 2019, Vol. 39
2019, Vol. 39文章信息
- 王亚林, 丁忆, 胡艳, 陈静, 范文武
- WANG Yalin, DING Yi, HU Yan, CHEN Jing, FAN Wenwu
- 中国灌木生态系统的干旱化趋势及其对植被生长的影响
- The study of shrubland drought dynamics in China and its impacts on vegetation growth
- 生态学报. 2019, 39(6): 2054-2062
- Acta Ecologica Sinica. 2019, 39(6): 2054-2062
- http://dx.doi.org/10.5846/stxb201803300699
-
文章历史
- 收稿日期: 2018-03-30
- 网络出版日期: 2018-12-21

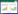




研究表明, 未来中国乃至全球的气温将会持续升高, 干旱化将越来越严重[1-3]。干旱化使得全球植被的NPP(Net Primary Production, NPP)减少[4], 也使得中国的植被在经历了1980—1990年代的快速增长以后生长减缓甚至停滞[5-7]。灌木与乔木和草本相比具有更强的抗旱能力[8-10], 且在中国有着广泛的分布, 从亚热带到温带再到高寒地区均有分布, 其总面积超过了我国陆地面积的20%。在全球变暖干旱加剧的大背景下, 灌木生态系统将会在未来的区域乃至全球生态系统过程中发挥重要作用。那么, 中国灌木生态系统的干旱状况在过去几十年具体有怎样的变化趋势, 灌木生态系统EVI(Enhanced Vegetation Index, EVI)对于干旱变化的响应如何, 不同的灌木生态系统之间是否又具有不同的差异?由于目前对灌木抗旱性的研究大多集中在北方的干旱半干旱地区[9, 11-12], 我国南方湿润地区的灌木对干旱的响应是否也与干旱地区的灌木相一致呢?对这些问题的研究, 将有助于我们对区域生态系统过程变化的理解。
本文根据全国1:100万植被图提取了6类主要灌木生态系统类型[13]:高寒荒漠灌木半灌木(High-cold Desert shrubland, DSRTHC)、温带荒漠灌木半灌木(Temperate Desert shrubland, DSRTTP)、温带落叶灌木(Temperate Deciduous shrubland, DCDSTP)、亚高山常绿灌木(Subalpine evergreen shrubland, EVGNMT)、亚高山落叶灌木(Subalpine deciduous shrubland, DCDSMT)和亚热带常绿灌木(Subtropical evergreen shrubland, EVNGST)。本文的主要研究目标为:(1)基于SPEI干旱指数分析中国灌木生态系统干旱状况的时空趋势;(2)分析中国灌生态系统EVI对SPEI变化的响应;(3)研究不同区域灌木生态系统EVI对干旱的响应差异。
1 数据和方法 1.1 数据计算SPEI(Standardized Precipitation Evapotranspiration Index, SPEI)所需要的气象数据来源于中国气象数据网, 包括月平均气温、月降水量、经纬度等信息。SPEI的原理是利用降水量与潜在蒸散之间的差值所偏离平均状态的程度来表征一个地区的干旱程度。要计算SPEI需要至少30年时间序列的气候数据, 且数据序列越长计算的结果越可靠[14], 因此, 本文利用尽可能长的气象数据来计算得到SPEI数据集。但由于较早时期中国的气象站点较少且数据缺失严重, 经过筛选本文选择了1961—2013年间的气象数据来计算SPEI。对于SPEI的具体计算方法可参考Vicent-Serrano等人[15]和周丹等人[16]的研究。
SPEI具有多时间尺度的特征, 通常选用1、3、6、12、24、36个月等尺度来计算SPEI。1个月时间尺度的SPEI可以比较清楚的反映出旱涝的细微变化, 而3个月时间尺度的SPEI则可以清晰地反映出季节的干旱情况, 同理, 12个月时间尺度的SPEI则可以反应出年际间的干旱情况。本文利用SPEI-3(即3个月尺度的SPEI)来计算季节SPEI, 因为SPEI-3序列中某一个月相对应的值表示当前月以及之前两个月的综合干旱程度(即5月的SPEI值可以反应春季的干旱程度), 因此, 本文选用每年中第5月、8月、11月以及来年2月的SPEI来表示每年的春、夏、秋、冬季的干旱程度[14]。相应的, 用SPEI-12(12个月尺度的SPEI)序列中每年的第12月的SPEI值来反应每年的干旱程度。
本研究所用EVI数据为16-day, 1 km分辨率的MODIS-EVI(MOD13Q1)数据, 时间序列为2001年至2013年。首先, 对EVI时间序列进行了逐栅格的质量筛选和插值处理[17-19];再利用最大值合成法将16-day影像生成月值影像时间序列;最后, 利用全国6种灌木类型空间分布图进行裁剪得到不同灌木类型的EVI时间序列。另外, 对在研究时间内年平均EVI值小于0.05的像元做剔除处理, 从而减小裸地或者稀疏植被带来的影响[6, 19]。
1.2 数据分析本文用一元线性回归模型对不同类型灌木的SPEI和EVI进行回归分析以获取其变化速率, 其公式可以表示为:
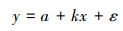
|
式中, y表示SPEI或EVI;x代表年;k为变化速率;a为截距;ε为残差。为了分析中国灌木生态系统SPEI变化趋势是否有显著的转折点, 本文应用分段回归模型来对SPEI序列进行回归分析。分段回归模型已广泛应用于生态学研究中[5-6, 20], 其模型可以表示为:
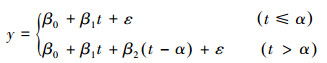
|
式中y为SPEI;t为年;α为时间序列趋势的转折点, 表示时间序列的趋势发生改变的时间;β0为截距;β1和β1+β2分别为转折点前后时间序列的趋势;ε为残差。
为了分析中国灌木生态系统SPEI趋势的空间特征, 本文采用了Theil-Sen slope分析与Mann-Kendall检验相结合的方法来计算SPEI趋势的空间分布。Theil-Sen slope分析为非参数的趋势分析, Mann-Kendall检验为非参数检验, 它们都不要求数据符合正态分布, 且对时间序列中少数的异常值有很高的稳健性, 并且已经广泛应用于植被变化以及气候变化研究中[19, 21-22]。
本文采用多元线性回归模型将年EVI与不同季节SPEI进行多元回归分析, 以分析EVI对不同季节SPEI变化的响应, 其公式可以表示为:
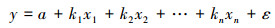
|
式中, y为年平均EVI;a为截距;x为不同季节SPEI;k为拟合的系数;ε为残差。模型采用广义最小二乘法来拟合。由于植被的生长主要发生在生长季, 因此在本文的研究中仅关注生长季的EVI变化, 生长季定义为4—10月[5-6]。由于MODIS-EVI数据最早仅能追溯到2001年, 因此本文从SPEI时间序列中截取2001—2013年间的SPEI时间序列与其相对应的EVI进行分析。
2 结果 2.1 中国灌木生态系统SPEI在1961—2013年间的变化趋势从全国尺度来说, 中国灌木生态系统的SPEI从1961—2013年间呈显著的减小趋势, 说明在这过去的53间年中国灌木生态系统在持续干旱化(表 1)。对于不同的灌木类型来说, 6种类型灌木生态系统的SPEI也均呈显著的减小趋势。
| 灌木生态系统类型 Shrub ecosystem type |
变化速率 Change rate/10-2 SPEI a-1 |
转折点 Turning point |
转折点前变化速率 Change rate before turning point/10-2 SPEI a-1 |
转折点后变化速率 Change rate after turning point/10-2 SPEI a-1 |
| DSRTHC | -1.85*** | 1989* | 0.65 | -4.72*** |
| DSRTTP | -1.72*** | 1988** | 1.02 | -4.81*** |
| DCDSTP | -1.38*** | 2009 | -1.84*** | 6.53*** |
| EVGNMT | -0.51* | 2000 | 0.44 | -5.93** |
| DCDSMT | -0.84*** | 1985 | 1.07* | -2.15*** |
| EVGNST | -0.92*** | 1998 | -0.05 | -4.22*** |
| CONTRY | -1.40*** | 1992*** | 0.23 | -4.33*** |
| DSRTHC:高寒荒漠灌木半灌木, High-cold desert shrubland;DSRTTP, 温带荒漠灌木半灌木, Temperate desert shrubland;DCDSTP, 温带落叶灌木, Temperate deciduous shrubland;EVGNMT, 亚高山常绿灌木, Subalpine evergreen shrubland;DCDSMT, 亚高山落叶灌木, Subalpine deciduous shrubland;EVGNST, 亚热带常绿灌木, Subtropical evergreen shrubland;CONTRY, 全国灌木, Countrywide shrubland; ‘*’, ‘**’和‘***’分别代表P < 0.1, 0.05和0.01 | ||||
本文用分段回归函数对不同灌木生态系统的SPEI进了回归分析, 以检测在过去的53年间其SPEI的变化趋势是否出现显著的改变(表 1, 图 1)。从全国尺度来说, 中国灌木生态系统的SPEI的变化趋势在1992年出现了显著的改变。在1992年以前, 中国灌木生态系统是变湿润的趋势, 其SPEI以0.23×10-2 a-1的速度增加, 在1992年以后中国灌木生态系统的SPEI以-4.33×10-2 a-1(P < 0.01)的速率减小。对于不同的灌木生态系统类型来说, 两种荒漠类型灌木(DSRTHC和DSRTTP)的SPEI分别在1989年和1988年检测到了显著的变化趋势。在转折点之前两种荒漠灌木的SPEI均为不显著的增加趋势, 而在转折点之后, 两种类型灌木的SPEI均显著减小, 而另外4种类型灌木生态系统SPEI的趋势并没有显著的改变。

|
| 图 1 不同灌木生态系统1961—2013年间SPEI的变化趋势 Fig. 1 Temporal trends of different shrub ecosystems SPEI from 1961 to 2013 SPEI:标准化降水蒸散指数, Standardized Precipitation Evapotranspiration Index; 图中实线表示SPEI在整个研究时间段内的变化趋势, 点状虚线表示SPEI在转折点以前的变化趋势, 短线虚线表示SPEI在转折点以后的变化趋势 |
从变化趋势来看, 两种荒漠灌木、两种亚高山灌木的SPEI均从增加转变为减小, 说明这4种类型灌木生态系统所在区域均从之前变湿润趋势转变为显著干旱化的趋势。亚热带常绿灌木的SPEI在1998年之前几乎保持不变, 但之后则呈显著下降趋势。而温带落叶灌木的SPEI的变化趋势与另外5种灌木相反, 由转折点之前的显著下降趋势转变为显著增加趋势, 其值由负变为正, 说明温带落叶灌木所在区域由干旱化转变化湿润化。总的来说, 中国灌木生态系统的干旱状况从1990年代初期以来呈显著的干旱化趋势, 这与我国植被增长在1990年代中期因干旱停滞的趋势相一致[5-7]。
2.2 不同灌木生态系统SPEI和EVI在2001—2013年间的变化趋势由于MODIS-EVI数据的最早日期仅能追溯到2001年, 为了分析EVI对SPEI变化的响应, 本文从不同时间尺度的SPEI序列中截取了2001—2013年的数据序列进行分析(表 2)。6种灌木类型中, 温带落叶灌木的SPEI以6.13×10-2 a-1(P < 0.05)的速度显著增加, 表明温带落叶灌木生态系统在逐渐变湿润, 而亚高山常绿灌木则与之相反, 以-4.59×10-2 a-1的速度显著下降, 表明其正在干旱化。此外, 高寒荒漠、温带荒漠灌木半灌木以及亚热带常绿灌木的SPEI也均呈下降的趋势, 而亚高山落叶灌木的SPEI则呈增加的趋势, 但均不显著。从全国尺度来看, 中国灌木生态系统SPEI在2001—2013年间以-1.88×10-2 a-1(P>0.1)的速度下降, 表明中国灌木生态系统正在逐渐变干旱。在2001—2013年间, 全国灌木生态系统EVI以1.01×10-3 a-1的(P < 0.01)速率显著增加, 其中高寒荒漠灌木半灌木、温带荒漠灌木半灌木、温带落叶灌木和亚热带常绿灌木的EVI均显著增加, 而亚高山常绿灌木的EVI则显著减小, 亚高山落叶灌木的EVI减小但不显著。
| 灌木生态系统类型 Shrub ecosystem type |
年际变化速率 Annual change rate | SPEI季节变化速率 Seasonal SPEI change rate/10-2/a | |||||
| EVI/10-3/a | SPEI/10-2/a | 春 Spring | 夏 Summer | 秋 Autumn | 冬 Winter | ||
| DSRTHC | 0.36** | -1.21 | -2.72 | 1.17 | 1.09 | -3.12 | |
| DSRTTP | 0.55** | -2.48 | -6.20 | 2.50 | -1.06 | 0.05 | |
| DCDSTP | 1.89*** | 6.13** | 4.87* | 5.30* | 5.02 | -1.66 | |
| EVGNMT | -0.64* | -4.59* | -2.28 | -4.85 | 0.23 | -3.43 | |
| DCDSMT | -0.04 | 0.30 | -2.48 | 1.00 | 0.79 | -0.08 | |
| EVGNST | 1.88*** | -2.67 | -2.03 | -4.81** | 4.99** | -5.68* | |
| CONTRY | 1.01*** | -1.88 | -3.71 | 0.02 | 0.91 | -1.77 | |
| 注:EVI:增强型植被指数, Enhanced Vegetation Index;SPEI:标准化降水蒸散指数, Standardized Precipitation Evapotranspiration Index; “*”, “**”和“***”分别表示P < 0.1、0.05和0.01 | |||||||
在春季, 仅温带落叶灌木的SPEI显著增加, 而其他5种类型灌木的SPEI则均呈减小的趋势。在夏季, 温带落叶灌木SPEI显著升高(P < 0.1), 而亚热带常绿灌木SPEI则显著下降(P < 0.05)。此外, 两种荒漠灌木也有增加趋势, 而亚高山常绿灌木则为下降的趋势。亚热带常绿灌木SPEI在秋季显著增加但在冬季显著下降, 而其他几种类型灌木的SPEI在这两个季节都没有显著的变化趋势。从全国来看, SPEI在春、冬两季有减小的趋势, 夏、秋两季有增加的趋势, 但均未达到α=0.1的显著性水平。
2.3 中国灌木生态系统SPEI在2001—2013年间变化趋势的空间特征全国灌木生态系统1.04%的区域的SPEI显著增加, 其主要分布于温带落叶灌木区域(表 3, 图 2)。而SPEI显著减小的面积比例为1.88%, 主分布在云南省以及四川的西南部, 主要为亚高山常绿灌木以及亚热带常绿灌木。SPEI增加但不显著的区域占31.92%, 其主要分布于东北、华北、以及沿海地区, 而减小但不显著的地区占65.16%, 其主要分布于华中、西南以及西北地区。SPEI趋势的空间分布特征与不同灌木类型的变化趋势能够很好地吻合。
| Theil-Sen趋势分析 Theil-Sen slope analysis |
Mann-Kendall检验 Mann-Kendall test |
SPEI变化趋势 SPEI trend |
面积占比/% Area percentage |
| >0 | >1.96 | 显著增加 | 1.04 |
| >0 | -1.96—1.96 | 不显著增加 | 31.92 |
| < 0 | -1.96—1.96 | 不显著减小 | 65.16 |
| < 0 | < -1.96 | 显著减小 | 1.88 |

|
| 图 2 中国灌木生态系统2001—2013年间SPEI变化趋势的空间分布 Fig. 2 Spatial distribution of China shrub ecosystem SPEI variation from 2001 to 2013 不同的圈代表不同灌木类型的分布区域; DSRTHC:高寒荒漠灌木半灌木, High-cold desert shrubland;DSRTTP, 温带荒漠灌木半灌木, Temperate desert shrubland;DCDSTP, 温带落叶灌木, Temperate deciduous shrubland;EVGNMT, 亚高山常绿灌木, Subalpine evergreen shrubland;DCDSMT, 亚高山落叶灌木, Subalpine deciduous shrubland;EVGNST, 亚热带常绿灌木, Subtropical evergreen shrubland |
本文利用多元线性回归模型对2001—2013年间年平均EVI与各季节SPEI进行了拟合, 并获得回归系数以分析季节SPEI变化对EVI的影响(表 4)。高寒荒漠灌木半灌木和温带荒漠灌木半灌木年平均EVI与夏季的SPEI均呈正相关, 而其夏季SPEI均呈增加趋势, 表明其所在区域呈湿润的趋势, 这将有利于植被的生长。温带落叶灌木的年EVI与其春季和夏季的SPEI显著正相关, 而与其秋季和冬季的SPEI显著负相关。亚高山常绿灌木EVI与季节SPEI的回归结果不能很好的解释EVI的变化, 即导致亚高山常绿灌木EVI的变化的原因很多, 全年的干旱化趋势可能仅是其部分原因, 也可能还存在着其他因素对EVI的变化产生了影响, 例如人类活动等。亚热带常绿灌木EVI与其夏季SPEI负相关而与秋季的SPEI正相关。亚热带常绿灌木夏季SPEI的下降主要是由于夏季气温升高和降水向秋季转移综合作用所导致的[19], 由于该区域年降水量充足, 水分并不是植被生长的限制因子, 此外, 降水量减小意味着云量会减少, 从而可以增加太阳辐射[23], 加上气温升高对植被生长的共同促进作用最终导致了EVI的增加。此外, 亚热带常绿灌木春季SPEI的下降趋势并没有对其EVI产生影响, 反而使其春季的EVI呈显著的增加趋势, 这是由于春季气温升高, 使植被生长加强所造成的。亚高山落叶灌木EVI与SPEI均未检测到显著的变化趋势, 也无显著的相关性。
| 灌木生态系统类型 Shrub ecosystem type |
截距 Intercept |
SPEI系数 SPEI coefficients/10-2 EVI·a-1·SPEI-1 | |||
| 春 Spring | 夏 Summer | 秋 Autumn | 冬 Winter | ||
| DSRTHC | 0.0711*** | 0.18* | |||
| DSRTTP | 0.0915*** | 0.54** | |||
| DCDSTP | 0.3299*** | 0.44* | 3.15*** | -0.83** | -0.63** |
| EVGNMT | 0.2743*** | -0.89** | |||
| DCDSMT | |||||
| EVGNST | 0.4143*** | -1.00** | 0.79* | ||
| ‘*’, ‘**’和‘***’分别表示P<0.1, 0.05和0.01 | |||||
本文还基于像元尺度计算了EVI与SPEI时间序列的相关性系数, 并进行了统计(表 5, 图 3)。EVI与SPEI呈显著负相关和不相关的面积比例非常小, 仅分别为1.33%和0.18%, 而呈不显著正相关的比例最大, 达到55.00%, 其大多分布于北方, 南方地区也有少量分布。显著正相关的比例为14.17%, 其主要分布在我国的北方(两种荒漠灌木以及温带落叶灌木)。不显著负相关的比例为29.32%, 其主要分布于我国的南方(两种亚高山灌木和亚热带常绿灌木)。此结果在一定程度上说明了北方的灌木生态系统比南方的灌木生态系统对干旱变化更加敏感。北方灌木分布区域年降水量较小, 通常受到水分胁迫, 因此其植被的生长对水分很敏感, 而南方的灌木区的降水量却很充足, 水分通常不是其限制因子, 适当的升温反而有利于植被的生长。
| Pearson相关系数 Pearson correlation coefficients |
相关性 Correlation |
面积百分比/% Area percentage |
| ≤-0.55 | 显著负相关 | 1.33 |
| -0.55 < r < 0 | 不显著负相关 | 29.32 |
| =0 | 不相关 | 0.18 |
| 0 < r < 0.55 | 不显著正相关 | 55.00 |
| ≥0.55 | 显著正相关 | 14.17 |
| r=±0.55对应5%的显著性水平 | ||

|
| 图 3 EVI与SPEI时间序列Pearson相关系数的空间分布 Fig. 3 Spatial distribution of Pearson correlation coefficients between EVI and SPEI r=±0.55对应5%的显著性水平 |
在中国的西南部地区其SPEI整体呈减小趋势, 其中四川与云南交界处以及云南的大部分地区检测到了SPEI显著减小的趋势(图 2), 这说明在2001—2013年间该地区呈显著的变干旱趋势, 这与其他人的研究结果相吻合[24-25]。西南地区整个SPEI的减小, 甚至云南省内大面积的SPEI显著减小并没有伴随着相应的EVI的显著下降(图 3), 且有研究表明干旱使中国西南地区的植被的NPP减小[24], 这说明南方湿润地区的灌木生态系统对干旱同样具有很强的抵抗作用。
3.2 中国灌木生态系统的SPEI趋势SPEI是通过计算水分平衡状态(降水-潜在蒸散)偏离多年平均值的程度来表征干旱的。SPEI < 0表示该地区水分平衡小于平均值, 该地区较平均状况偏干旱;SPEI>0表示该地区水分平衡大于平均值, 该地区较平均状况偏湿润。中国灌木生态系统SPEI的趋势自1961年以来在1992年出现了显著的改变(表 1), 从1992年之前的不显著上升趋势转变为显著下降趋势, 说明中国的灌木生态系统正在呈干旱化趋势, 这与我国的总体趋势相一致[26]。在6种不同的灌木类型中, 仅有温带落叶灌木的SPEI在2001—2013年间呈显著的增加的趋势(表 2), 其值从小于-1逐渐上升到大于0, 表明其由干旱状态回归到偏湿润状态。温带落叶灌木干旱胁迫的缓解将有利于植被的生长, 加之其生长季的延长[19], 将更加有利于其与夏季活跃型的温带草原之间的竞争, 使之更加有能力在未来的草灌竞争中占据上风。
干旱可以显著影响生态系统的碳循环, 可以通过影响植被的光合作用速率以及生态系统的呼吸来影响生态系统的NEE(Net Ecosystem Exchange, NEE), 并最终影响生态系统的碳循环[24]。此外, 干旱还能引发火灾[27]、造成树木死亡[28]以及引发虫害[29], 从而间接的影响生态系统的碳循环。全球植被的NPP在2001—2010年间因干旱呈显著下降的趋势[4], 中国的植被在干旱的影响下其NPP也显著下降, 甚至使全国的陆地生态系统从碳汇变成碳源[30]。灌木生态系统所表现出对干旱的抵抗性将在未来全球变暖、干旱加剧的背景下在区域乃至全球生态系统中将发挥重要的作用。
4 总结本文利用基于降水和蒸散水分平衡的干旱指数(SPEI)来模拟了中国灌木生态系统1961—2013年间的干旱状况。在1961—2013年间, 中国灌木生态系统在全国尺度呈显著的干旱趋势, 并且从不同灌木生态系统类型来看也全部呈显著的干旱化趋势, 这与目前全球气温升高, 干旱化越来越严重的趋势一致。本文还利用分段线性回归模型对SPEI时间序列进行了分析, 以判断其趋势是否有显著地改变。全国灌木生态系统以及两种荒漠类型灌木生态系统的SPEI均在1990年附近检测到了显著的转折点, 均从不显著的增加趋势转变为显著的下降趋势, 这与我国植被增长趋势在1990年代因干旱停滞相一致。在2001—2013年间, 中国灌木生态系统SPEI在全国尺度上呈下降趋势, 而对于不同灌木生态系统来说, 除温带落叶灌木的SPEI显著增加外其余5种灌木生态系统的SPEI均呈下降趋势(但是仅亚高山常绿灌木显著)。
本文还研究了不同灌木生态系统EVI对SPEI变化的响应, 结果显示, 南北灌木生态系统EVI对SPEI的变化影响不同, 北方灌木生态系统EVI对SPEI的变化更加敏感。其原因可能是南北降水量的显著差异所致, 在北方的干旱半干旱区域植物主要受到水分胁迫, 因而对水分变化敏感。西南地区SPEI在2001—2013年间显著下降, 但其EVI却并没有相应地减小, 表明南方湿润地区的灌木生态系统同样具有较强的抗干旱能力。在全球变暖, 干旱化越来越严重的将来, 灌木生态系统对区域乃至全球陆地生态系统的结构和功能必将起到越来越重要的作用。
| [1] |
Dai A G. Drought under global warming:a review. Climate Change, 2011, 2(1): 45-65. |
| [2] |
Ding Y H, Ren G Y, Zhao Z C, Xu Y, Luo Y, Li Q P, Zhang J. Detection, causes and projection of climate change over China:an overview of recent progress. Advances in Atmospheric Sciences, 2007, 24(6): 954-971. DOI:10.1007/s00376-007-0954-4 |
| [3] |
Hansen J, Ruedy R, Sato M, Lo K. Global surface temperature change. Reviews of Geophysics, 2010, 48(4): RG4004. |
| [4] |
Zhao M S, Running S W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science, 2010, 329(5994): 940-943. DOI:10.1126/science.1192666 |
| [5] |
Piao S L, Wang X H, Ciais P, Zhu B, Wang T, Liu J. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Global Change Biology, 2011, 17(10): 3228-3239. DOI:10.1111/gcb.2011.17.issue-10 |
| [6] |
Peng S S, Chen A P, Xu L, Cao C X, Fang J Y, Myneni, R B, Pinzon J E, Tucker C J, Piao S L. Recent change of vegetation growth trend in China. Environmental Research Letters, 2011, 6(4): 044027. DOI:10.1088/1748-9326/6/4/044027 |
| [7] |
Piao S L, Fang J Y, Zhou L M, Guo Q H, Henderson M, Ji W, Li Y, Tao S. Interannual variations of monthly and seasonal normalized difference vegetation index (NDVI) in China from 1982 to 1999. Journal of Geophysical Research, 2003, 108(D14): 4401. DOI:10.1029/2002JD002848 |
| [8] |
Gao Q, Zhao P, Zeng X, Cai X, Shen W. A model of stomatal conductance to quantify the relationship between leaf transpiration, microclimate and soil water stress. Plant, Cell & Environment, 2002, 25(11): 1373-1381. |
| [9] |
Gao Q, Yu M, Zhang X S, Xu H M, Huang Y M. Modelling seasonal and diurnal dynamics of stomatal conductance of plants in a semiarid environment. Functional Plant Biology, 2005, 32(7): 583-598. DOI:10.1071/FP04092 |
| [10] |
Gao Q, Yu M, Zhou C. Detecting the differences in responses of stomatal conductance to moisture stresses between deciduous shrubs and Artemisia subshrubs. PLoS One, 2013, 8(12): e84200. DOI:10.1371/journal.pone.0084200 |
| [11] |
谭永芹, 柏新富, 朱建军, 王仲礼, 刘林德. 干旱区五种木本植物枝叶水分状况与其抗旱性能. 生态学报, 2011, 31(22): 6815-6823. |
| [12] |
Smith S E, Fendenheim D M, Halbrook K. Epidermal conductance as a component of dehydration avoidance in Digitaria californica and Eragrostis lehmanniana, two perennial desert grasses. Journal of Arid Environments, 2006, 64(2): 238-250. DOI:10.1016/j.jaridenv.2005.04.012 |
| [13] |
中国科学院中国植被图编辑委员会. 中华人民共和国植被图 1:1000000. 北京: 地质出版社, 2007.
|
| [14] |
World Meteorological Organization. Standardized Precipitation Index User Guide. WMO-NO-1090. Geneva: WMO, 2012.
|
| [15] |
Vicente-Serrano S M, Beguería S, López-moreno J I. A multiscalar drought index sensitive to global warming:the standardized precipitation evapotranspiration index. Journal of Climate, 2010, 23(7): 1696-1718. DOI:10.1175/2009JCLI2909.1 |
| [16] |
周丹, 张勃, 任培贵, 张春玲, 杨尚武, 季定民. 基于标准化降水蒸散指数的陕西省近50a干旱特征分析. 自然资源学报, 2014, 29(4): 677-688. |
| [17] |
Zhao M S, Heinsch F A, Nemani R R, Running S W. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sensing of Environment, 2005, 95(2): 164-176. DOI:10.1016/j.rse.2004.12.011 |
| [18] |
Samanta A, Ganguly S, Hashimoto H, Devadiga S, Vermote E, Knyazikhin Y, Nemani R R, Myneni R B. Amazon forests did not green-up during the 2005 drought. Geophysical Research Letters, 2010, 37(5): L05401. |
| [19] |
Wang Y L, Gao Q, Liu T, Tian Y Q, Yu M. The greenness of major shrublands in China increased from 2001 to 2013. Remote Sensing, 2016, 8(2): 121. DOI:10.3390/rs8020121 |
| [20] |
Liang W, Yang Y T, Fan D M, Guan H D, Zhang T, Long D, Zhou Y, Bai D. Analysis of spatial and temporal patterns of net primary production and their climate controls in China from 1982 to 2010. Agricultural and Forest Meteorology, 2015, 204: 22-36. DOI:10.1016/j.agrformet.2015.01.015 |
| [21] |
Jiang W G, Yuan L H, Wang W J, Cao R, Zhang Y F, Shen W M. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecological Indicators, 2015, 51: 117-126. DOI:10.1016/j.ecolind.2014.07.031 |
| [22] |
王亚林, 龚容, 吴凤敏, 范文武. 2001-2013年中国灌木生态系统净初级生产力的时空变化特征及其对气候变化的响应. 植物生态学报, 2017, 41(9): 925-937. |
| [23] |
Piao S L, Fang J Y, Zhou L M, Ciais P, Zhu B. Variations in satellite-derived phenology in China's temperate vegetation. Global Change Biology, 2006, 12(4): 672-685. DOI:10.1111/gcb.2006.12.issue-4 |
| [24] |
Zhang L, Xiao J F, Li J, Wang K, Lei L P, Guo H D. The 2010 spring drought reduced primary productivity in southwestern China. Environmental Research Letters, 2012, 7(4): 045706. DOI:10.1088/1748-9326/7/4/045706 |
| [25] |
王东, 张勃, 安美玲, 张调风, 季定民, 任培贵. 基于SPEI的西南地区近53a干旱时空特征分析. 自然资源学报, 2014, 29(6): 1003-1016. |
| [26] |
栗健, 岳耀杰, 潘红梅, 叶信岳. 中国1961-2010年气象干旱的时空规律——基于SPEI和Intensity analysis方法的研究. 灾害学, 2014, 29(4): 176-182. DOI:10.3969/j.issn.1000-811X.2014.04.033 |
| [27] |
Xiao J F, Zhuang Q L. Drought effects on large fire activity in Canadian and Alaskan forests. Environmental Research Letters, 2007, 2(4): 044003. DOI:10.1088/1748-9326/2/4/044003 |
| [28] |
Allen C D, Macalady A K, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears D D, Hogg E H, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J L, Allard G, Running S W, Semerci A, Cobb N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 2010, 259(4): 660-684. DOI:10.1016/j.foreco.2009.09.001 |
| [29] |
Kurz W A, Dymond C C, Stinson G, Rampley G J, Neilson E T, Carroll A L, Ebata T, Safranyik L. Mountain pine beetle and forest carbon feedback to climate change. Nature, 2008, 452(7190): 987-990. DOI:10.1038/nature06777 |
| [30] |
Xiao J F, Zhuang Q L, Liang E Y, Shao X M, McGuire A D, Moody A, Kicklighter D W, Melillo J M. Twentieth-century droughts and their impacts on terrestrial carbon cycling in China. Earth Interactions, 2009, 13(10): 1-31. DOI:10.1175/2009EI275.1 |