 2019, Vol. 39
2019, Vol. 39文章信息
- 牛玉斌, 余海龙, 王攀, 樊瑾, 王艳红, 黄菊莹
- NIU Yubin, YU Hailong, WANG Pan, FAN Jin, WANG Yanhong, HUANG Juying
- 氮磷添加对荒漠草原植物群落多样性和土壤C:N:P生态化学计量特征的影响
- Effects of nitrogen and phosphorus addition on plant community diversity and soil C: N: P ecological stoichiometry in a desert steppe of China
- 生态学报. 2019, 39(22): 8462-8471
- Acta Ecologica Sinica. 2019, 39(22): 8462-8471
- http://dx.doi.org/10.5846/stxb201812022624
-
文章历史
- 收稿日期: 2018-12-02
- 网络出版日期: 2019-09-05





2. 浙江农林大学省部共建亚热带森林培育国家重点实验室, 杭州 311300;
3. 宁夏大学环境工程研究院, 银川 750021
2. State Key Laboratory of Subtropical Silviculture, Zhejiang A & F University, Hangzhou 311300, China;
3. Institute of Environmental Engineering, Ningxia University, Yinchuan 750021, China
自工业革命以来, 化石燃料燃烧和农牧业集约化发展等人类活动导致全球大气活性氮(Nr)排放快速上升[1]。近期的研究指出, 2010年我国N沉降总量约为7.6×1012 g, 已成为继美国和欧洲以来的第三大N沉降区[2-3]。空间分布上, 我国N沉降总量具有东高西低、沿海高于内陆的区域分异特征。就西北地区而言, 2000—2010年宁夏大部分区域N沉降速率达到2.0 g m-2 a-1, 部分区域N沉降速率甚至超过4.0 g m-2 a-1, 高于西北地区的平均水平[4]。长期N沉降增加加速了土壤N循环, 引起土壤碳(C):N:磷(P)比失衡和植物P限制增强, 进一步导致植物种丧失和生态系统退化[5]。P是细胞间能量传递和核苷酸结构组成的重要元素, 在植物生长发育的多个生理生态活动中扮演着重要角色。同时, P又是多数陆地生态系统植物生长的主要限制因子之一[6]。施P肥提高了土壤P有效性, 因此可能会缓解N沉降增加引起的植物群落多样性降低。然而, 目前针对N添加下增施P肥是否会有助于提高植物群落多样性的研究还较为缺乏, 尤其是针对宁夏荒漠草原开展的相关研究更是少见。
生态化学计量学是研究生态系统多种化学元素平衡关系的一个新兴生态学研究领域, 为研究土壤-植物相互作用的养分平衡制约关系提供了新思路[7]。土壤-植被系统C:N:P比可以指示系统C饱和状态和养分受限类型[8-9]。其中, 土壤C:N和C:P比的高低, 决定着微生物活动的变化趋势(释放或固持)。微生物量C:N:P比则决定了凋落物分解过程中养分的释放与否, 进而影响着植物获得有效养分的情况。因此, C、N、P在土壤和微生物之间传递和转化的耦合关系, 对于维持植物群落结构稳定性具有重要意义[10]。然而, 近年来随着N沉降的急剧增加, 土壤和微生物量C:N:P计量平衡关系趋于解耦, 导致生态系统服务功能降低[11-13]。土壤-植被系统N:P比失衡已被认为是N沉降增加引起植物种丧失的重要机制之一[14]。目前, 已有较多研究将植物C:N:P平衡关系应用于解释N沉降增加引起的物种丧失机理[5, 15], 但从土壤和微生物量C:N:P生态化学计量特征角度的探讨还比较缺乏。
宁夏荒漠草原是我国典型的生态脆弱区。近几十年来全球变化的不断增强和人类活动的持续干扰, 进一步加剧了区域植被的退化。尽管模型估测的N沉降量低于我国南方等地区, 但宁夏荒漠草原对N沉降比较敏感[16], 且即使是低剂量的N沉降, 其长期影响也是不容忽视的[17]。因此, 有必要在宁夏荒漠草原开展N沉降效应的研究。鉴于此, 2011年项目组在宁夏荒漠草原设置了模拟N沉降增加的野外试验。结果表明, 低于5—10 g m-2 a-1的N添加促进了植物生长, 随着N添加量的增加植物种数有减少趋势[18]。那么, 少量N添加下施P肥是否会缓解N沉降增加引起的植物群落多样性降低?为了回答这一问题, 项目组于2015年设置了N和P添加的原位试验, 研究了少量N添加下施用P肥对宁夏荒漠草原植物生物量和群落多样性的影响, 分析了引起植被群落组成发生改变的关键土壤因子, 研究结果可为充分认识全球变化下地上群落结构和地下生态过程的内在联系提供新思路。
1 材料与方法 1.1 研究区概况N和P添加试验设立于1998年开始围封的宁夏盐池县柳杨堡乡杨寨子村草地内。该草地(北纬37.80°, 东经107.45°, 海拔约1367 m)地处毛乌素沙地西南边缘, 是黄土高原向鄂尔多斯台地的过渡地带。试验区属于中温带大陆性气候, 年平均气温为7.7℃, 其中, 1月平均气温为-8.9℃, 7月平均气温为22.5℃; 年均蒸发量为2131.8 mm, 年均降水量为289.4 mm, 且75%以上的降水集中在植物生长季(5—9月份)[19]。试验区内主要土壤类型为灰钙土, 主要土壤质地为沙壤土; 主要植被类型为荒漠草原, 植物群落结构简单, 多为连续片状分布, 并且年际变化大。自退耕还林还草政策的实施以来, 与未围封草地相比围栏内草地植被恢复良好, 物种组成以一年生和多年生草本为主, 如猪毛蒿(Artemisia scoparia)、猪毛菜(Salsola collina)、牛枝子(Lespedeza potaninii)、草木樨状黄芪(Astragalus melilotoides)、苦豆子(Sophora alopecuroides)、中亚白草(Pennisetum centrasiaticum)、糙隐子草(Cleistogenes squarrosa)、冰草(Agropyron cristatum)和针茅(Stipa capillata)等。
1.2 试验设计2015年1月, 在围栏草地内选择地势平坦、植被均匀有代表性的地段作为N和P添加处理的长期试验样地。N肥施用量以2011—2013年于宁夏盐池县四佟子乡围栏草地设立的N添加原位试验的观测结果为主要依据[18], 同时参考了区域大气N沉降水平[4]。P肥施用量以2011—2013年设立的N:P供给盆栽试验的观察结果为主要依据[20], 同时参考了研究区土壤P供给水平。2015年4月底在选定的试验样地, 按照随机区组试验设计, 设置了7个水平的N和P添加处理:在统一施用5 g m-2 a-1纯N的基础上, 通过添加不同量的P肥(0、0.5、1.0、2.0、4.0、8.0和16.0 g m-2 a-1)形成7个N和P添加处理, 分别为N5P0、N5P0.5、N5P1、N5P2、N5P4、N5P8和N5P16。每个试验小区面积为8 m×8 m, 小区之间设置了1.5 m宽的缓冲带, 每个处理5次重复。所施N肥为含34%纯N的NH4NO3 (分析纯), 所施P肥为含51.7% P2O5的KH2PO4 (分析纯)。施用时, 将每个样方年施用量平分成4等分, 于5—8月每月月初选择阴天或者雨后傍晚将NH4NO3和KH2PO4溶于少量体积的水中, 用撒壶在该水平样方中来回均匀喷洒。同时, 为避免因水分施用量不同而造成的试验误差, 在对照样地(N10P0处理)喷洒同体积的清水。
2016年8月中旬, 在每个小区内随机选择3个1 m×1 m的小样方进行植被群落调查。调查内容包括植物种组成、物种数、高度和频度等。植被调查结束后, 齐平地面剪下植物地上部分, 按物种分装入牛皮纸袋带回实验室烘干(65℃, 48h)并称重。本试验选择目前常用的四个指数:Patrick丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数对植物群落多样性进行评价。各指标计算公式如下[21]:
重要值: Pi=(相对生物量+相对高度+相对频度)/3
Patrick丰富度指数: R=S
Shannon-Wiener多样性指数: 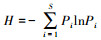
Pielou均匀度指数: E=H/lnS
Simpson优势度指数: 
式中, S为群落中物种数, Pi为种i的相对重要值。
同期, 用内径为5 cm的土钻采取每个小区0—20 cm的土壤样品, 每个小区取3钻, 混匀后带回实验室。在实验室, 将混匀后的土样过2 mm筛后分为两部分:一部分自然风干, 用于有机C含量、全N含量、全P含量和速效P浓度的测定[22]; 另一部分4℃下冷藏保存, 用于pH、NH4+-N浓度、NO3--N浓度和微生物量的测定。其中, 有机C含量采用重铬酸钾容量-外加热法测定; 全N含量采用凯氏定氮法测定; 全P含量采用HClO4-H2SO4法测定; 速效P浓度采用0.5 mol/L NaHCO3法测定; CaSO4·2H2O溶液中浸提后, 采用连续流动分析仪(Auto Analyzer 3, SEAL Analytical GmbH, Hanau, Germany)分析新鲜土样NH4+-N和NO3--N浓度; pH采用便携式酸度计测定; 微生物量C、N和P含量采用改进的氯仿熏蒸法测定。称取过2 mm筛的新鲜土样2份, 一份进行氯仿熏蒸, 一份不进行熏蒸。熏蒸与未熏蒸的土壤样品用0.5 mol/L K2SO4溶液振荡浸提30 min, 浸提液用中速定量滤纸过滤, 并通过0.45 μm微孔滤膜, 用总有机碳分析仪测定微生物量C和N含量。两份土样经0.5 mol/L NaHCO3溶液浸提、振荡、过滤后, 消化钼蓝比色法测定微生物量P含量。
1.3 数据处理与分析原始数据的整理分析在Excel 2003中完成, 图的绘制在Origin 9.0中完成, 数据的统计学分析在SPSS 19.0和Canoco 4.5中完成。数据分析前, 进行了植物指标的方差齐性检验。通过方差齐性检验的指标, 采用One-Way ANOVA进行单因素方差分析, 采用最小显著性差异法(LSD)进行处理间的多重比较。未通过齐性检验的指标, 利用非参数检验(Mann-Whitney)进行处理间的显著性分析。对植物群落多样性指数和土壤因子的对应关系进行冗余分析(RDA)。由于各植物群落多样性指数之间差异较大, RDA排序过程中对数据进行了转换, 转换公式为Y′=log(Y+1)。图中数据为平均值+标准误(n=5),表中数据为平均值±标准误(n=5)。
2 结果与分析 2.1 N和P添加对植物生物量和群落多样性的影响5 g m-2 a-1的N添加下, 植物群落地上生物量随P添加量增加呈现出先略有增加后降低的趋势(图 1); 随着P添加量增加, 牛枝子、草木樨状黄芪和苦豆子种群生物量亦呈现出先增加后降低的趋势, 猪毛蒿种群生物量持续增加, 猪毛菜种群生物量持续降低(图 2), 说明增施P肥促进了多数植物生物量积累, 但中高量P肥抑制了牛枝子、草木樨状黄芪和苦豆子的生长。

|
| 图 1 N和P添加对植物群落生物量的影响 Fig. 1 Effects of N and P addition on plant community biomass N5P0、N5P0.5、N5P1、N5P2、N5P4、N5P8和N5P16代表在统一施用5.0 g N/m2/a的基础上, 分别施入0.0、0.5、1.0、2.0、4.0、8.0和16.0 g m-2 a-1的P; 不同小写字母表示群落生物量在N和P添加处理间差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) |

|
| 图 2 N和P添加对植物种群生物量的影响 Fig. 2 Effects of N and P addition on population biomass of the studied species N5P0、N5P0.5、N5P1、N5P2、N5P4、N5P8和N5P16代表在统一施用5.0 g N/m2/a的基础上, 分别施入0.0、0.5、1.0、2.0、4.0、8.0和16.0 g m-2 a-1的P; 植被群落调查时, N5P0和N5P16处理未发现草木樨状黄芪, N5P16处理未发现苦豆子, N5P1处理未发现其他物种, 故相应处理上无数据; 不同小写字母表示种群生物量在N和P添加处理间差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) |
5 g m-2 a-1的N添加下, 少量P添加对牛枝子、苦豆子、猪毛蒿和猪毛菜重要值影响较小; 中高量P添加降低了牛枝子、苦豆子、猪毛菜和其他物种重要值, 提高了猪毛蒿重要值。此外, N5P16处理下草木樨状黄芪和苦豆子重要值为0, 表明高量P添加导致两个物种从群落中逐渐被淘汰(表 1)。
| 处理 Treatments |
牛枝子 Lespedeza potaninii |
草木樨状黄芪 Astragalus melilotoides |
苦豆子 Sophora alopecuroides |
猪毛蒿 Artemisia scoparia |
猪毛菜 Salsola collina |
其他物种 Other species |
| N5P0 | 0.19±0.01 a | nd | 0.14±0.03 a | 0.18±0.02 c | 0.45±0.04 a | 0.04±0.03 a |
| N5P0.5 | 0.19±0.01 a | 0.02±0.02 b | 0.15±0.04 a | 0.21±0.01 c | 0.43±0.03 a | 0.01±0.01 a |
| N5P1 | 0.18±0.01 a | 0.04±0.02 b | 0.14±0.06 a | 0.23±0.01 c | 0.41±0.03 a | nd |
| N5P2 | 0.18±0.02 a | 0.15±0.02 a | 0.09±0.04 ab | 0.26±0.07 c | 0.29±0.09 ab | 0.04±0.03 a |
| N5P4 | 0.19±0.03 a | 0.12±0.01 a | 0.06±0.04 ab | 0.42±0.03 b | 0.15±0.07 b | 0.06±0.03 a |
| N5P8 | 0.13±0.04 ab | 0.10±0.03 a | 0.02±0.02 b | 0.48±0.04 ab | 0.22±0.10 b | 0.05±0.03 a |
| N5P16 | 0.09±0.04 b | nd | nd | 0.55±0.07 a | 0.27±0.03 ab | 0.09±0.06 a |
| N5P0、N5P0.5、N5P1、N5P2、N5P4、N5P8和N5P16代表在统一施用5.0 g N/m2/a的基础上, 分别施入0.0、0.5、1.0、2.0、4.0、8.0和16.0 g m-2 a-1的P; nd代表该处理下无相应物种; 不同小写字母表示重要值在N和P添加处理间差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) | ||||||
5 g m-2 a-1的N添加下, 增施P肥改变了植物群落多样性(图 3):随着P添加量增加Shannon-Wiener多样性指数和Patrick丰富度指数呈现出先增加后降低的趋势, 二者均在2 g m-2 a-1的P添加水平下达到最大值; 沿着P添加量增加梯度, Simpson优势度指数呈现出先略有降低后持续增加的趋势。总体而言, 中少量P添加对Simpson优势度指数的影响未达到显著水平; 与N5P0处理相比较, 除N5P4处理以外其他几个N和P添加处理显著降低了Pielou均匀度指数。

|
| 图 3 N和P添加对植物群落多样性指数的影响 Fig. 3 Effects of N and P addition on plant community diversity indices N5P0、N5P0.5、N5P1、N5P2、N5P4、N5P8和N5P16代表在统一施用5.0 g N/m2/a的基础上, 分别施入0.0、0.5、1.0、2.0、4.0、8.0和16.0 g m-2 a-1的P; 不同小写字母表示同一指标在N和P添加处理间差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) |
5 g m-2 a-1的N添加下, 增施P肥改变了土壤C:N:P生态化学计量特征(图 4):随着P添加量增加, 有机C含量和全N含量没有明显的变化, 全P含量则持续增高; C:N比呈现先增加后降低的变化趋势, 其最大值出现在N5P1处理下。C:P比亦呈现出先增加后降低的变化趋势, 但少量P添加的影响未达到显著水平。N:P比呈现逐渐降低的趋势, 且在N5P8和N5P16处理下达到显著水平。

|
| 图 4 N和P添加对土壤C:N:P生态化学计量特征的影响 Fig. 4 Effects of N and P addition on soil C:N:P ecological stoichiometry N5P0、N5P0.5、N5P1、N5P2、N5P4、N5P8和N5P16代表在统一施用5.0 g N/m2/a的基础上, 分别施入0.0、0.5、1.0、2.0、4.0、8.0和16.0 g m-2 a-1的P; 不同小写字母表示同一指标在N和P添加处理间差异显著(P < 0.05), 相同字母表示差异不显著(P>0.05) |
对植物群落多样性指数与土壤因子的关系进行了RDA排序(图 5):前两个排序轴的特征值分别为0.787和0.208, 占总特征值的99.5%。植物群落多样性指数与土壤因子两个排序轴的相关度均为1, 表明前两个轴的物种环境相关度很高, 共解释物种和环境总方差的99.5%。

|
| 图 5 植物群落多样性指数与土壤因子关系的RDA排序图 Fig. 5 RDA ordination diagram of the relationships between plant community diversity indices and soil factors R、H、E和D分别代表Patrick丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数; SOC、TN、TP、NH4+-N、NO3--N、AP、C:Ns、C:Ps、N:Ps、C:Nm、C:Pm和N:Pm分别代表土壤有机C含量、土壤全N含量、土壤全P含量、土壤NH4+-N浓度、土壤NO3--N浓度、土壤速效P浓度、土壤C:N比、土壤C:P比、土壤N:P比、微生物量C:N比、微生物量C:P比和微生物量N:P比 |
图 5中, 实线箭头越长土壤因子对植物群落多样性的影响程度越高; 实线箭头与虚线箭头之间的夹角表示土壤因子与植物群落多样性指数的相关关系, 锐角为正相关, 钝角为负相关。根据图 5中箭头的长度可以看出, 对植物群落多样性影响最大的土壤因子分别是土壤N:P比、土壤全P含量、土壤速效P浓度和微生物量C:P比。根据实线与虚线的夹角可以看出, Patrick丰富度指数分别与土壤N:P比、C:P比以及NH4+-N浓度正相关, 与土壤全P含量、速效P浓度以及微生物量C:P比负相关; Shannon-Wiener多样性指数分别与土壤N:P比、C:P比以及NH4+-N浓度正相关, 与土壤全P含量和速效P浓度负相关; Simpson优势度指数分别与土壤全P含量和速效P浓度正相关, 与土壤N:P比、C:P比以及NH4+-N浓度负相关; Pielou均匀度指数分别与土壤N:P比和微生物量C:P比正相关, 与土壤全P含量和速效P浓度负相关。
3 讨论 3.1 少量N添加下增施P肥对荒漠草原植物群落组成的影响研究发现, 在P受限制的情况下降低N的添加量或提高P的添加量会促进植物生物量的积累[23]。对青藏高原高寒草甸植物[24]、内蒙古温带典型草原植物[25]以及陕西黄土丘陵森林-草地区植物[26]的研究进一步证实, N添加下增施P肥依然对植物生长有促进作用。本研究中, 少量N添加下随着施P量增加, 植物群落和种群生物量均表现出先增加后降低的趋势。这可能是因为宁夏荒漠草原土壤P有效性比较低, 而5.0 g m-2 a-1的N添加进一步加剧了土壤P限制。逐渐施入的KH2PO4提高了土壤P的供给能力, 缓解了N添加引起的土壤和植物间的P供需压力, 因此促进了植物生物量的积累。然而, 持续增加的KH2PO4添加量可能超出了植物自身的内稳态调节能力[27]。此时, P肥在植物体内的过量积累, 不仅产生了毒害作用, 而且使N受限性增加, 从而抑制了牛枝子、草木樨状黄芪、苦豆子和猪毛菜地上部分的生长, 导致植物群落和种群生物量下降(图 1和图 2), 与甘肃亚高寒草甸植物的结果一致[28]。
N添加下, 植物P受限性增加是群落多样性降低的重要机制之一[5, 15]。增施P肥增强了土壤P有效性, 从而提高了植物群落多样性[29]。本研究中, 适量增施P肥提高了Shannon-Wiener多样性指数和Patrick丰富度指数; 中高量增施P肥降低了植物群落多样性, 与青藏高原高寒草甸的研究结果类似[30]。N和P共同添加下植物群落组成的变化趋势, 主要取决于物种竞争和共存关系的改变[31-33]。适量P添加下几个主要植物种重要值变化幅度小, 表明土壤肥力水平较高时, 几个物种具有较好的共存关系(表 1)。随着P肥的持续施入, 几个豆科植物重要值降低, 猪毛蒿重要值增加。其可能原因, 一方面在于中高量P添加对牛枝子和苦豆子等多年生植物根系产生的P毒效应; 另一方面在于降水在介导N和P肥效中发挥了重要作用。研究区2015年和2016年降水量分别为365.6 mm和347.8 mm(高于多年平均值, 且主要集中在4—9月), 雨水丰沛期与施肥时间(每年5—8月)基本一致。此外, 为提高肥料利用效率, 我们在进行N和P处理时大多选择雨后的傍晚施入。因此, 在水肥资源较为丰富的情况下, 一、二年生藜科植物猪毛蒿凭借其较强的生存能力(R对策)开始大量繁殖[34-36], 与其他物种产生资源竞争关系, 导致群落多样性降低。
3.2 少量N添加下增施P肥对荒漠草原土壤C:N:P生态化学计量特征的影响研究表明, 短期N添加对土壤C含量的影响无一致的规律性, 但可以提高N和P有效性[37]。随着N的逐渐输入, 土壤N有效性上升, N和P相对稳定的平衡关系受到影响。N添加下元素的非同步变化, 导致土壤N:P比升高、C:N比降低[13]。增施P肥提高了P与土壤有机物的结合、加速了有机物分解和养分释放, 从而降低了土壤N:P和C:P比[38-39], 因此可能会缓解N添加引起的土壤元素化学计量比失衡。本研究中, 土壤C:N比变化较小, 反映了环境变化下较为紧密的C和N耦合关系[11]; 由于试验样地土壤类型为灰钙土, 土壤中游离态的CaCO3易与P结合形成溶解度较低的磷酸钙盐[40-41]。因此少量增施P肥对土壤C:P和N:P比影响较小, 但高量P添加显著降低了二者。低的土壤C:P和N:P比意味着生态系统主要受N限制, 反之则意味着主要为P限制[8-9]。结合植物生物量和群落多样性的变化特点, 以上结果可能证实了适量P添加有助于缓解土壤P限制, 但高P添加反而会导致植物群落N受限性增强。
3.3 植物群落多样性与土壤C:N:P生态化学计量特征及其他关键因子的关系依据利比希(Liebig)最小因子定律和Shelford耐性定律, 土壤中N和P的相对丰富或不足影响着植物N和P利用策略。因此土壤C:N:P平衡关系在维持植物群落结构方面发挥着重要的作用[42-44]。针对沙地的研究发现, 植物群落多样性随土壤C:N和N:P比增加而增加[45]。本研究中, 多样性指数与土壤N:P和C:P比正相关, 与土壤全P含量和速效P浓度负相关(图 5)。综合植物群落组成和土壤C:N:P比的变化情况而言, 以上结果进一步证实P添加增强了土壤P供给能力, 调节了土壤元素化学计量平衡关系, 从而提高了植物群落多样性; 但随着P的持续施入, 土壤C:P和N:P比降低幅度过大, 导致多数植物在群落中的优势度降低、植物群落组成趋于单一; 相比之下, C和N间较为紧密的耦合关系(图 4), 使得土壤C:N比对植物群落多样性的影响较小, 与针对沙地植物群落的研究结果不同; 此外, 微生物量C:P比与多样性指数具有一定的相关性, 不仅反映了微生物和植物对土壤P资源的竞争与依存关系, 而且意味着微生物对P的固持与释放会影响到植物群落结构[10, 12]。
4 结论综合以上分析, 少量N添加下增施P肥促进了植物生长发育, 但过量P添加降低了植物群落多样性; N沉降增加下趋于解耦的土壤元素计量平衡关系会直接影响到植物群落组成; 适量P添加可以通过降低土壤C:P和N:P比、提高凋落物归还量和刺激微生物P释放等直接或间接途径, 提高土壤P的可利用性, 从而调节土壤P供给和植物P需求间的压力、缓解N添加引起的植物群落多样性降低。然而, 过量P添加不仅增加了P毒风险, 而且引起了N限制, 导致多数植物生长受阻和群落组成单一。由于本项研究仅是连续两年N和P添加后的野外试验观测结果, 而短期控制试验效应易受到降水等气候因子变化的影响, 因此有必要通过长期的定位观测对本项研究结果进行反复验证。
| [1] |
Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle:recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889-892. DOI:10.1126/science.1136674 |
| [2] |
郑丹楠, 王雪松, 谢绍东, 段雷, 陈东升. 2010年中国大气氮沉降特征分析. 中国环境科学, 2014, 34(5): 1089-1097. |
| [3] |
Liu X J, Duan L, Mo J M, Du E Z, Shen J L, Lu X K, Zhang Y, Zhou X B, He C N, Zhang F S. Nitrogen deposition and its ecological impact in China:an overview. Environmental Pollution, 2011, 159(10): 2251-2264. DOI:10.1016/j.envpol.2010.08.002 |
| [4] |
顾峰雪, 黄玫, 张远东, 闫慧敏, 李洁, 郭瑞, 钟秀丽. 1961-2010年中国区域氮沉降时空格局模拟研究. 生态学报, 2016, 36(12): 3591-3600. |
| [5] |
Li Y, Niu S L, Yu G R. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading:a meta-analysis. Global Change Biology, 2016, 22(2): 934-943. DOI:10.1111/gcb.13125 |
| [6] |
Vitousek P M, Porder S, Houlton B Z, Chadwick O A. Terrestrial phosphorus limitation:mechanisms, implications, and nitrogen-phosphorus interactions. Ecological Applications, 2010, 20(1): 5-15. DOI:10.1890/08-0127.1 |
| [7] |
Sterner R W, Elser J J. Ecological Stoichiometry:the Biology of Elements from Molecules to the Biosphere. Princeton: Princeton University Press, 2002.
|
| [8] |
Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C:N:P ratios in China's soils:a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139-151. |
| [9] |
王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937-3947. DOI:10.3321/j.issn:1000-0933.2008.08.054 |
| [10] |
Zechmeister-Boltenstern S, Keiblinger K M, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecological Monographs, 2015, 85(2): 133-155. DOI:10.1890/14-0777.1 |
| [11] |
Yang Y H, Fang J Y, Ji C J, Datta A, Li P, Ma W H, Mohammat A, Shen H H, Hu H F, Knapp B O, Smith P. Stoichiometric shifts in surface soils over broad geographical scales:evidence from China's grasslands. Global Ecology and Biogeography, 2014, 23(8): 947-955. DOI:10.1111/geb.12175 |
| [12] |
Mayor J R, Mack M C, Schuur E A G. Decoupled stoichiometric, isotopic, and fungal responses of an ectomycorrhizal black spruce forest to nitrogen and phosphorus additions. Soil Biology and Biochemistry, 2015, 88: 247-256. DOI:10.1016/j.soilbio.2015.05.028 |
| [13] |
Yue K, Fornara D A, Yang W Q, Peng Y, Li Z J, Wu F Z, Peng C H. Effects of three global change drivers on terrestrial C:N:P stoichiometry:a global synthesis. Global Change Biology, 2017, 23(6): 2450-2463. DOI:10.1111/gcb.13569 |
| [14] |
Stevens C J, Thompson K, Grime J P, Long C J, Gowing D J G. Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition. Functional Ecology, 2010, 24(2): 478-484. DOI:10.1111/j.1365-2435.2009.01663.x |
| [15] |
Zhan S X, Wang Y, Zhu Z C, Li W H, Bai Y F. Nitrogen enrichment alters plant N:P stoichiometry and intensifies phosphorus limitation in a steppe ecosystem. Environmental and Experimental Botany, 2017, 134: 21-32. DOI:10.1016/j.envexpbot.2016.10.014 |
| [16] |
段雷, 郝吉明, 谢绍东, 周中平. 用稳态法确定中国土壤的硫沉降和氮沉降临界负荷. 环境科学, 2002, 23(2): 7-12. |
| [17] |
Clark C M, Tilman D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451(7179): 712-715. DOI:10.1038/nature06503 |
| [18] |
黄菊莹, 余海龙. 四种荒漠草原植物的生长对不同氮添加水平的响应. 植物生态学报, 2016, 40(2): 165-176. |
| [19] |
黄菊莹, 余海龙, 刘吉利, 马飞, 韩磊. 控雨对荒漠草原植物、微生物和土壤C、N、P化学计量特征的影响. 生态学报, 2018, 38(15): 5362-5373. |
| [20] |
Huang J Y, Yu H L, Lin H, Zhang Y, Searle E B, Yuan Z Y. Phosphorus amendment mitigates nitrogen addition-induced phosphorus limitation in two plant species in a desert steppe, China. Plant and Soil, 2016, 399(1/2): 221-232. |
| [21] |
张金屯. 数量生态学. 北京: 科学出版社, 2004.
|
| [22] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000.
|
| [23] |
Güsewell S. Responses of wetland graminoids to the relative supply of nitrogen and phosphorus. Plant Ecology, 2005, 176(1): 35-55. |
| [24] |
杨晓霞, 任飞, 周华坤, 贺金生. 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应. 植物生态学报, 2014, 38(2): 159-166. |
| [25] |
何利元, 胡中民, 郭群, 李胜功, 白文明, 李凌浩. 氮磷添加对内蒙古温带草地地上生物量的影响. 应用生态学报, 2015, 26(8): 2291-2297. |
| [26] |
张少康, 刘海威, 焦峰. 黄土丘陵区微地形梯度下草地群落及土壤对氮、磷添加的响应. 水土保持研究, 2018, 25(1): 76-83. |
| [27] |
Güsewell S. N:P ratios in terrestrial plants:variation and functional significance. The New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [28] |
张仁懿, 史小明, 李文金, 王刚, 郭睿. 氮、磷添加对亚高寒草甸地上生物量的影响. 生态科学, 2016, 35(5): 15-20. |
| [29] |
赵一楠.氮、磷添加及AM真菌调控对松嫩草地土壤养分与植物群落特征的影响[D].长春: 东北师范大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10200-1017142600.htm
|
| [30] |
Li J H, Yang Y J, Li B W, Li W J, Wang G, Knops J M H. Effects of nitrogen and phosphorus fertilization on soil carbon fractions in alpine meadows on the Qinghai-Tibetan Plateau. PLoS One, 2014, 9(7): e103266. DOI:10.1371/journal.pone.0103266 |
| [31] |
Bai W M, Guo D L, Tian Q Y, Liu N N, Cheng W X, Li L H, Zhang W H. Differential responses of grasses and forbs led to marked reduction in below-ground productivity in temperate steppe following chronic N deposition. Journal of Ecology, 2015, 103(6): 1570-1579. DOI:10.1111/1365-2745.12468 |
| [32] |
Xu X L, Ouyang H, Cao G M, Richter A, Wanek W, Kuzyakov Y. Dominant plant species shift their nitrogen uptake patterns in response to nutrient enrichment caused by a fungal fairy in an alpine meadow. Plant and Soil, 2011, 341(1/2): 495-504. |
| [33] |
Peng Y F, Yu P, Zhang Y, Sun G, Ning P, Li X X, Li C J. Temporal and spatial dynamics in root length density of field-grown maize and NPK in the soil profile. Field Crops Research, 2012, 131: 9-16. DOI:10.1016/j.fcr.2012.03.003 |
| [34] |
Wilson J B, Lee W G. C-S-R triangle theory:community-level predictions, tests, evaluation of criticisms, and relation to other theories. Oikos, 2000, 91(1): 77-96. DOI:10.1034/j.1600-0706.2000.910107.x |
| [35] |
杜峰, 山仑, 陈小燕, 梁宗锁. 陕北黄土丘陵区撂荒演替研究-撂荒演替序列. 草地学报, 2005, 13(4): 328-333. |
| [36] |
李璠.施肥对高寒草原群落结构的影响[D].北京: 中国科学院大学, 2013. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2440317
|
| [37] |
周纪东, 史荣久, 赵峰, 韩斯琴, 张颖. 施氮频率和强度对内蒙古温带草原土壤pH及碳、氮、磷含量的影响. 应用生态学报, 2016, 27(8): 2467-2476. |
| [38] |
白玉婷, 卫智军, 代景忠, 闫瑞瑞, 刘文亭, 王天乐. 施肥对羊草割草地植物群落和土壤C:N:P生态化学计量学特征的影响. 生态环境学报, 2017, 26(4): 620-627. |
| [39] |
俄胜哲, 杨志奇, 曾希柏, 王亚男, 罗照霞, 袁金华, 车宗贤. 长期施肥黄绵土有效磷含量演变及其与磷素平衡和作物产量的关系. 应用生态学报, 2017, 28(11): 3589-3598. |
| [40] |
Tunesi S, Poggi V, Gessa C. Phosphate adsorption and precipitation in calcareous soils:the role of calcium ions in solution and carbonate minerals. Nutrient Cycling in Agroecosystems, 1999, 53(3): 219-227. DOI:10.1023/A:1009709005147 |
| [41] |
Cross A F, Schlesinger W H. Biological and geochemical controls on phosphorus fractions in semiarid soils. Biogeochemistry, 2001, 52(2): 155-172. DOI:10.1023/A:1006437504494 |
| [42] |
Zhang X N, Yang X D, Li Y, He X M, Lv G H, Yang J J. Influence of edaphic factors on plant distribution and diversity in the arid area of Xinjiang, Northwest China. Arid Land Research and Management, 2018, 32(1): 38-56. DOI:10.1080/15324982.2017.1376004 |
| [43] |
Busch V, Klaus V H, Penone C, Schäfer D, Boch S, Prati D, Muller J, Socher S A, Niinemets Ü, Peñuelas J, Hölzel N, Fischer M, Kleinebecker T. Nutrient stoichiometry and land use rather than species richness determine plant functional diversity. Ecology and Evolution, 2018, 8(1): 601-616. DOI:10.1002/ece3.3609 |
| [44] |
Jiao F, Wen Z M, An S S. Changes in soil properties across a chronosequence of vegetation restoration on the Loess Plateau of China. Catena, 2011, 86: 110-116. DOI:10.1016/j.catena.2011.03.001 |
| [45] |
杨祥祥, 李梦琦, 何兴东, 尤万学, 余殿, 张彩华, 陈娜. 沙地土壤C:N:P比对早期植物群落物种多样性的影响. 土壤学报, 2019, 56(1): 242-249. |