 2019, Vol. 39
2019, Vol. 39文章信息
- 姚雪玲, 姜丽娜, 李龙, 王锋, 吴波, 郭秀江
- YAO Xueling, JIANG Lina, LI Long, WANG Feng, WU Bo, GUO Xiujiang
- 浑善达克沙地6种灌木生物量模拟
- Biomass simulation of six shrub species in Otindag sandy land
- 生态学报. 2019, 39(3): 905-912
- Acta Ecologica Sinica. 2019, 39(3): 905-912
- http://dx.doi.org/10.5846/stxb201803290637
-
文章历史
- 收稿日期: 2018-03-29
- 网络出版日期: 2018-11-15





2. 中国林业科学研究院林业新技术研究所, 北京 100091;
3. 内蒙古农业大学沙漠治理学院, 呼和浩特 010018;
4. 华北油田技术监督检验处计量中心站, 任丘 062552
2. Research Institute of Forestry New Technology, Chinese Academy of Forestry, Beijing 100091, China;
3. College Of Desert Control Science, Inner Mongolia Agricultural University, Hohhot 010018, China;
4. Technical Supervision Department of Huabei Oilfield, Metrology Center Station, Renqiu 062552, China
灌木林是自然界中广泛存在的一种重要的陆地生态系统类型。长期以来, 灌木林因与乔木林相比所占比例较小处于次要地位而较少受到重视, 关于灌木林生物量的研究国内外报道也相对较少[1]。灌木在森林生态系统中虽然占全部生物量的很小部分, 但是在荒漠、高原、干热河谷、荒山荒地和石漠化等生态脆弱地区, 灌木仍然占有很大比重, 对生态系统的稳定与平衡具有十分重要的意义[2]。在我国干旱、半干旱地区有大量灌木分布, 灌木兼有防风固沙、保持水土、改良盐碱地等多种生态功能。研究灌木生物量是植被生态学研究的一个重要内容, 是衡量植被生产力的重要指标, 它对于研究灌木生长发育规律、灌木在生态系统中的作用和地位等都具有重要意义[3-4]。
常用的植物生物量测定方法主要有3种:一种是直接收获法, 刈割地表植株后称重;第二种是样方法, 用单位面积的生物量推算整体生物量;第三种是数学模型法, 通过建立生物量与易测因子(树高、地径等)的相关方程, 来推算单株树木、单位面积部分或整体的生物量[5]。在干旱半干旱区, 植被稀少, 直接刈割采伐灌木会对生态环境造成破坏, 因而不适宜大规模采用[6]。而灌木在水平分布上又具有明显的不连续性和不均匀性, 不适合像草本那样用样方收获法测定其生物量。较为适宜的方法是数学模型法。目前预测灌木生物量的数学模型主要有线性模型、多项式模型、幂指数异速生长模型等[1]。常用的预测因子主要包括地径、高度、冠幅、盖度、年龄、分枝数(株数)等, 以及以此为基础构建的植冠面积、植冠体积、地径与高度乘积、地径平方与高度乘积等复合因子, 在文献中已有大量应用[7-13]。目前的灌木生物量预测方法大都来源于乔木生物量预测方法, 专门针对灌木的生物量预测方法研究尚不充分。而灌木具有其特殊的个体形态结构, 如灌木较低矮, 多是丛生, 主杆不明显等[5]。针对灌木本身的形态特征, 探讨特别适用于灌木的生物量预测方法, 提高灌木生物量预测精度, 将对灌木的生物量以及碳汇贡献的研究具有积极意义。
本研究以浑善达克沙地榆树疏林6种主要灌木为研究案例, 采用几种常用的灌木生物量预测方法, 同时设计了一种圆台体积-生物量预测方法, 通过对这些方法对生物量的预测能力的评估, 来探讨更适用于灌木的生物量预测方法。研究最终给出了6种沙地灌木的最优生物量预测模型。
1 研究方法 1.1 研究区概况研究样地(面积1 km2)位于浑善达克沙地中部的典型榆树疏林分布区, 位于锡林郭勒盟正蓝旗桑根达来镇东北30 km处(图 1)。该地区平均海拔1300 m, 受温带半干旱气候控制, 年均温度0.5—3.5℃, 年均降水量250—400 mm, 年均蒸发量2000—2700 mm, 全年盛行西风, 年均风速3.5—5.5 m/s, 主要土壤类型为栗钙型风沙土。榆树疏林是浑善达克沙地和科尔沁沙地中的顶级植被群落, 也是沙地分布最广泛的生态系统之一, 在京津冀风沙源区起到重要的防风固沙作用[14]。榆树疏林景观特征为榆树乔木稀疏散布于草地之中, 沙甸和沙丘背风坡分布有密集灌丛。其中灌木丛分布面积大, 植株密集, 是地表植被生物量的重要组成部分。灌木林因其覆盖度大, 树冠靠近地表, 生长密集, 物种多样性较高, 在防护地表沙化、改良土壤、为野生动物提供栖息地等方面发挥着重要的生态功能。对于榆树疏林生态系统中榆树生物量的研究已有不少报道[15]。而对于其中灌木生物量的研究较为缺乏[16]。

|
| 图 1 研究区位置示意图 Fig. 1 Schematic of the study location |
2015年8月, 对浑善达克沙地疏林草地的六种主要灌木类型, 包括楔叶茶藨子(Ribes diacanthum Pall.)、柴桦(Betula fruticosa)、小叶锦鸡儿(Caragana microphylla)、小红柳(Salix microstachya)、小黄柳(Salix flavida)、耧斗叶绣线菊(Spiraea aquilegifolia), 每种灌木根据其高度和冠幅等级, 选择不同大小的20株代表木, 测定每株灌木的长地径、短地径、长冠幅、短冠幅、高度等属性。按一定比例采用地面爻割法, 测定其地表鲜生物量, 取一定比例的枝条, 实验室烘干后测得水分含量, 通过比例换算得出整个植株的干生物量。
1.3 预测指标的选择本研究选择应用最为广泛的高度(h)、长冠幅(a1)、短冠幅(b1)、长地径(a2)、短地径(b2)5项测量因子(图 2)。通过这5个单因子计算得出植冠体积(AH)、地径平方与高度乘积(D2H)两个复合指标, 计算公式如下:
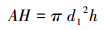
|
(1) |

|
| 图 2 灌木属性测量指标示意图 Fig. 2 Schematic diagram of the measurements of the shrub a1:最大冠幅, maximum crown diameter;b1:最大冠幅的垂直交叉冠幅, the perpendicular diameter of a1;a2, 最大地径, maximum diameter at the base;b2:最大地径的垂直交叉地径, the perpendicular diameter of b1;h:最大高度, the distance between the ground surface and highest crown point |
式中, d1是冠幅的半径, 既长半径和短半径1/2a1、1/2b1的平均值。h是植株高度。
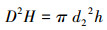
|
(2) |
式中, d2是基径的半径, 既长半径和短半径1/2a2、1/2b2的平均值。h是植株高度。
另外, 根据灌木的形态特征, 设计了倒圆台体积模型(V)作为新的预测指标。圆台体积模型公式如下:
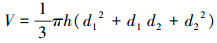
|
(3) |
式中, d1、d2和h含义同上。
以上述5个单因子和3个复合因子作为初始预测指标, 进一步通过相关分析和拟合分析, 评选出最优预测指标, 并采用最优指标对沙地6种灌木的生物量进行建模。
1.4 灌木生物量模型构建及精度评价相对生长模型(也称异速生长方程)是应用最为普遍的一种生物量估计模型。大量研究表明, 相对生长模型相比其他模型, 如线性模型、多项式模型等, 其外推性和稳定性更好, 因此本研究选用该模型对灌木生物量进行拟合[1, 17]。相对生长模型的结构式表达如下[18]:
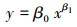
|
(4) |
式中y为灌木总生物量。x为代表灌木大小的变量, 即预测指标。β0、β1为模型参数[1]。
模型拟合效果采用校正决定系数Adj.R2、显著性指标p值、估计值的标准误SEE指标进行评价。这些评价指标的公式及含义在很多文献中都有介绍, 这里不再赘述[3, 10]。
2 研究结果 2.1 预测指标与灌木生物量的相关性评价对6种灌木生物量及各个预测指标进行相关分析, 相关系数及其显著性能初步反映各指标对生物量的预测能力[19]。结果显示, 灌木高度和地径与生物量之间的相关性较弱, 相关系数小于0.7。对于一些物种, 如锦鸡儿、绣线菊、柴桦等, 其相关性达不到显著水平(表 1)。冠幅与生物量相关系数在0.7—0.8之间, 相关性明显优于地径。复合因子能明显改善单因子与生物量之间的相关性。基于地径和高度的复合因子D2H与生物量之间的相关性均值在0.76, 明显优于高度和地径单因子。基于冠幅和高度的植冠体积与生物量的相关系数在0.84, 也明显优于冠幅单因子。预示着复合因子对灌木生物量的预测能力更好, 这与已有研究结论一致[20-21]。圆台体积能复合更多的测量指标, 其与生物量之间的相关性为0.88, 在所有指标中表现最好。综合考虑各预测指标与生物量之间的相关性, 选择圆台体积、植冠体积、D2H三个指标进一步进行拟合评价。
| 物种 Shrub species |
高度 Height |
长冠幅 Long crown diameter |
短冠幅 Short crown diameter |
长地径 Long base diameter |
短地径 Short base diameter |
D2H | 植冠体积 Crown volume |
圆台体积 Round table volume |
| 楔叶茶彪子Ribes diacanthum Pall. | 0.54** | 0.85** | 0.79** | 0.80** | 0.80** | 0.80** | 0.88** | 0.91** |
| 柴桦Betula fruticosa | 0.48** | 0.68** | 0.65** | 0.65** | 0.17 | 0.63** | 0.70** | 0.79** |
| 小叶锦鸡儿Caragana microphylla | 0.29 | 0.73** | 0.68** | 0.69** | 0.48 | 0.72** | 0.77** | 0.87** |
| 小红柳Salix microstachya | 0.74** | 0.75** | 0.89** | 0.77** | 0.70** | 0.88** | 0.91** | 0.95** |
| 小黄柳Salix flavida | 0.61** | 0.84** | 0.89** | 0.71** | 0.82** | 0.85** | 0.97** | 0.93** |
| 耧斗叶绣线菊Spiraea aquilegifolia. | 0.51* | 0.79** | 0.78** | 0.49* | 0.59** | 0.69** | 0.82** | 0.84** |
| 平均值Average | 0.53 | 0.77 | 0.78 | 0.69 | 0.59 | 0.76 | 0.84 | 0.88 |
| D2H:地径平方与高度的乘积, the product of square base diameter and height; **表示达到极显著相关P < 0.001;*表示达到显著相关P < 0.05 | ||||||||
为进一步评价圆台体积、植冠体积、D2H的3个指标对灌木生物量的预测能力, 建立3个指标与生物量之间的异速生长模型, 对模型的拟合效果采用校正决定系数Adj.R2和估计值标准误SEE进行评价。模型的结构见公式4, 模型参数以及评价指标值见表 2。为了使模型之间具有可比性, 将原始数据进行标准化, 绘制生物量和各个预测指标之间的异速生长方程拟合线(图 3), 并对拟合方程的Adj.R2和SEE值绘制箱图进行对比(图 4)。结果显示, 圆台体积-生物量异速生长模型的Adj.R2均值(0.74)较高, SEE值(0.016)较小, 说明相比植冠体积、D2H, 圆台体积对生物量具有更好的拟合精度。其中圆台体积模型和植冠体积模型之间的Adj.R2均值相差较小, 然而植冠体积模型的Adj.R2值和SEE值在不同物种之间波动较大(表 2), 在箱图中表现为变异范围较大(图 4), 说明其对不同灌木种类进行生物量预测时稳定性较差。综合来看, 植冠体积对生物量的预测效果优于D2H, 而圆台体积对生物量的预测精度以及稳定性方面均优于前两者指标。

|
| 图 3 灌木生物量和各预测指标的异速生长模型拟合曲线 Fig. 3 Fitting curve of allometry model for biomass and various predictive indexes of the six shrub species |

|
| 图 4 基于Adj.R2和SEE的灌木生物量模型拟合精度评价 Fig. 4 Evaluation of fitting precision of shrub biomass model based on Adj.R2 and SEE |
| 物种 Shrub species |
植冠体积 Crown volume |
圆台体积 Round table volume |
D2H | |||||||||||
| β0 | β1 | Adj.R2 | SEE | β0 | β1 | Adj.R2 | SEE | β0 | β1 | Adj.R2 | SEE | |||
| 楔叶茶彪子 Ribes diacanthum Pall. |
0.827 | 1.483 | 0.796 | 0.013 | 0.855 | 1.272 | 0.844 | 0.010 | 0.801 | 0.598 | 0.724 | 0.018 | ||
| 柴桦 Betula fruticosa |
0.792 | 0.879 | 0.562 | 0.023 | 0.848 | 1.027 | 0.652 | 0.018 | 0.723 | 0.773 | 0.511 | 0.026 | ||
| 小叶锦鸡儿 Caragana microphylla |
1.185 | 1.388 | 0.478 | 0.032 | 1.069 | 2.249 | 0.819 | 0.011 | 0.973 | 1.338 | 0.730 | 0.016 | ||
| 小红柳 Salix microstachya |
1.063 | 1.108 | 0.838 | 0.015 | 0.753 | 1.057 | 0.745 | 0.017 | 1.021 | 0.876 | 0.754 | 0.024 | ||
| 小黄柳 Salix flavida |
1.004 | 0.954 | 0.843 | 0.011 | 0.892 | 0.939 | 0.795 | 0.015 | 0.792 | 0.666 | 0.569 | 0.031 | ||
| 耧斗叶绣线菊 Spiraea aquilegifolia. |
0.950 | 0.572 | 0.600 | 0.027 | 1.029 | 0.639 | 0.612 | 0.026 | 0.840 | 0.564 | 0.332 | 0.045 | ||
| 平均值Average | 0.686 | 0.020 | 0.744 | 0.016 | 0.603 | 0.027 | ||||||||
| β0:异速生长模型参数, parameter of the allometric model; β1:异速生长模型参数, parameter of the allometric model; Adj.R2:校正决定系数, adjusted coefficient of determination; SEE:估计值的标准误, standard deviation; 异速生长模型公式为:y=β0xβ1, Adj.R2和SEE表征异速生长模型的模拟精度 | ||||||||||||||
基于以上各预测指标的对比分析, 以圆台体积作为灌木的预测指标更为理想。对6种灌木的干生物量的实测值和圆台体积进行异速生长方程拟合, 模型参数及拟合结果见图 5, 模型均通过显著性检验(P < 0.05)。

|
| 图 5 6种灌木的圆台体积-生物量的异速生长模型 Fig. 5 Allometric growth model of the round table volumetric-biomass of the six shrub species |
本研究对几种常见的生物量预测指标进行了对比分析, 探讨了其对灌木生物量的预测能力。其中, 特别设计了一种更贴合灌木形态特征的圆台体积作为新的预测指标。研究结果表明, 单因素测量指标中, 相比高度和地径, 冠幅与灌木生物量的之间的相关性更好, 预示着冠幅对生物量的预测能力较高, 很多研究中也有类似结论[6, 21-22]。圆台体积、植冠体积和D2H等复合指标与生物量的相关性比单因素指标更强, 预示着复合指标能进一步提高灌木生物量预测能力, 这也与已有研究结果一致[20]。通过进一步的拟合精度评价, 圆台体积相较于植冠体积和D2H, 其拟合度更好, 误差较小。对于不同的灌木种类, 如小红柳、柴桦、小叶锦鸡儿等, 其植冠形态和个体大小存在明显差异, 然而其圆台体积与生物量之间均能保持较高的相关性, 并且在拟合分析中, 拟合误差变异较小。这意味着圆台体积对不同灌木种类的预测能力会较为稳定。另外, 因圆台体积综合了包括高度、冠幅、地径等较多的测量属性在内, 在野外测量较为充分的情况下, 圆台体积是理想的生物量预测指标, 而在野外测量指标不充分的情况下, 冠幅或者基于冠幅的复合变量更适宜用来预测灌木生物量。
| [1] |
曾伟生. 国内外灌木生物量模型研究综述. 世界林业研究, 2015, 28(1): 31-36. |
| [2] |
Zandler H, Brenning A, Samimi C. Quantifying dwarf shrub biomass in an arid environment:comparing empirical methods in a high dimensional setting. Remote Sensing of Environment, 2015, 158: 140-155. DOI:10.1016/j.rse.2014.11.007 |
| [3] |
李钢铁, 秦富仓, 贾守义, 董锦兰, 吉日格勒, 王满贵. 旱生灌木生物量预测模型的研究. 内蒙古林学院学报, 1998, 20(2): 26-32. |
| [4] |
Conti G, Enrico L, Casanoves F, Díaz S. Shrub biomass estimation in the semiarid Chaco forest:a contribution to the quantification of an underrated carbon stock. Annals of Forest Science, 2013, 70(5): 515-524. DOI:10.1007/s13595-013-0285-9 |
| [5] |
卢振龙, 龚孝生. 灌木生物量测定的研究进展. 林业调查规划, 2009, 34(4): 37-40, 45-45. DOI:10.3969/j.issn.1671-3168.2009.04.011 |
| [6] |
Yang H T, Wang Z R, Tan H J, Gao Y H. Allometric models for estimating shrub biomass in desert grassland in northern China. Arid Land Research and Management, 2017, 31(3): 283-300. DOI:10.1080/15324982.2017.1301595 |
| [7] |
万五星, 王效科, 李东义, 冯学全, 张千千, 于小红. 暖温带森林生态系统林下灌木生物量相对生长模型. 生态学报, 2014, 34(23): 6985-6992. |
| [8] |
林伟, 李俊生, 郑博福, 郭建明, 胡理乐. 井冈山自然保护区12种常见灌木生物量的估测模型. 武汉植物学研究, 2010, 28(6): 725-729. |
| [9] |
赵蓓, 郭泉水, 牛树奎, 孙武, 张志旭. 大岗山林区几种常见灌木生物量估算与分析. 东北林业大学学报, 2012, 40(9): 28-33. DOI:10.3969/j.issn.1000-5382.2012.09.008 |
| [10] |
何列艳, 亢新刚, 范小莉, 高延, 冯启祥. 长白山区林下主要灌木生物量估算与分析. 南京林业大学学报:自然科学版, 2011, 35(5): 45-50. |
| [11] |
仇瑶, 常顺利, 张毓涛, 王文栋, 何平, 王慧杰, 谢锦. 天山林区六种灌木生物量的建模及其器官分配的适应性. 生态学报, 2015, 35(23): 7842-7851. |
| [12] |
罗永开, 方精云, 胡会峰. 山西芦芽山14种常见灌木生物量模型及生物量分配. 植物生态学报, 2017, 41(1): 115-125. |
| [13] |
Ludwig J A, Reynolds J F, Whitson P D. Size-biomass relationships of several Chihuahuan desert shrubs. The American Midland Naturalist, 1975, 94(2): 451-461. DOI:10.2307/2424437 |
| [14] |
史宇飞, 金永焕, 金兰淑. 国内榆树疏林研究现状. 水土保持应用技术, 2011(2): 32-35. DOI:10.3969/j.issn.1673-5366.2011.02.12 |
| [15] |
赵云杰, 田赟, 王晓. 典型榆树疏林碳氮储量及其分配特征. 中国水土保持科学, 2014, 12(6): 82-89. DOI:10.3969/j.issn.1672-3007.2014.06.012 |
| [16] |
马媛, 李钢铁, 潘羿壅, 麻云霞, 王强, 张驰. 浑善达克沙地3种灌木生物量的预测模型. 干旱区资源与环境, 2017, 31(6): 198-201. |
| [17] |
Rojo V, Arzamendia Y, Pérez C, Baldo J, Vilá B. Double sampling methods in biomass estimates of Andean shrubs and tussocks. Rangeland Ecology & Management, 2017, 70(6): 718-722. |
| [18] |
罗红, 李百炼. 异速生长模型研究概述. 生态学杂志, 2011, 30(9): 2060-2065. |
| [19] |
Vora R S. Predicting biomass of five shrub species in northeastern California. Journal of Range Management, 1988, 41(1): 63-65. DOI:10.2307/3898792 |
| [20] |
曾珍英, 刘琪璟, 曾慧卿. 江西千烟洲几种灌木生物量模型的研究. 福建林业科技, 2005, 32(4): 68-72. DOI:10.3969/j.issn.1002-7351.2005.04.019 |
| [21] |
Huff S, Ritchie M, Temesgen H. Allometric equations for estimating aboveground biomass for common shrubs in northeastern California. Forest Ecology and Management, 2017, 398: 48-63. DOI:10.1016/j.foreco.2017.04.027 |
| [22] |
Zhang L, Cui G S, Shen W, Liu X S. Cover as a simple predictor of biomass for two shrubs in Tibet. Ecological Indicators, 2016, 64: 266-271. DOI:10.1016/j.ecolind.2016.01.009 |