 2019, Vol. 39
2019, Vol. 39文章信息
- 沈园, 李涛, 唐明方, 邓红兵
- SHEN Yuan, LI Tao, TANG Mingfang, DENG Hongbing
- 西南地区生物文化多样性空间格局定量研究
- Spatial patterns of biocultural diversity in Southwest China
- 生态学报. 2019, 39(7): 2454-2461
- Acta Ecologica Sinica. 2019, 39(7): 2454-2461
- http://dx.doi.org/10.5846/stxb201804060776
-
文章历史
- 收稿日期: 2018-04-06
- 网络出版日期: 2019-01-10





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
人类社会存续与发展的历程, 既是人类开发并利用自然的过程, 也是人类创造和发展文化的过程。在此过程中, 生物多样性是人类社会赖以生存和发展的基础[1];文化多样性是人类社会的基本特征, 也是人类文明进步的重要动力[2]。虽然生物多样性是自然系统的属性, 而文化多样性反映的是社会系统的特征, 但两者都是保持自然界和人类社会健康的基础, 两者之间也存在密不可分的联系[3]。文化多样性和生物多样性的联系常反映在地理空间上的交叉重合。在早期的生物多样性研究中, 就有学者观察到生物多样性丰富的地区相应地拥有丰富的传统文化[4]。因高山、河流等复杂的地理要素所引起的地理隔离, 不仅推动了生物类群的演化, 也促进了不同群体、民族的分化[5]。生物文化多样性是指生物多样性和文化多样性之间的复杂连结, 可被认为是世界差异的总和[3, 6]。生物文化多样性作为完整的概念用于研究与管理的时间不长[2], 但越来越多的学者强调了生物多样性与文化多样性连结的重要性[7], 并形成了生物文化视角及应用于管理实践的生物文化途径[8]。
西南地区幅员辽阔、地貌复杂、气候多样、水汽丰沛, 为动植物的演化和发展创造了有利条件。因而, 西南地区拥有垂直地带性较为完整, 类型多样、种类齐全、物种丰富的生态系统类型, 成为我国重要的生物资源宝库[9]。与此同时, 西南地区是我国少数民族的聚居区, 世居的34个民族人口占全国少数民族总人口的48.32%[10]。由于分布区的地理环境、气候条件、生产资料等因素各异, 不同民族在历史的演化过程中形成了各具特色的传统文化[11]。西南地区独特的生物资源条件与民族文化形式对多样性的空间格局与影响因素研究具有重要的价值, 但当前仍然缺乏对该区域生物文化多样性的定量研究以及较大尺度上的空间格局分析。因此, 在分别探讨文化多样性与生物多样性的分布格局的基础上, 综合生物多样性和文化多样性指标来计算生物文化多样性, 从而识别西南地区生物文化多样性空间格局, 并分析不同自然因素对多样性分布格局的影响, 研究结果有助于从综合的视角来了解和保护区域生物文化多样性。
1 数据与方法 1.1 数据来源本研究中, 西南地区横跨85°30′—112°30′E, 20°54′—36°29′N, 包括广西壮族自治区、重庆市、四川省、贵州省和云南省的全部, 以及西藏自治区的拉萨市、那曲地区、山南地区、林芝地区、昌都地区与青海省的玉树藏族自治州(含格尔木市飞地唐古拉乡)。文化多样性数据是通过借用Shannon-Wiener指数对2010年第六次人口普查中各民族人口数进行计算所得[12-14]。生物多样性数据来源于《2010年中国生态系统服务空间数据集》通过收集整理区域内的指示物种数量得到生物多样性保护重要性模拟数据[15]。行政区划与边界矢量、水体矢量数据以及数字高程模型来源于中国科学院遥感与数字地球研究所。研究中以县级行政区为基本单元, 2010年的研究单元总数为603个。
1.2 生物文化多样性在较大尺度的研究中, 生物文化多样性指标的选取或构建是重要的前提, 常用的简便方法是将定量生物多样性和文化多样性的指标加以综合用来表征整体的多样性。本研究中, 在咨询专家的意见并参考国际上相关研究的基础上[6], 认为文化多样性与生物多样性在西南地区具有同等重要性, 对两者赋等权求和得到生物文化多样性。第i个研究单元的生物文化多样性BCDi, 具体表示为:
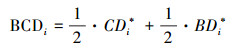
|
(1) |
式中, CDi*与BDi*分别为第i个研究单元的文化多样性与生物多样性经无量纲化处理后的值。
为使文化多样性与生物多样性具有可比性, 并消除两种多样性各单元变量在变异程度上的差异, 本研究采用标准差化无量纲处理方法。第i个研究单元经处理后的多样性Di*, 具体表示为:
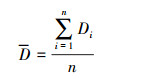
|
(2) |
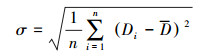
|
(3) |
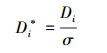
|
(4) |
式中, Di为第i个研究单元的多样性数值, D为全部研究单元的平均值, σ为全部研究单元的标准差, n为研究单元总数。
1.3 优化的热点分析当空间数据可视化后, 目标单元不仅具有高值, 而且被其他同样具有高值的单元所包围, 才能成为具有显著统计学意义的热点。优化的热点分析能够自动聚合事件数据、识别适当的分析范围, 并通过对数据进行查询获得产生最佳热点结果的设置[16]。通过汇总空间现象聚类的程度, 不同区域的聚类水平得以比较, 同时其高值或低值在空间上发生聚集的位置也得以识别。在Getis-Ord G*统计工具的运行过程中, 会对比目标单元及其相邻单元的局部总和与所有单元的总和;当局部总和与预期值有很大差异, 不能取得随机产生的结果时, 就会产生一个具有显著统计学意义的得分Gi*, 具体表示为[17]:

|
(5) |
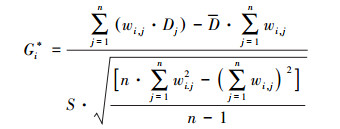
|
(6) |
式中, Dj为第j个研究单元的多样性数值, wi, j为单元i与单元j间的空间权重。
2 研究结果 2.1 文化多样性与生物多样性空间格局对比西南地区的文化多样性与生物多样性空间分布如图 1所示。一方面, 西南地区文化多样性高的区域为澜沧江流域、怒江流域和雅鲁藏布江流域的下游区, 元江流域全境, 以及珠江流域上游区;而文化多样性低的区域为长江流域的四川盆地大部, 澜沧江流域、怒江流域和雅鲁藏布江流域的上游区, 以及广西沿海地区。另一方面, 生物多样性高的区域主要在青藏高原与四川盆地的过渡带至三江并流的横断山地区, 以及澜沧江流域与元江流域下游的边境处;而生物多样性低的区域为各流域的上游区, 珠江流域全境, 以及重庆全境。凭借ArcGIS 10.2的多种工具[18], 提取河流干流处县级单元的文化多样性与生物多样性数值, 并沿河流流向对比多样性的变化情况。在文化多样性方面, 澜沧江干流、怒江干流、雅鲁藏布江干流与元江干流沿河流流向大体上都呈现“低-高”的变化状态, 而长江干流与珠江干流都呈现“低-高-低”的变化状态。在生物多样性方面, 雅鲁藏布江干流与元江干流沿河流流向都呈现“低-高”的变化状态, 而长江干流、珠江干流与怒江干流都呈现“低-高-低”的变化状态, 此外, 澜沧江干流呈现出“低-高-低-高”的变化状态。对比文化多样性与生物多样性的分布格局发现, 两者在西南地区存在一定的地理空间分布差异。

|
| 图 1 西南地区文化多样性与生物多样性空间分布 Fig. 1 The spatial distribution of cultural and biological diversity of Southwest China |
运用优化的热点分析工具[18], 对西南地区文化多样性与生物多样性进行空间聚类分析发现, 两者的高低值聚类区空间分布具有一定差异, 但也存在高低值聚类的重叠区域。从图 2可以看出, 文化多样性在西南地区南部形成明显的条带状高值聚类区;同时, 在雅鲁藏布江流域、怒江流域和澜沧江流域上游区, 与长江流域内的四川盆地, 以及广西沿海地区形成低值聚类区。生物多样性在青藏高原与四川盆地过渡带至横断山区形成明显的条带状高值聚类区;同时, 在雅鲁藏布江流域、怒江流域和长江流域上游, 与重庆周围形成低值聚类区。对比文化多样性与生物多样性的空间聚类格局发现, 横断山区, 与澜沧江流域和元江流域下游为文化多样性与生物多样的高值聚类区;而雅鲁藏布江流域和怒江流域的上游区, 与以重庆为中心的四川盆地平原区, 为两种多样性的低值聚类区。此外, 贵州南部至广西北部为文化多样性高值聚类而非生物多样性高值聚类区, 青藏高原东缘至四川盆地交界处为生物多样性高值聚类而非文化多样性高值聚类区。

|
| 图 2 西南地区文化多样性与生物多样性高低聚类空间分布 Fig. 2 The spatial distribution of high/low clustering of cultural and biological diversity of Southwest China |
基于西南地区的文化多样性与生物多样性县级单元数据, 计算得到各单元的生物文化多样性(图 3)。西南地区生物文化多样性高的区域为青藏高原与四川盆地的过渡带至三江并流的横断山地区, 以及澜沧江流域与元江流域下游的边境处;而生物文化多样性低的区域为长江流域、澜沧江流域、怒江流域和雅鲁藏布江流域的上游区, 以及四川盆地大部和广西沿海地区。通过提取河流干流处县级单元的生物文化多样性并沿河流流向对比发现, 雅鲁藏布江干流与元江干流沿河流流向都呈现“低-高”的变化状态, 而长江干流、珠江干流与怒江干流都呈现“低-高-低”的变化状态, 此外, 澜沧江干流呈现出“低-高-低-高”的变化状态。将生物文化多样性的空间格局与文化多样性和生物多样性的相比较发现, 高生物文化多样性的区域与高生物多样性的区域分布较为相似, 而低生物文化多样性的区域与低文化多样性的区域分布较为接近。

|
| 图 3 西南地区生物文化多样性及其高低聚类空间分布 Fig. 3 The spatial distribution of biocultural diversity and its high/low clustering of Southwest China |
运用优化的热点分析工具, 对西南地区生物文化多样性进行空间聚类分析发现, 其高低值聚类区呈现明显的片状格局。从图 3可以看出, 生物文化多样性自青藏高原东缘至广西与贵州西部形成明显的片状高值聚类区;同时, 在长江流域内的四川盆地, 以及雅鲁藏布江流域、怒江流域、澜沧江流域和长江流域的上游, 分别形成明显的片状低值聚类区。与文化多样性和生物多样性的高低值聚类分布格局相比较发现, 生物文化多样性高低值聚类分布区的片状格局更为明显, 这主要是由文化多样性与生物多样性的高低值聚类的重叠效应所引起的。
2.3 影响因素分析本研究通过对西南地区的区域面积、水体面积、水体周长、水体覆盖率、平均海拔、海拔标准差、平均坡度与坡度标准差进行分析, 探究自然因素对文化多样性、生物多样性与生物文化多样性空间格局所产生的影响。运用ArcGIS 10.2的Exploratory Regression工具得到各因素的变量显著性[18-19], 反映不同影响因素对多样性指数的预测程度。此外, 应用IBM SPSS Statistics 20统计软件将所有研究单元的多样性数值与因素值进行Pearson相关分析[20], 探究文化多样性、生物多样性、生物文化多样性与影响因素之间的线性相关关系。
各影响因素对文化多样性空间分布的研究发现(表 1), 平均坡度的变量显著性为100%, 体现出了研究单元内地形坡度的平均值对文化多样性的空间分布具有很强的影响。同时, 水体覆盖率和海拔标准差对文化多样性的变量显著性也较高, 反映出研究单元内水体面积占区域面积的比值和海拔高程的差异程度对文化多样性空间分布产生一定影响[12]。相反地, 区域面积与水体面积的影响则较弱。Pearson相关分析的结果表明, 文化多样性与平均坡度、坡度标准差和海拔标准差呈极显著(P<0.01)的正相关关系, 而与水体覆盖率呈极显著(P<0.01)的负相关关系。同时, 文化多样性与水体周长呈显著(P<0.05)的负相关, 但相关性较低。综上所述, 西南地区文化多样性分布格局受到平均坡度([1.37°, 32.69°])的重要影响, 文化多样性水平高的单元倾向于聚集在地形坡度大、海拔与坡度的差异程度较高而水体覆盖率较低的区域。
| 多样性 Diversity |
统计值 Statistics |
区域面积 Area |
水体面积 Water area |
水体周长 Water perimeter |
水体覆盖率 Water coverage |
平均海拔 Average altitude |
海拔标准差 Altitude SD |
平均坡度 Average slope |
坡度标准差 Slope SD |
| 文化Cultural | 变量显著性/% | 23.23 | 27.27 | 40.40 | 74.75 | 54.55 | 73.74 | 100 | 60.61 |
| Pearson相关系数 | -0.0274 | -0.0787 | -0.0889* | -0.1890** | 0.0461 | 0.1845** | 0.3204** | 0.2621** | |
| 生物Biological | 变量显著性/% | 17.17 | 60.61 | 2.02 | 11.11 | 64.65 | 100 | 79.80 | 36.36 |
| Pearson相关系数 | 0.0134 | -0.0739 | -0.0300 | -0.1216** | 0.1489** | 0.3940** | 0.3596** | 0.2694** | |
| 生物文化Biocultural | 变量显著性/% | 25.25 | 39.39 | 18.18 | 54.55 | 66.67 | 57.58 | 100 | 57.58 |
| Pearson相关系数 | -0.0082 | -0.0896 | -0.0698 | -0.1824** | 0.1145** | 0.3397** | 0.3994** | 0.3121** | |
| *表示在0.05水平上(双侧)显著相关, **表示在0.01水平上(双侧)显著相关 | |||||||||
各影响因素对生物多样性空间分布的研究发现(表 1), 海拔标准差的变量显著性达100%, 其次为平均坡度, 而水体周长的变量显著性最低仅有2.02%, 体现出研究单元内海拔高程的差异程度和地形坡度的平均值对生物多样性的空间分布具有很强的影响, 但水体的周界长度对生物多样性的空间分布影响很弱。与此同时, Pearson相关分析的结果表明, 生物多样性与海拔标准差、平均坡度、坡度标准差和平均海拔呈极显著(P<0.01)的正相关关系, 而与水体覆盖率呈极显著(P<0.01)的负相关关系。因此, 西南地区生物多样性分布格局受到海拔标准差([5.34 m, 1619.60 m])与平均坡度([1.37°, 32.69°])的影响, 生物多样性高的单元倾向于聚集在地形坡度大且海拔与坡度的差异程度较大、平均海拔较高但水体覆盖率较低的区域。
对比文化多样性与生物多样性的影响因素发现, 各因素对两种多样性所产生的影响较为相似。因此, 在各因素的基础上增加生物多样性作为变量对文化多样性进行探索性回归分析, 其变量显著性也达到100%, 反映出生物多样性对文化多样性的空间分布产生强烈影响。同时, Pearson相关分析也表明, 基于县级单元的生物多样性与文化多样性的相关系数为0.4499, 两者达到极显著(P<0.01)的正相关关系, 反映出西南地区的文化多样性与生物多样性彼此产生相互促进的作用。
各影响因素对生物文化多样性空间分布的研究发现(表 1), 平均坡度的变量显著性为100%, 体现出了研究单元内地形坡度的平均值对生物文化多样性的空间分布具有很强的影响。与文化多样性和生物多样性的影响因素相似, 水体周长、区域面积与水体面积对生物文化多样性的空间分布影响较弱。Pearson相关分析的结果表明, 生物文化多样性与平均坡度、海拔标准差、坡度标准差和平均海拔呈极显著(P<0.01)的正相关关系, 而与水体覆盖率呈极显著(P<0.01)的负相关关系。因此, 西南地区生物文化多样性分布格局受到平均坡度([1.37°, 32.69°])的重要影响, 生物文化多样性高的单元倾向于聚集在地形坡度大且海拔与坡度的差异程度较大、平均海拔较高但水体覆盖率较低的区域。
3 结论与讨论在较大尺度的研究中, 语言常被用来指示文化多样性, 而脊椎动物与植物的数量常被用来指征生物多样性[21]。虽然这些指标并不能完全代表两种多样性, 但考虑到数据的可获得性, 它们仍在定量研究中具有优势。然而, 用单一的生物多样性或文化多样性的指标来反映世界的多样性都是不全面的。因而, 为量化国家或者地区尺度的多样性水平, 学者们引入了一些测度生物文化多样性的指标, 一般的做法是将文化多样性和生物多样性的定量化指标加以综合来表征整体的多样性[2]。例如, Loh与Harmon以宗教、语言和种族数量来测度文化多样性, 同时以鸟类、哺乳动物和植物数量来测度生物多样性, 在此基础上将两者的算术平均作为生物文化多样性指数, 并以国家为单元进行了多样性的综合评价[6]。对本研究的西南地区而言, 一方面, 由于中央政府在进行民族识别的过程中考虑到了民族语言、宗教信仰和历史渊源等因素[22], 因而利用民族人口数量来衡量研究单元的文化多样性水平具有指示性[23];另一方面, 包括野生动物与野生植物在内的指示物种数目及其受威胁状态也反映了区域的生物资源水平, 用以衡量研究单元的生物多样性水平具有一定的代表性[24]。基于此, 通过选取合适的无量纲化方法以消除两种指标在变异程度上的差异并求和所得到的生物文化多样性指标具有一定的科学性与实用性。
本研究对西南地区文化多样性与生物多样性的分布格局对比发现, 两者存在一定的空间分布差异, 但也存在明显的高低值聚类重叠区域。这反映出文化多样性与生物多样性具有密切联系, 并表现在地理空间上的交叉重合, 与Moore等[25]和Sutherland[26]的研究结果相吻合。通过提取河流干流处文化多样性、生物多样性与生物文化多样性的数值, 对比发现三者在研究区内沿河流流向大多呈现“低-高”或“低-高-低”的变化状态。西南地区各流域上游的文化多样性普遍比下游低, 是因为上游区域一般是高海拔地区不适宜大规模的人口定居。随后, 长江干流与珠江干流在流经山地与丘陵之后进入平原区, 该区域土地平整且水资源丰沛吸引了汉族、壮族等多人口民族的定居, 使得区域的文化多样性降低。与文化多样性的变化原因类似, 西南地区各流域上游过高的海拔对生物栖息也并不有利, 而中部的山地与丘陵地带为生物提供多种生境条件, 但平原区因自然条件较为单一且人口密度增大导致的人为干扰增强。文化多样性与生物多样性的交叉重合, 使得西南地区生物文化多样性沿河流流向呈现出“低-高”或“低-高-低”的变化状态。
伴随全球贸易和文化壁垒的打破, 人们赖以生存的多样化的文化与生态景观双双面临严重威胁[7]。利益驱动的高强度资源开发, 不仅造成了生物资源的减少, 也破坏了传统文化的物质基础[27]。农产品的工业化经营使得原有的多元生产模式转换为单一物种培植, 降低了农田生态系统的生物多样性, 并减少了适宜的物种栖息地[28]。与此同时, 传统的作物培育知识与饮食结构, 以及在此过程中提炼得到的生态知识、技巧和实践也随之消失[29]。此外, 极端的自然灾害事件和环境损害事故也是造成生物多样性减少和基于空间的本土文化受到侵蚀的重要原因[30]。本研究的结果表明, 西南地区文化多样性与生物多样性具有相似的格局和影响因素, 存在密切的联系。因此, 西南地区的多样性保护应统筹兼顾文化多样性与生物多样性两方面, 更多从生物文化多样性的视角或者途径来开展相关研究与保护实践。
| [1] |
蒋志刚, 马克平, 韩兴国. 保护生物学. 杭州: 浙江科学技术出版社, 1997.
|
| [2] |
毛舒欣, 沈园, 邓红兵. 生物文化多样性研究进展. 生态学报, 2017, 37(24): 8179-8186. |
| [3] |
Posey D A. Cultural and spiritual values of biodiversity: a complementary contribution to the global biodiversity assessment//Posey D A, ed. Cultural and Spiritual Values of Biodiversity. London: United Nations Environment Programme and Intermediate Technology Publications, 1999.
|
| [4] |
王娟, 杨宇明, 王泾, 石明, 裴盛基.试论文化多样性与生物多样性的相互关系//中国生物多样性保护与研究进展Ⅵ——第六届全国生物多样性保护与持续利用研讨会论文集.丽江: 中国科学院生物多样性委员会, 2004: 57-68.
|
| [5] |
季维智. 保护生物学基础. 北京: 中国林业出版社, 2000.
|
| [6] |
Loh J, Harmon D. A global index of biocultural diversity. Ecological Indicators, 2005, 5(3): 231-241. DOI:10.1016/j.ecolind.2005.02.005 |
| [7] |
Pretty J, Adams B, Berkes F, De Athayde S F, Dudley N, Hunn E, Maffi L, Milton K, Rapport D, Robbins P, Sterling E, Stolton S, Tsing A, Vintinner E, Pilgrim S. The intersections of biological diversity and cultural diversity:towards integration. Conservation and Society, 2009, 7(2): 100-112. DOI:10.4103/0972-4923.58642 |
| [8] |
Gavin M C, McCarter J, Mead A, Berkes F, Stepp J R, Peterson D, Tang R F. Defining biocultural approaches to conservation. Trends in Ecology & Evolution, 2015, 30(3): 140-145. |
| [9] |
李德铢, 杨湘云, 王雨华, 蔡杰. 中国西南野生生物种质资源库. 中国科学院院刊, 2010, 25(5): 565-569, 550-550. DOI:10.3969/j.issn.1000-3045.2010.05.014 |
| [10] |
杨圣敏, 丁宏. 中国民族志. 北京: 中央民族大学出版社, 2003.
|
| [11] |
王恩涌, 胡兆量, 周尚意, 赫维红, 刘岩. 中国文化地理. 北京: 科学出版社, 2008.
|
| [12] |
沈园, 毛舒欣, 邱莎, 李涛, 邓红兵. 西南地区文化多样性时空格局研究. 生态学报, 2018, 38(21): 1-11. DOI:10.5846/stxb201802060315 |
| [13] |
渠爱雪, 孟召宜. 我国文化多样性时空格局及其成因研究. 人文地理, 2014, 29(6): 53-59, 124-124. |
| [14] |
国家统计局人口和就业统计司, 国家民族事务委员会经济发展司. 中国2010年人口普查分民族人口资料(下). 北京: 民族出版社, 2013.
|
| [15] |
张路, 肖燚, 郑华, 徐卫华, 逯非, 江凌, 饶恩明, 肖洋, 吴炳方, 曾源, 欧阳志云. 2010年中国生态系统服务空间数据集. (2017-07-10)[2018-3-28] http://www.csdata.org/p/106/#history.
|
| [16] |
蒋敏. 中国省域交通与城市化的耦合度分析. 新疆社会科学, 2008(5): 19-24. DOI:10.3969/j.issn.1009-5330.2008.05.005 |
| [17] |
Ord J K, Getis A. Local spatial autocorrelation statistics:distributional issues and an application. Geographical Analysis, 1995, 27(4): 286-306. |
| [18] |
Mitchell A. The ESRI Guide to GIS Analysis. Redlands, CA: ESRI Press, 2005.
|
| [19] |
Duque J C, Ramos R, Suriñach J. Supervised regionalization methods:a survey. International Regional Science Review, 2007, 30(3): 195-220. DOI:10.1177/0160017607301605 |
| [20] |
IBM. IBM SPSS Statistics for Windows, Version 20. Armonk, NY: IBM Corporation, 2011.
|
| [21] |
Maffi L. On Biocultural Diversity:Linking Language, Knowledge, and the Environment. Washington, DC: Smithsonian Institution Press, 2001.
|
| [22] |
尤伟琼.云南民族识别研究[D].昆明: 云南大学, 2012.
|
| [23] |
孟召宜, 沈正平, 渠爱雪, 马晓冬. 文化多样性研究述评与展望. 淮海工学院学报:人文社会科学版, 2015, 13(4): 74-81. |
| [24] |
周红章. 物种与物种多样性. 生物多样性, 2000, 8(2): 215-226. DOI:10.3321/j.issn:1005-0094.2000.02.011 |
| [25] |
Moore J L, Manne L, Brooks T, Burgess N D, Davies R, Rahbek C, Williams P, Balmford A. The distribution of cultural and biological diversity in Africa. Proceedings of the Royal Society B:Biological Sciences, 2002, 269(1501): 1645-1653. DOI:10.1098/rspb.2002.2075 |
| [26] |
Sutherland W J. Parallel extinction risk and global distribution of languages and species. Nature, 2003, 423(6937): 276-279. DOI:10.1038/nature01607 |
| [27] |
刘国华. 西南生态安全格局形成机制及演变机理. 生态学报, 2016, 36(22): 7088-7091. |
| [28] |
Samson C, Pretty J. Environmental and health benefits of hunting lifestyles and diets for the Innu of Labrador. Food Policy, 2006, 31(6): 528-553. DOI:10.1016/j.foodpol.2006.02.001 |
| [29] |
Pilgrim S E, Cullen L C, Smith D J, Pretty J. Ecological knowledge is lost in wealthier communities and countries. Environmental Science and Technology, 2008, 42(4): 1004-1009. DOI:10.1021/es070837v |
| [30] |
Rapport D J, Whitford W G. How ecosystems respond to stress:common properties of arid and aquatic systems. BioScience, 1999, 49(3): 193-203. DOI:10.2307/1313509 |