LN:无互利关系处理样地Lac insects with ants; NT:自然对照样地Natural control | |||||||||||
表 2显示了三种样地下行效应的显著路径系数的两两对比的参数差异临界比值。在LA样地中, 捕食者多度←蚂蚁多度路径系数显著大于消费者多度←捕食者多度路径(C.R.=4.621), 消费者多度←蚂蚁多度路径系数也显著大于消费者多度←捕食者多度(C.R.=4.583), 消费者多度←蚂蚁多度路径和捕食者多度←蚂蚁多度路径系数无显著差异(C.R.=1.72)。在NT样地, 捕食者多度←蚂蚁多度的路径系数显著大于消费者多度←捕食者多度路径(C.R.=3.737), 消费者多度←蚂蚁多度路径系数显著大于消费者多度←捕食者多度路径(C.R.=3.474)和捕食者多度←蚂蚁多度路径(C.R.=2.205)。
| 样地类型 Plot type |
对比路径 Contrast path |
参数差异临界比值 C.R. |
|
| LA | 消费者多度←捕食者多度 | 捕食者多度←蚂蚁多度 | -4.621 |
| LA | 消费者多度←捕食者多度 | 消费者多度←蚂蚁多度 | -4.583 |
| LA | 捕食者多度←蚂蚁多度 | 消费者多度←蚂蚁多度 | 1.72 |
| NT | 消费者多度←捕食者多度 | 捕食者多度←蚂蚁多度 | -3.737 |
| NT | 消费者多度←捕食者多度 | 消费者多度←蚂蚁多度 | -3.474 |
| NT | 捕食者多度←蚂蚁多度 | 消费者多度←蚂蚁多度 | 2.205 |
从表 1中可以看出, 3种样地植物多样性对植物生物量均有显著影响, 其标准化路径系数β分别为:LA样地β=0.46, P=0.02;LN样地β=0.60, P=0.039;NT样地β=0.51, P=0.027。受植物多样性影响的植物生物量在3种样地的上行效应路径具有差异:在LA样地, 植物生物量对消费者多度和捕食者多度均有显著影响(β=0.64, P < 0.001; β=0.42, P < 0.001), 但是植物生物量对蚂蚁多度没有显著影响(β=0.201, P=0.223);在LN样地, 植物生物量只对消费者多度有显著影响(β=0.71, P < 0.001), 而对捕食者多度没有显著影响(β=-0.33, P=0.416);在NT样地, 植物生物量对只对消费者多度有显著影响(β=0.75, P < 0.001), 对捕食者多度和蚂蚁多度均没有显著影响(β=0.27, P=0.210; β=0.20, P=0.750)。
3种样地消费者多度对捕食者多度均有显著影响, 其标准化路径系数β分别为:LA样地β=0.50, P < 0.001;LN样地β=0.62, P < 0.001;NT样地β=0.86, P < 0.001。而消费者多度、捕食者多度对蚂蚁多度均没有显著影响。
表 3显示了3种样地上行效应的显著路径系数的两两对比的参数差异临界比值。LA样地中任意两条路径的参数差异临界比值均大于1.96, 即任意两条路径系数的大小具有显著差异。在LN样地, 植物多样性—消费者多度和消费者多度—捕食者多度这两条路径的C.R.=2.880>1.96, 两条路径之间有显著差异;在NT样地, 这两条路径的C.R.=1.110 < 1.96, 没有显著差异。
| 样地类型 Plot type |
对比路径 Contrast path |
参数差异临界比值 C.R. |
|
| LA | 消费者多度←植物生物量 | 捕食者多度←消费者多度 | 3.319 |
| LA | 消费者多度←植物生物量 | 捕食者多度←植物生物量 | -4.301 |
| LA | 捕食者多度←消费者多度 | 捕食者多度←植物生物量 | -3.905 |
| LN | 消费者多度←植物生物量 | 捕食者多度←消费者多度 | 2.88 |
| NT | 消费者多度←植物生物量 | 捕食者多度←消费者多度 | 1.11 |
通过多群组比较(表 4), 植物生物量←植物多样性路径、消费者多度←植物生物量和捕食者多度←消费者多度这3条上行路径在3种样地之间的两两比较结果显示, 这几条路径系数在3种样地之间均没有显著差异。在下行路径中, 只有捕食者多度←蚂蚁多度这条路径LA样地要显著高于NT样地(C.R.=2.236), 其他的路径系数之间没有显著差异。
| 对比路径 Contrast path |
多群组比较Multi-group comparison | ||
| LA-LN | LA-NT | LN-NT | |
| 植物生物量←植物多样性 Plant biomass←Plant diversity |
0.335 | 0.247 | 0.142 |
| 消费者多度←植物生物量 Consumer abundance←Plant biomass |
1.045 | 0.826 | 0.818 |
| 捕食者多度←消费者多度 Consumer abundance← Predator abundance |
0.357 | 0.596 | 0.378 |
| 捕食者多度←蚂蚁多度 Predator abundance← Ant abundance |
2.236 | ||
| 消费者多度←蚂蚁多度 Consumer abundance← Ant abundance |
0.454 | ||
| 消费者多度←捕食者多度 Consumer abundance← Predator abundance |
0.176 | 0.143 | 1.201 |
在各种生态系统中, 食物网的上行效应与下行效应的相对强弱一直是生态学家争论的主题之一[1, 29-30]。许多蚂蚁是杂食性种类, 但是它们在节肢动物食物网中起了重要的捕食作用, 它们有活跃的捕食习性、广泛的捕食对象、高效的募集能力和庞大的群体数量, 是食物网的重要组成部分。因此, 我们将参与互利关系的杂食性蚂蚁作为一类捕食者进行分析, 同时, 蚂蚁与产蜜露昆虫所形成的互利关系可能会增强蚂蚁作为捕食者对节肢动物群落的影响[1, 31-32]。在本研究中, 试验所设置的样地类型为人工紫胶林生态系统, 该生态系统中蚂蚁类群由于和紫胶虫形成了互利关系, 蚂蚁的群体数量大为提升, 蚂蚁类群的多度为14767, 其中90%与紫胶虫形成了互利关系, 而包括鞘翅目、鳞翅目等其他捕食者和消费者的多度分别为264和1421, 因此, 蚂蚁在人工紫胶林生态系统占据了优势地位, 它们会作为捕食者对不同的营养级产生影响[33-34]。我们的结果显示, 在有互利关系(LA)样地, 蚂蚁对捕食者的下行效应显著强于蚂蚁对消费者的下行效应。而在自然对照样地(NT), 蚂蚁对捕食者的下行效应和对消费者的下行效应没有显著差异。不同样地间的多群组比较显示, 有互利关系的蚂蚁对捕食者的下行效应要显著强于自然对照(NT)样地, 但是蚂蚁对消费者的下行效应在两种处理下没有显著差异。虽然蚂蚁会同时攻击或捕食食物网中其他的捕食者和消费者, 但是在不同样地间蚂蚁只会对捕食者的下行效应有显著差异, 说明互利关系增强了蚂蚁对捕食者的下行效应。
已有许多研究表明, 植物的多样性会对植物的生物量(生产力)有正效应[35-37]。在之前的研究中, 我们也发现植物的物种多样性会提高其生产力[38]。在本研究中, 所有类型的样地中的植物多样性对植物的生物量均有显著的正相关, 而植物生物量也均对消费者多度有正效应, 说明植物多样性会通过增加植物生物量而对消费者阶层产生上行效应。寄主植物的生物量不仅对消费者阶层有上行效应, 同样也对捕食者阶层有间接的影响, 而消费者也会对蚂蚁产生一定的影响。植物生物量对消费者多度、捕食者多度以及消费者多度对捕食者多度的上行路径系数分别为0.64, 0.50和0.42, 其路径系数随着营养级的升高而逐渐减少。另一方面, 只有LA样地的植物多样性对捕食者有上行效应, 而LN样地和NT样地植物多样性的上行效应均不会传导到捕食者阶层。也就是说, 生产者的上行效应随着营养级的升高而减弱, 这与前人提出的营养衰减假说(Nutrient Attenuation Hypothesis)相一致[39]。但是互利关系会强化这种传递强度, 使其达到捕食者营养级。
综合互利关系的下行效应和植物多样性的上行效应对消费者和捕食者多度的影响效果, 我们的结果显示:蚂蚁-紫胶虫互利关系对食物网中的捕食者和消费者都有显著的下行效应, 同时, 植物多样性对消费者有显著的上行效应, 这种效应在3种处理下的路径系数都显著高于蚂蚁-紫胶虫互利关系的下行效应。因此, 对于消费者来说, 植物多样性的上行效应影响要强于互利关系的下行效应。对捕食者来说, 蚂蚁-紫胶虫互利关系对捕食者的影响要显著高于自然对照, 而植物多样性对捕食者的上行效应路径不显著, 说明其主要受到来自互利关系的下行效应影响。但是总的来说, 在有互利关系的存在时, 食物网中各营养级之间的路径更多, 相互关系更为复杂, 这对于群落生态系统的稳定性是有利的。
结构方程模型(SEM)可以同时将各种变量作为自变量和因变量, 并且SEM分析方法可以量化他们的相互影响的重要值, 进而进行比较[40]。已有许多研究利用结构方程模型来研究水生生态系统中食物网的营养级联, 但是该模型在陆地生态系统的应用还较少。本研究利用SEM分析研究了以蚂蚁-紫胶虫互利关系为核心作用的紫胶林生态系统中, 互利关系和植物多样性对节肢动物食物网中各营养级的影响, 揭示了上行效应和下行效应对节肢动物各营养级的影响路径和强度。本研究的样本量(n=108)对于食物网中的多营养级研究来说是足够的, 但对于使用结构方程模型来说是偏小的[41], 因此, 我们的结果可能不能被认定为确定性的, 但能作为有益的探索。
本研究范围是小尺度空间的节肢动物多营养级之间相互关系, 事实上生物之间相互作用的影响是极为广泛的, 比如对植物生理性状、对土壤营养状况等等[42-43]。在未来的研究中, 应兼顾研究的空间、时间尺度与营养级的复杂程度, 以期能在更大尺度、更多营养级数上来探讨食物网中上行-下行效应的影响。
| [1] |
Kay C E. Are ecosystems structured from the top-down or bottom-up:a new look at an old debate. Wildlife Society Bulletin (1973-2006), 1998, 26(3): 484-498. |
| [2] |
Duffy J E. Biodiversity loss, trophic skew and ecosystem functioning. Ecology Letters, 2003, 6(8): 680-687. DOI:10.1046/j.1461-0248.2003.00494.x |
| [3] |
Grange S, Duncan P. Bottom-up and top-down processes in African ungulate communities:resources and predation acting on the relative abundance of zebra and grazing bovids. Ecography, 2006, 29(6): 899-907. DOI:10.1111/j.2006.0906-7590.04684.x |
| [4] |
Srivastava D S. Habitat structure, trophic structure and ecosystem function:interactive effects in a bromeliad-insect community. Oecologia, 2006, 149(3): 493-504. DOI:10.1007/s00442-006-0467-3 |
| [5] |
Banse K. Do we live in a largely top-down regulated world?. Journal of Biosciences, 2007, 32(4): 791-796. DOI:10.1007/s12038-007-0080-6 |
| [6] |
Bronstein J L. Our current understanding of mutualism. Quarterly Review of Biology, 1994, 69(1): 31-51. DOI:10.1086/418432 |
| [7] |
Stachowicz J J. Mutualism, facilitation, and the structure of ecological communities:positive interactions play a critical, but underappreciated, role in ecological communities by reducing physical or biotic stresses in existing habitats and by creating new habitats on which many species depend. BioScience, 2001, 51(3): 235-246. DOI:10.1641/0006-3568(2001)051[0235:MFATSO]2.0.CO;2 |
| [8] |
Bruno J F, Stachowicz J J, Bertness M D. Inclusion of facilitation into ecological theory. Trends in Ecology & Evolution, 2003, 18(3): 119-125. |
| [9] |
Crowley P H, Cox J J. Intraguild mutualism. Trends in Ecology & Evolution, 2011, 26(12): 627-633. |
| [10] |
Kaplan I, Eubanks M D. Aphids alter the community-wide impact of fire ants. Ecology, 2005, 86(6): 1640-1649. DOI:10.1890/04-0016 |
| [11] |
Wang R W, Shi L, Ai S M, Zheng Q. Trade-off between reciprocal mutualists:local resource availability-oriented interaction in fig/fig wasp mutualism. Journal of Animal Ecology, 2008, 77(3): 616-623. DOI:10.1111/j.1365-2656.2008.01359.x |
| [12] |
Stadler B, Dixon A F G. Ecology and evolution of aphid-ant interactions. Annual Review of Ecology, Evolution, and Systematics, 2005, 36: 345-372. DOI:10.1146/annurev.ecolsys.36.091704.175531 |
| [13] |
Ness J H, Mooney K, Lach L. Ants as mutualists//Lach L, Parr C, Abbott K, eds. Ant Ecology. Oxford: Oxford University, 2010: 97-114.
|
| [14] |
Heil M, McKey D. Protective ant-plant interactions as model systems in ecological and evolutionary research. Annual Review of Ecology, Evolution, and Systematics, 2003, 34: 425-453. DOI:10.1146/annurev.ecolsys.34.011802.132410 |
| [15] |
Chen Y Q, Wang S M, Lu Z X, Lu Z X, Liu C J, Zhang W. The effects of ant attendance on aggregation of the honeydew producing lac insect Kerria yunnanensis. Tropical Ecology, 2013, 54(3): 301-308. |
| [16] |
Way M J. Mutualism between ants and honeydew-producing homoptera. Annual Review of Entomology, 1963, 8: 307-344. DOI:10.1146/annurev.en.08.010163.001515 |
| [17] |
Chen Y Q, Lu Z X, Li Q, Hoffmann B D, Zhang W. Multiple ant species tending lac insect Kerria yunnanensis (Hemiptera:Kerriidae) provide asymmetric protection against parasitoids. PLoS One, 2014, 9(6): e98975. DOI:10.1371/journal.pone.0098975 |
| [18] |
Styrsky J D, Eubanks M D. Ecological consequences of interactions between ants and honeydew-producing insects. Proceedings of the Royal Society B:Biological Sciences, 2007, 274(1607): 151-164. DOI:10.1098/rspb.2006.3701 |
| [19] |
陈彦林, 陈又清, 李巧, 章彦, 周兴银. 紫胶虫生境蜘蛛群落的初步研究. 福建林学院学报, 2008, 28(2): 179-183. DOI:10.3969/j.issn.1001-389X.2008.02.018 |
| [20] |
陈又清, 李巧, 王思铭. 紫胶林-农田复合生态系统的蚱总科昆虫多样性. 林业科学, 2011, 47(9): 100-107. |
| [21] |
李巧, 陈又清, 陈彦林. 紫胶林-农田复合生态系统蝽类昆虫群落多样性. 云南大学学报:自然科学版, 2009, 31(2): 208-216. |
| [22] |
卢志兴, 陈又清, 张威, 王思铭, 李巧. 蚂蚁-紫胶虫兼性互利关系对蚂蚁群落多样性的影响. 生物多样性, 2013, 21(3): 343-351. |
| [23] |
Petermann J S, Müller C B, Weigelt A, Weisser W W, Schmid B. Effect of plant species loss on aphid-parasitoid communities. Journal of Animal Ecology, 2010, 79(3): 709-720. DOI:10.1111/j.1365-2656.2010.01674.x |
| [24] |
李妍, 李国勇, 慕军鹏, 孙书存. 消费者多样性对食物网结构和生态系统功能的影响. 生态学报, 2008, 28(1): 388-398. DOI:10.3321/j.issn:1000-0933.2008.01.045 |
| [25] |
Leles B, Xiao X, Pasion B O, Nakamura A, Tomlinson K W. Does plant diversity increase top-down control of herbivorous insects in tropical forest?. Oikos, 2017, 126(8): 1142-1149. DOI:10.1111/oik.03562 |
| [26] |
Moreira X, Mooney K A, Zas R, Sampedro L. Bottom-up effects of host-plant species diversity and top-down effects of ants interactively increase plant performance. Proceedings of the Royal Society B:Biological Sciences, 2012, 279(1746): 4464-4472. DOI:10.1098/rspb.2012.0893 |
| [27] |
吴明隆. 结构方程模型-AMOS的操作与应用(第二版). 重庆:重庆大学出版社, 2010, 412-417. |
| [28] |
Grace J B. Structural Equation Modeling and Natural Systems. Cambridge, UK:Cambridge University Press, 2006, 115-142. |
| [29] |
Shoesmith E. Structural equation modeling with AMOS:basic concepts, applications, and programming by B. M. Byrne. Journal of the Royal Statistical Society. Series D (The Statistician), 2002, 51(3): 408-409. |
| [30] |
Sinclair A R E, Krebs C J. Complex numerical responses to top-down and bottom-up processes in vertebrate populations. Philosophical Transactions of the Royal Society of Society B:Biological Sciences, 2002, 357(1425): 1221-1231. DOI:10.1098/rstb.2002.1123 |
| [31] |
Dyer LA, Letourneau D. Top-down and bottom-up diversity cascades in detrital vs. living food webs. Ecology Letters, 2003, 6(1): 60-68. |
| [32] |
Hölldobler B, Wilson E O. The Ants. Cambridge, Mass.: Belknap Press of Harvard University Press, 1990.
|
| [33] |
Mooney K A. The disruption of an ant-aphid mutualism increases the effects of birds on pine herbivores. Ecology, 2006, 87(7): 1805-1815. DOI:10.1890/0012-9658(2006)87[1805:TDOAAM]2.0.CO;2 |
| [34] |
张霜, 张育新, 马克明. 保护性的蚂蚁-植物相互作用及其调节机制研究综述. 植物生态学报, 2010, 34(11): 1344-1353. DOI:10.3773/j.issn.1005-264x.2010.11.012 |
| [35] |
Tilman D, Lehman C L, Thomson K T. Plant diversity and ecosystem productivity:theoretical considerations. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(5): 1857-1861. DOI:10.1073/pnas.94.5.1857 |
| [36] |
Grace J B, Anderson T M, Seabloom E W, Borer E T, Adler P B, Harpole W S, Hautier Y, Hillebrand H, Lind E M, Pärtel M, Bakker J D, Buckley Y M, Crawley M J, Damschen E I, Davies K F, Fay P A, Firn J, Gruner D S, Hector A, Knops J M H, MacDougall A S, Melbourne B A, Morgan J W, Orrock J L, Prober S M, Smith M D. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature, 2016, 529(7586): 390-393. DOI:10.1038/nature16524 |
| [37] |
江小雷, 张卫国. 一年生植物群落内取样效应和互补效应的分离. 生态学报, 2006, 26(6): 1896-1902. DOI:10.3321/j.issn:1000-0933.2006.06.034 |
| [38] |
王庆, 卢志兴, 赵婧文, 陈又清. 物种丰富度及物种间相互作用对植物生长的影响——以木本紫胶虫寄主植物苗木为例. 云南大学学报:自然科学版, 2018, 40(2): 398-404. |
| [39] |
邱东茹, 吴振斌. 生物操纵、营养级联反应和下行影响. 生态学杂志, 1998, 17(5): 27-32. DOI:10.3321/j.issn:1000-4890.1998.05.007 |
| [40] |
Ruehl C B, Trexler J C. A suite of prey traits determine predator and nutrient enrichment effects in a tri-trophic food chain. Ecosphere, 2013, 4(6): 444-454. |
| [41] |
Kline R B. Principles and practice of structural equation modeling, third edition. New York: Guilford, 2011: 46-73.
|
| [42] |
Holland J N, Chamberlain S A, Horn K C. Optimal defence theory predicts investment in extrafloral nectar resources in an ant-plant mutualism. Journal of Ecology, 2009, 97(1): 89-96. DOI:10.1111/j.1365-2745.2008.01446.x |
| [43] |
Sampedro L, Moreira X, Martíns P, Zas R. Growth and nutritional response of Pinus pinaster after a large pine weevil (Hylobius abietis) attack. Trees, 2009, 23(6): 1189-1197. DOI:10.1007/s00468-009-0358-4 |