 2019, Vol. 39
2019, Vol. 39文章信息
- 宋楠, 李新蓉, 狄林楠
- SONG Nan, LI Xinrong, DI Linnan
- 荒漠孑遗植物裸果木种子时空扩散特性
- Temporal and spatial characteristics of seed dispersal of the relict Gymnocarpos przewalskii in deserts
- 生态学报. 2019, 39(7): 2462-2469
- Acta Ecologica Sinica. 2019, 39(7): 2462-2469
- http://dx.doi.org/10.5846/stxb201803290640
-
文章历史
- 收稿日期: 2018-03-29
- 网络出版日期: 2019-01-10

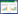




种子扩散是植物的繁殖体远离母株扩散出去的过程, 是植物主要的运动机制, 是植物生活史重要的组成部分[1-4]。荒漠植物的种子扩散策略是生活史中重要的环节, 也是其对严酷的环境条件的一种适应[5]。种子扩散受自然选择的影响强烈, 外界环境(风向、降水和媒介动物等)和植物的自身特性是影响种子扩散的主要因素, 测定种子扩散参数比植物生活史中其他大多数参数更困难[6-8]。不同物种及同一物种的不同种群种子扩散的时间动态和空间格局可能存在差异, 这对种子萌发和幼苗建成的时机及种群的分布具有深远的影响, 是物种或种群对其生境长期适应的结果[9-13], 例如生长在古尔班通古特沙漠的70种植物的种子共有5种不同的扩散方式[14], 蒙古扁桃的种子除了依靠自身重力扩散, 动物和雨水也是其扩散媒介[15], 霸王生长的地形条件、邻近的植物以及风向等影响该物种种子扩散时空特征[16]。
裸果木(Gymnocarpos przewalskii), 属于石竹科(Caryophyllaceae)裸果木属(Gymnocarpos)亚灌木, 稀有种, 亚洲中部荒漠区鲜有第三纪孑遗物种, 属古地中海旱生植物区系成分, 其生存和演化发展过程对研究古地中海气候变化和旱生植物演化过程极为重要[17]。近年来, 由于裸果木植物种群遭羊群啃食、牧民樵采、分布区内自然环境不断恶化及结籽率较低等影响, 数量明显减少, 分布范围不断缩小以及破碎化。植物种子从母株开始扩散, 能够提高种子存活的几率, 使其到达适宜萌发和生长的微生境, 帮助种群开辟新生境[18-19], 对维持物种生存及生物多样性至关重要。目前, 对裸果木的研究主要集中在种群结构[20-21]、繁育系统[22]、开花物候[23]和种子萌发[24]等方面, 有关种子扩散时空特性的研究尚未见报道。
本文通过布设种子收集器等方法, 对裸果木哈密种群种子扩散的时空动态进行连续定点观测, 目的是了解其种子在极端环境下的扩散特性及对环境的适应性, 为进一步掌握该物种生活史特征及种群动态提供理论资料。
1 研究区和材料研究地点位于新疆哈密地区三道岭(92°33.171′E, 43°15.557′N), 海拔1323 m, 属新疆东部极端干旱区。日照充足, 年均日照3358 h, 夏季酷热, 冬季寒冷, 昼夜温差大, 年平均气温9.8℃, 全年中7月温度最高。多风, 常年盛行北风和西北风。年平均降水量为33.8 mm, 降水稀少且降水量在年度内分布不均, 降水主要集中在6月、7月(图 1)。蒸发强烈, 年均蒸发量3300 mm, 年均无霜期约182 d。气候类型为典型的温带大陆性干旱气候, 土壤类型为石质性很强的棕色石膏荒漠土, 主要伴生种有霸王(Sarcozygium xanthoxylon)、琵琶柴(Reaumuria songonica)、松叶猪毛菜(Salsola laricifolia)、盐生草(Halogeton glomeratus)和石生驼蹄瓣(Zygophyllum rosovii)等。

|
| 图 1 2015年、2016年和10年(2007—2016年)观测区域的月降水量及月平均气温 Fig. 1 The monthly rainfall and average monthly temperature at the studied area in 2015, 2016 and 10 years (2007—2016) 资料源自中国气象数据网(http://data.cma.cn) |
裸果木为多年生亚灌木, 株高约13—80 cm, 东西冠幅约10—124 cm, 南北冠幅约9—125 cm, 植物无明显主茎, 茎曲折且多分枝, 节膨大。裸果木3月底至4月初长嫩叶, 4月下旬到5月上旬进入初花期, 聚伞花序腋生, 花小, 不显著, 每个花序约有单花5—15朵不等, 具宽椭圆形的白色膜质苞片, 苞片易碎、易脱落, 花萼宿存且下部连合, 萼片呈倒披针形, 数量为5, 萼片长度约2.30 mm。果期6月, 瘦果包于宿存萼内, 自然结实率低, 种子小, 长圆形, 表皮光滑, 呈褐色。
2 研究方法 2.1 种子收集器布设及种子扩散观察2015年和2016年每年的5月下旬, 选取所处地形条件(海拔、坡度和坡向)和植株大小(株高和冠幅)基本一致, 且20 m范围内无其他株丛分布的3株孤立的裸果木植株为观察对象, 布设种子收集器。收集器由半径0.075 m的白色圆形PVC管和孔径1×10-3 m的尼龙网组成。具体布设方法如下:以裸果木株丛基部为起始点, 用罗盘确定方向, 分别沿东、南、西、北、东南、东北、西南、西北8个方向的延伸线布设收集器, 每个方向均以0.1 m为间隔布设收集器, 前期野外观察发现, 裸果木种子多在植株周围扩散, 故最后一个收集器到母株的距离是3 m, 各方向均有13个收集器, 3株植株总计布设种子收集器312个。裸果木种子扩散的时间和空间变化动态观测具体方法见2.1.1和2.1.2。
2.1.1 种子扩散时间动态观察在裸果木结实期间开始观察种子扩散情况, 收集器内第1粒种子出现的时间记为种子扩散开始时间, 每5 d观察统计一次所有收集器内裸果木种子数量, 并将收集器内的种子移出, 直至连续15 d收集器内均不再有种子出现, 视为其种群种子完成扩散过程。同时, 收集观测期间每日的温度、降水量及风速, 数据均来自中国气象数据网(http://data.cma.cn)。将收集器里的种子数量换算为种子扩散密度(1 m2单位面积扩散的种子数量), 种子扩散密度=收集器内种子数量/收集器面积, 粒/m2;收集器面积=π×0.0752, m2。
2.1.2 种子扩散空间动态观察为探讨风向对裸果木种子扩散的影响, 依次对东、南、西、北、东南、东北、西南和西北8个不同方向种子收集器内的种子数量进行观察和统计, 每个方向的种子扩散数是各方向13个收集器中种子数总和。同时, 为了解裸果木的种子扩散数量与其距母株扩散距离之间关系, 依次对各不同方向距母株0、0.25、0.50、0.75、1、1.25、1.50、1.75、2、2.25、2.50、2.75 m和3 m收集器内的种子数量进行观察统计。种子扩散数量均换算成种子扩散密度, 方法同2.1.1, 各种子在不同距离的扩散百分比=各方向上某距离范围内种子扩散数/某方向种子扩散总数×100%。
2.2 数据处理所有数据在Microsoft Excel 2007中进行整理, 采用SPSS 20.0对数据进行分析, 所有数据首先用K-S检验数据分布类型, 所有数据均呈正态分布。随后, 采用单因素方差分析(One-way ANOVA)比较不同时期种子扩散密度、不同方向种子扩散密度、不同扩散距离种子扩散密度以及不同年份种子扩散密度的差异;最小显著差异法(LSD)检验其差异性是否达到显著水平(P≤0.05), 用Spearman双侧相关性分析研究不同方向种子扩散密度与扩散距离的关系(P≤0.01)。采用Origin Pro 8.5进行绘图。统计结果均以平均值±标准差(mean±SD)表示。
3 结果与分析 3.1 种子扩散的时间变化特征裸果木哈密种群种子于每年5月底成熟, 随后开始扩散, 由图 2可知, 2015年和2016年种子开始扩散时间分别是6月6日和6月11日, 2015年种子开始扩散的时间略早于2016年, 2015年6月21日种子扩散进入高峰期, 比2016年提前5 d, 种子扩散密度达到峰值后逐渐减少, 两年的种子扩散趋势均呈单峰曲线, 扩散持续时间分别约50 d(6月6日—7月21日)和45 d(6月11日—7月21日);2015年和2016年种子平均扩散密度分别为(396.32±82.03)粒/m2和(236.53±49.84)粒/m2, 二者间不存在显著差异(F=2.7, P>0.05);2015年和2016年种子集中扩散期分别是6月21日—7月1日和6月26日—7月6日;扩散数量分别占扩散总量的68.57%和69.15%, 这说明虽然不同年份的种子扩散进程相似, 不同年份种子集中扩散期的扩散密度分别与当年其他时期种子扩散密度存在显著差异(2015:F=143.06, P<0.05;2016:F=98.523, P<0.05)。同时, 从图 2可知, 2015年、2016年观测期间平均温度范围约19—33℃, 平均风速范围约0—3.6 m/s, 平均温度和平均风速波动不大, 降水量均呈“双峰曲线”, 2015年峰值分别是6月21日和7月6日, 降水量为22.1 mm和10.5 mm, 2016年峰值分别是6月26日和7月11日, 降水量为6.4 mm和18.3 mm, 降水量的第一个高峰期与种子扩散高峰期相吻合。

|
| 图 2 2015年和2016年裸果木种子扩散密度、降水量、温度及风速随时间的变化 Fig. 2 Temporal variation of the seeds dispersal density of Gymnocarpos przewalskii, rainfall, temperature and wind speed during the studied period in 2015 and 2016 |
由图 3可知, 2015年正南和东南方向种子扩散密度分别为(1641.90±149.80)粒/m2和(1396.56±279.29)粒/m2, 2016年正南和东南方向种子扩散密度分别为(849.26±56.62)粒/m2和(792.65±163.44)粒/m2, 2015年和2016年均是正南和东南方向种子扩散密度大, 且两年均与其他各方向上种子扩散密度间存在显著差异(2015:F=57.42, P<0.05;2016:F=17.36, P<0.05);2015年正北和西北方向种子扩散密度分别为(207.60±32.69)粒/m2和(264.22±32.69)粒/m2, 2016年正北和西北方向种子扩散密度分别为(150.90±32.82)粒/m2和(150.98±32.69)粒/m2, 2015年和2016年正北和西北方向种子扩散密度分别与正西和东北方向种子扩散密度差异不显著(P>0.05), 与其他4个方向上种子扩散密度差异显著(P<0.05);2015年和2016年正南方向种子扩散密度存在显著差异(F=58.80, P<0.05), 东南方向种子扩散密度差异不显著(F=6.92, P>0.05), 正北方向种子扩散密度差异不显著(F=4.49, P>0.05), 西北方向种子扩散密度存在显著差异(F=18.00, P<0.05)。

|
| 图 3 2015年和2016年裸果木不同方向的种子扩散密度 Fig. 3 The seeds dispersal density of Gymnocarpos przewalskii in different directions in 2015 and 2016 |
裸果木种子自植株扩散后, 在母株周围分布, 种子扩散密度与距离之间存在非线性的负相关(图 4), 2015年和2016年种子扩散密度均在母株下最大(距母株0 m), 种子最远扩散距离分别是2 m和1.50 m, 种子扩散分别集中在距母株0.75 m和0.50 m范围内, 分别占总扩散量的91.35%和86.36%, 2015年距母株0 m、0.25 m、0.50 m和0.75 m范围内的种子扩散密度分别为(2981.84±182.00)粒/m2、(1434.31±172.97)粒/m2、(603.91±86.49)粒/m2和(358.57±32.68)粒/m2, 2016年距母株0 m、0.25 m和0.50 m范围内的种子扩散密度分别为(1755.12±315.23)粒/m2、(754.9±68.05)粒/m2和(358.57±49.93)粒/m2, 2015年和2016年集中扩散范围内的种子扩散密度均与其他各距离间差异显著(2015:F=353.87, P<0.05;2016:F=79.16, P<0.05);2015年和2016年距母株0 m、0.25 m、0.50 m和0.75 m时, 两年间种子扩散密度均存在显著差异(0 m:F=34.07, P<0.05;0.25 m:F=31.61, P<0.05;0.50 m:F=12.07, P<0.05;0.75 m:F=40.50, P<0.05)。

|
| 图 4 2015年和2016年裸果木距母株不同扩散距离的种子密度 Fig. 4 The seeds density in different distances from mother plant of Gymnocarpos przewalskii in 2015 and 2016 |
由表 1可知, 2015年、2016年种子在正北、西北、东北和正西方向全部扩散在距母株0.25 m范围内, 正南方向种子扩散最远, 扩散距离分别为2 m和1.50 m;在距母株1 m范围内, 正南、东南、西南和正东方向的种子扩散数量均占该方向上总扩散数量的85%以上;2015年、2016年每年不同方向种子扩散距离存在极显著差异(2015:F=4.88, P<0.01;2016:F=4.16, P<0.01), 各方向种子扩散密度和扩散距离呈极显著非线性负相关。
| 年份 Year |
方向 Direction |
最大扩散距离 Max distance/m |
种子扩散百分比/% Percentage of seed dispersal |
种子扩散密度和 扩散距离的相关系数Spearman correlation coefficients of seed density and distance |
|||
| 0—0.25 m | 0—0.50 m | 0—0.75 m | 0—1.00 m | ||||
| 2015 | 北 | 0.25 | 100 | 100 | 100 | 100 | -0.58** |
| 东北 | 0.25 | 100 | 100 | 100 | 100 | -0.63** | |
| 东 | 1.00 | 75 | 87.5 | 95 | 100 | -0.85** | |
| 东南 | 1.75 | 63.51 | 77.03 | 85.13 | 90.54 | -0.95** | |
| 南 | 2.00 | 63.22 | 77.01 | 83.91 | 88.51 | -0.96** | |
| 西南 | 0.75 | 81.25 | 91.67 | 100 | 100 | -0.82** | |
| 西 | 0.25 | 100 | 100 | 100 | 100 | -0.63** | |
| 西北 | 0.25 | 100 | 100 | 100 | 100 | -0.63** | |
| 2016 | 北 | 0.25 | 100 | 100 | 100 | 100 | -0.58** |
| 东北 | 0.25 | 100 | 100 | 100 | 100 | -0.63** | |
| 东 | 0.75 | 68.18 | 83.68 | 100 | 100 | -0.81** | |
| 东南 | 1.50 | 61.90 | 76.19 | 83.33 | 90.48 | -0.91** | |
| 南 | 1.50 | 64.44 | 77.78 | 84.44 | 91.11 | -0.91** | |
| 西南 | 0.75 | 85.18 | 96.30 | 100 | 100 | -0.76** | |
| 西 | 0.00 | 100 | 100 | 100 | 100 | -0.46** | |
| 西北 | 0.00 | 100 | 100 | 100 | 100 | -0.46** | |
| **在0.01水平(双侧)上显著相关 | |||||||
种子扩散随时间动态变化的过程是植物世代更替的重要环节, 在种群更新中起着重要作用, 不同物种种子扩散特性不同, 这是物种或种群对特定环境长期适应的结果[7, 11]。在本研究中, 虽然不同年份裸果木种子扩散高峰期略有不同, 但是两年的种子扩散进程与大部分荒漠植物例如额济纳胡杨(Populus euphratica)[25]、古尔班通古特沙漠边缘羽毛针禾(Stipagrostis pennata)[13]和浑善达克沙地沙地榆(Ulmus pumila L.var.sabulosa)[26]等相似, 需要经历起始期、高峰期和消退期三个阶段, 扩散趋势呈单峰曲线, 也有一些荒漠植株的种子扩散趋势也呈单峰曲线, 但是其种子在扩散初期就达到最大扩散量, 随后种子扩散密度不断减少, 直至趋于稳定, 例如吕朝燕等[27]研究发现, 准噶尔盆地东南缘梭梭(Haloxylon ammodendron)种子扩散虽然历时约90 d, 但其种子在刚开始扩散时, 就已经达到了最大扩散量。裸果木具有集中大量开花的开花模式, 开花进程呈单峰曲线[23], 不同年份其种子扩散过程均与开花进程相似, 也呈现出集中大量扩散的模式, 一些学者认为植物的开花物候往往与结实物候及种子的扩散进程相联系[28-29], 这可以解释裸果木的开花进程与种子扩散趋势相似, 而不同年份裸果木种子扩散扩散高峰期略有不同, 可能是与当年气温、降水及光照等气象因子的差异有关。在裸果木种子扩散高峰期, 观测区均有一次大量降水, Liu等[14]对70种荒漠植物扩散特性研究的结果表明, 小种子比大种子更加容易被雨水冲洗, 我们认为裸果木种子成熟期间, 一次突发的降水有利于其种子集中扩散, 这种在短时间内大量扩散的策略可以减少蚂蚁、蜥蜴等摄食动物对该物种种子摄取的数量, 从而提高种子在严酷环境条件下的留存几率[30]。杨赵平等[24]对裸果木种子萌发的研究表明, 裸果木种子不休眠, 不同变温条件下的平均萌发时间不到2 d, 30/35℃萌发率为95%, 这表明种子适宜在夏季萌发且具有快速萌发的萌发策略。一般而言, 伴随降水进行种子扩散的物种, 其种子往往是非休眠的, 如果有足够的土壤湿度, 种子会快速发芽[31]。观测地点7月温度最高, 6月和7月降水量最大, 且与该地区10年(2007年—2016年)的温度和降水量波动不大, 7月份大部分裸果木种子已经完成了其扩散过程, 此时, 观测区降水频繁且温度适宜, 满足该物种种子萌发的条件, 可以确保扩散的种子抓住机会在合适的条件下萌发, 这在一定程度上提高了种子萌发的几率, 有利于其实现种群的更新, 同时, 降水所形成的地表径流有助于种子进行二次扩散。通过野外观察发现, 2015年和2016年, 雨后在裸果木植株下均有少量的幼苗, 我们认为该物种在大量降水前完成种子扩散, 是对其生境长期适应的结果。
研究区盛行北风和西北风, 因此正南和东南方向为顺风方向, 正北和西北方向为逆风方向, 不同年份裸果木种子均密集扩散在顺风的正南和东南方向, 且扩散距离远, 逆风的正北和西北方向种子扩散密度小, 且扩散距离较近。Heydel等[32]提出, 风的方向和强度的变化会影响种子在时间和空间上的扩散模式, 这可以解释不同年份裸果木不同方向扩散密度及不同方向上种子扩散距离的差异, 因此, 风是其种子扩散的关键媒介之一。在微生境尺度上, 一些植物的种子扩散与幼苗库具有一致性[33], 关于裸果木种子扩散和实生苗定居方面的问题还有待进一步的研究。在本研究中, 虽然2015年和2016年各方向裸果木种子扩散距离不同, 但其种子均集中扩散在植物冠幅内, 随着距母株距离的增加, 种子扩散密度减少, 扩散密度与扩散距离呈极显著的负相关关系(P<0.01), 这种以母株为中心的扩散格局与阔叶红松(Pinus koraiensis)、梭梭以及差巴嘎蒿(Artemisia halodendron)等植物的扩散格局相似[12, 27, 34]。植物种子扩散的空间通常是非常有限的, 扩散距离取决于植物特性以及环境条件, 并且在时间和空间上变化很大[6]。裸果木没有明显的主茎, 枝条繁多且极为密集, 对植株内部的风力起到了一定程度的削弱作用, 从而导致种子集中扩散在母株附近并呈现植株成簇生长的现象。有学者认为, 一些种子质量小且具有扩散结构(冠毛、果翅、苞片等)的植物果实在成熟后能够扩散到更远地方, 从而扩大物种的分布范围[35], 裸果木种子质量小且具有四棱形的宿存萼和膜质苞片, 尽管我们观察到裸果木种子扩散距离较短, 但其一次扩散的种子很有可能会在风力的作用下进行二次扩散占领较远的地域。
| [1] |
Levine J M, Murrell D J. The community-level consequences of seed dispersal patterns. Annual Review of Ecology, Evolution, and Systematics, 2003, 34: 549-574. DOI:10.1146/annurev.ecolsys.34.011802.132400 |
| [2] |
Nathan R, Schurr F M, Spiegel O, Steinitz O, Trakhtenbrot A, Soar A. Mechanisms of long-distance seed dispersal. Trends in Ecology & Evolution, 2008, 23(11): 638-647. |
| [3] |
Chanyenga T F, Geldenhuys C J, Harvey J. Variation in seed rain from Widdringtonia whytei growing in different conditions on Mulanje Mountain in Malawi. Southern Forests:A Journal of Forest Science, 2011, 73(3/4): 123-129. |
| [4] |
朱金雷, 刘志民. 种子传播生物学主要术语和概念. 生态学杂志, 2012, 31(9): 2397-2403. |
| [5] |
Gutterman Y. Strategies of seed dispersal and germination in plants inhabiting deserts. The Botanical Review, 1994, 60(4): 373-425. DOI:10.1007/BF02857924 |
| [6] |
Howe H F, Smallwood J. Ecology of seed dispersal. Annual Review of Ecology and Systematics, 1982, 13: 201-228. DOI:10.1146/annurev.es.13.110182.001221 |
| [7] |
Wang B C, Smith T B. Closing the seed dispersal loop. Trends in Ecology & Evolution, 2002, 17(8): 379-386. |
| [8] |
Bullock J M, Shea K, Skarpaas O. Measuring plant dispersal:an introduction to field methods and experimental design. Plant Ecology, 2006, 186(2): 217-234. DOI:10.1007/s11258-006-9124-5 |
| [9] |
Clark J S, Silman M, Kern R, Macklin E, Hillerislambers J. Seed dispersal near and far:patterns across temperate and tropical forests. Ecology, 1999, 80(5): 1475-1494. DOI:10.1890/0012-9658(1999)080[1475:SDNAFP]2.0.CO;2 |
| [10] |
Howe H F, Miriti M N. No question:seed dispersal matters. Trends in Ecology & Evolution, 2000, 15(11): 434-436. |
| [11] |
胡星明, 蔡永立, 李恺, 张天澍. 浙江天童常绿阔叶林栲树种子雨的时空格局. 应用生态学报, 2005, 16(5): 815-819. DOI:10.3321/j.issn:1001-9332.2005.05.008 |
| [12] |
刘双, 金光泽. 小兴安岭阔叶红松(Pinus koraiensis)林种子雨的时空动态. 生态学报, 2008, 28(11): 5731-5740. DOI:10.3321/j.issn:1000-0933.2008.11.060 |
| [13] |
谷伟, 岳永杰, 李钢铁, 潘文杰, 哈丽雅. 浑善达克沙地沙地榆种子雨的扩散规律. 生态学报, 2012, 32(11): 3440-3448. |
| [14] |
Liu H L, Zhang D Y, Yang X J, Huang Z Y, Duan S M, Wang X Y. Seed dispersal and germination traits of 70 plant species inhabiting the Gurbantunggut desert in northwest China. The Scientific World Journal, 2014, 2014: 346405. |
| [15] |
王进, 颜霞, 李军元, 张勇, 谢全刚, 李刚, 顾文豪. 蒙古扁桃种子雨散布及种子萌发特性研究. 冰川冻土, 2017, 39(2): 429-434. |
| [16] |
Zhao X Y, Baskin C C, Zhu C Q, Baskin J M. Fruit dispersal dynamics of the cold desert shrub Zygophyllum xanthoxylon. Seed Science Research, 2017, 27(4): 303-310. DOI:10.1017/S0960258517000253 |
| [17] |
柴永青, 曹致中, 蔡卓山, 万丽霞, 李学玲. 肃北地区稀有植物裸果木种群的空间分布格局. 草业学报, 2010, 19(5): 239-249. |
| [18] |
赵哈林. 沙漠化过程中植物的适应对策及植被稳定性机理. 北京: 海洋出版社, 2004: 91-96.
|
| [19] |
Trakhtenbrot A, Katul G G, Nathan R. Mechanistic modeling of seed dispersal by wind over hilly terrain. Ecological Modelling, 2014, 274: 29-40. DOI:10.1016/j.ecolmodel.2013.11.029 |
| [20] |
王立龙, 王亮, 张丽芳, 刘玉洋, 徐世健. 不同生境下濒危植物裸果木种群结构及动态特征. 植物生态学报, 2015, 39(10): 980-989. DOI:10.17521/cjpe.2015.0095 |
| [21] |
王博, 王亮, 王立龙, 杨增武, 姚静斐, 徐世健. 濒危植物裸果木(Gymnocarpos przewalskii)与其伴生种种间联结性及群落稳定性. 中国沙漠, 2017, 37(1): 86-92. |
| [22] |
李新蓉, 唐欣, 付文洁. 荒漠孑遗植物裸果木的花部特征及繁育系统. 生态学杂志, 2016, 35(10): 2592-2598. |
| [23] |
唐欣, 李新蓉. 荒漠孑遗植物裸果木的开花物候特征. 植物学报, 2017, 52(4): 487-495. |
| [24] |
杨赵平, 梁继业, 柴学平, 薛婉婷. 环境因子对珍稀濒危植物裸果木种子萌发特性的影响. 西南林业大学学报, 2017, 37(1): 49-53. |
| [25] |
张玉波, 李景文, 张昊, 邹大林, 武逢平, 程春龙, 李俊清, 李帅英. 胡杨种子散布的时空分布格局. 生态学报, 2005, 25(8): 1994-2000. DOI:10.3321/j.issn:1000-0933.2005.08.024 |
| [26] |
龙丽红, 王慧, 马晓丽, 徐鸿斌, 王绍明. 古尔班通古特沙漠羽毛针禾(Stipagrostis pennata)种群种子雨特征. 干旱区研究, 2014, 31(3): 516-522. |
| [27] |
吕朝燕, 张希明, 刘国军, 吴俊侠, 闫海龙, 邓潮洲. 准噶尔盆地东南缘梭梭种子雨特征. 生态学报, 2012, 32(19): 6270-6278. |
| [28] |
Culot L, Muñoz Lazo F J J, Huynen M C, Poncin P, Heymann E W. Seasonal variation in seed dispersal by tamarins alters seed rain in a secondary rain forest. International Journal of Primatology, 2010, 31(4): 553-569. DOI:10.1007/s10764-010-9413-7 |
| [29] |
李娟, 郭聪, 肖治术. 都江堰亚热带森林常见木本植物果实组成与种子扩散策略. 生物多样性, 2013, 21(5): 572-581. |
| [30] |
查同刚, 孙向阳, 王登芝, 阎海平. 北京西山地区人工侧柏林种子雨的研究. 北京林业大学学报, 2003, 25(1): 28-31. DOI:10.3321/j.issn:1000-1522.2003.01.007 |
| [31] |
Ramos D M, Diniz P, Ooi M K J, Borghetti F, Valls J F M. Avoiding the dry season:dispersal time and syndrome mediate seed dormancy in grasses in Neotropical savanna and wet grasslands. Journal of Vegetation Science, 2017, 28(4): 798-807. DOI:10.1111/jvs.2017.28.issue-4 |
| [32] |
Heydel F, Cunze S, Bernhardt-Römermann M, Tackenberg O. Long-distance seed dispersal by wind:disentangling the effects of species traits, vegetation types, vertical turbulence and wind speed. Ecological Research, 2014, 29(4): 641-651. DOI:10.1007/s11284-014-1142-5 |
| [33] |
于顺利, 郎南军, 彭明俊, 赵琳, 郭永清, 郑科, 张立新, 温绍龙, 李晖. 种子雨研究进展. 生态学杂志, 2007, 26(10): 1646-1652. |
| [34] |
赵丽娅, 李兆华, 赵锦慧, 赵哈林, 李锋瑞. 流动沙丘先锋灌木差巴嘎蒿的种子散布格局. 西北植物学报, 2005, 25(9): 1821-1826. DOI:10.3321/j.issn:1000-4025.2005.09.020 |
| [35] |
白成科, 曹博, 李桂双. 植物种子传播途径与基因组值和千粒重的相关性. 生态学杂志, 2013, 32(4): 832-837. |