 2019, Vol. 39
2019, Vol. 39文章信息
- 刘新春, 赵勇钢, 刘小芳, 朱兴菲, 高冉, 栗文玉
- LIU Xinchun, ZHAO Yonggang, LIU Xiaofang, ZHU Xingfei, GAO Ran, LI Wenyu
- 晋西黄土区人工林细根与土壤水碳的耦合关系
- Coupling fine roots with soil moisture and organic carbon in artificial forests in loess region of western Shanxi Province
- 生态学报. 2019, 39(21): 7987-7995
- Acta Ecologica Sinica. 2019, 39(21): 7987-7995
- http://dx.doi.org/10.5846/stxb201901120101
-
文章历史
- 收稿日期: 2019-01-12
- 网络出版日期: 2019-09-05

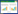




人工植被建设是干旱半干旱地区控制水土流失、扭转土地退化、改善区域生态环境的一项重要举措[1]。黄土高原大面积人工植被恢复对深层土壤水分利用及有机碳积累的影响是目前研究关注的重要科学问题。人工林作为广泛栽植的木本植物, 有利于减少地表径流, 改善土壤结构, 增加土壤碳储量[2-4]。然而, 不当的人工林种植也可能带来负面的影响, 尤其是引进外来植被引起的深层土壤干燥化[5]、土地退化[6]等问题日益凸显。近年来, 国内外关于人工林引起的土壤水分分布及碳储量的差异性已有诸多研究。杨磊等[7]针对甘肃龙滩流域不同人工植被土壤水分的研究发现, 柠条、油松、山杏林地土壤水分亏缺严重, 尤其是100 cm以下土层。Yuan等[8]研究认为木本植物的高蒸腾特性造成的土壤水分损失可能会加剧深层土壤水分短缺。杨景成等[9]对退耕农田转变的草地和次生林的研究发现, 植被恢复可增加19%—53%的土壤碳储量。邱甜甜等[10]对油松林与撂荒地的对比研究发现, 不同生长阶段的油松林其浅层和深层有机碳储量均显著大于撂荒地, 证实了油松林建设对于提高土壤有机碳储量有显著作用。王征等[11]通过对纸坊沟流域刺槐林土壤有机碳含量的研究发现, 其深土层(50—200 cm)含量约为浅层(0—50 cm)的25%, 证明了植被恢复对深层土壤有机碳的固定有重要影响。
细根(≤2 mm)是根系系统中最敏感的根, 是植物吸收水分和养分的重要途径[12]。植物对土壤水分和养分的竞争能力取决于其根系占据的土壤空间, 这一能力由一些根系特征决定, 如根密度、生物量、根表面积、根深等。近年来, 随着退耕还林还草工程的深入推进, 人工林生态系统中细根的变化规律受到了国内外学者的广泛关注。赵忠等[13]对安塞刺槐的细根表面积密度的研究发现, 细根分布密集区对应土层的土壤水分状况变化较为活跃, 认为其分布与土壤含水量随土层深度的变化有相似趋势。杨秀云等[14]对华北落叶松细根生物量的空间分布研究发现, 细根生物量与土壤含水量存在显著的相关关系。韩凤朋等[15]对裸地坡面和草坡地的对比研究结果显示, 根系密集区的土壤养分状况优于无根系存在的土壤, 且在根系密集区, 其土壤有机质具有明显的表聚现象。燕辉等[16]研究认为根径≤2 mm的细根生物量、表面积和根长密度与土壤有机质显著相关。从目前的研究来看, 不同人工植被细根分布具有差异, 并与土壤水碳的分布密切相关, 但在区域间对不同人工林类型, 尤其是深土层的土壤水分、有机碳的影响不够明确[17], 有进一步研究的必要。
晋西黄土丘陵沟壑区位于黄土高原腹地, 降水稀少, 蒸发量远大于降水量, 土壤中养分缺乏, 原始植被破坏严重, 水土流失严重。自20世纪开始, 该区域进行大规模林草植被建设, 刺槐(Robinia pseudoacacia)、侧柏(Platycladus orientalis)因其耐干旱、生长迅速、成活率高等特点[18], 被作为该区域人工植被恢复的适生树种, 核桃(Jugla regia)因其经济价值被广泛种植。目前关于该地区不同人工林类型下土壤水碳分布已有一些研究, 但主要集中在浅层土壤, 有关深剖面(>1 m)土壤水分亏缺和有机碳积累效应的研究相对较少[7, 19], 特别是比较缺乏从植被细根-水分-有机碳三者相互关系开展的研究[20], 这对于深入揭示植被恢复的土壤功能机制有所不足。因此, 本研究选取晋西黄土区3种典型人工林, 对深剖面(0—500 cm)植被细根参数进行研究, 并分析其与土壤水分和有机碳的耦合关系, 以期为区域人工林建设的评价与管理提供一定的科学依据。
1 研究地区与研究方法 1.1 研究区概况试验区位于山西省吕梁市离石区莲花池乡(111°7′39″—111°10′14″E, 37°28′14″—37°28′56″N), 属三川河流域一支, 海拔900—1200 m, 地形支离破碎, 沟壑纵横, 属典型黄土高原丘陵沟壑区。年平均气温8.9℃, 年平均降水量为450—550 mm, 主要集中在7—9月, 占全年降水的70%以上。年蒸发量为1850.8 mm, 最大蒸发量出现在4—6月, 属典型的温带大陆性气候。土壤类型主要为黄土母质直接发育的黄绵土, 颗粒组成以砂粒、粉砂粒为主, 含钙质, 结构松散, 遇水极易崩解, 土壤侵蚀程度较大。在植被区划上, 属于暖温带落叶阔叶林地带, 该地区常见人工造林树种主要有刺槐、侧柏、柠条(Caragana korshikii)等;人工经济林主要有核桃、苹果(Malus pumila)等。各样地基本信息见表 1。
| 样地名称 Sampling plot |
海拔 Elevation/m |
坡度 Slope gradient/(°) |
坡向 Slope aspect |
年限 Age/a |
平均株高 Average plant height/m |
胸/基径 Average DBH/m |
郁闭度/覆盖度 Coverage/% |
草本层优势种 Dominant herb |
种植密度 Stand density/ (株/hm2) |
| 农地Cropland | 1033 | 6 | SW12° | 30 | 2.31 | 1.39 | 87 | 玉米 Zea mays L. |
- |
| 刺槐 Robinia pseudoacacia |
1108 | 12 | EN58° | 17 | 9.23 | 3.94 | 69 | 苔草 Carex tristachya |
1250 |
| 侧柏 Platycladus orientalis |
1083 | 13 | SW4° | 8 | 5.33 | 1.8.7 | 38 | 白羊草 Bothriochloa ischaemum (L.) Keng |
3300 |
| 核桃 Jugla regia |
995 | 9 | NW14° | 16 | 5.74 | 2.64 | 45 | 灰藜 Chenopodium album L. |
1100 |
| DBH:Diameter at breast height | |||||||||
本研究选取该地区广植的3种典型人工林为研究对象, 包括退耕还林常用树种刺槐和侧柏及经济林核桃, 并以长期种植的农地作为对照。采样农地长期耕作(>30a), 为雨养旱作管理模式。种植作物为玉米, 采用单作模式, 一年一熟制。于2017年8月上旬进行土壤和根系样品采集。每个样地选择坡向和坡度相近的坡面, 沿坡面布置3条长50 m、宽10 m的样带, 在样带上沿坡面布置3个样方(乔木10 m×10 m), 每个样地共9个样方。在样方内进行每木检尺, 记录其株高、胸径、冠幅, 并据此选择3株平均木为代表树, 以其主干为中心, 在其半径1 m的弧线正方位上选点, 用根钻(内径9 cm)采集0—500 cm深度包含根系的土样, 其中0—100 cm土层间隔10 cm取样, 100—500 cm土层间隔20 cm取样。另用土钻(内径5 cm)在根钻临近位置采集0—500 cm深度土壤样品, 用于测定土壤含水量和有机碳含量, 采样土层设计同根钻取样。在打钻取样点旁挖取土壤剖面, 用环刀(100 cm3)采集100 cm土层(间隔10 cm)原状土, 用于土壤容重(Bulk density, BD)测定。
1.2.1 根样的获取及处理将野外采集的根系样品用清水过2 mm筛冲洗, 收集细根样品后用游标卡尺确定根系直径大小, 用镊子挑出杂物并拣出直径≤2 mm的全部细根。用根系扫描仪(EPSON PERFECTION V700 Photo)扫描并用WINRhizo根系分析系统进行根系的总根长、根表面积、根平均直径等参数的测定分析, 扫描后的根系装入信封于80℃烘箱中烘干至恒重。细根参数计算公式如下[21]:

|
(1) |
式中, X为相应各指标密度值, 即根长密度(Fine root length density, FRLD, m/m3);根质量密度(Fine root mass density, FRMD, g/m3);根表面积密度(Fine root surface area density, FRAD, m2/m3);xxj为根系指标, 代表根长(m);根干重(g);根表面积(m2);r为根钻半径(cm);h为土层厚度(cm);i和j分别为样本数及样点数;n和k分别为样本总数和样点总数。
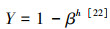
|
(2) |
式中, Y为从地表到一定土层深度的根系生物量累积百分比(%);h为土层深度(cm);β为根系消弱系数。
1.2.2 土壤理化性质测定与计算土壤含水量(Soil moisture content, SMC)采用烘干法(105—110℃, 24 h)测定;土壤有机碳(Soil organic carbon, SOC)测定采用重铬酸钾外加热法;0—100 cm土壤容重采用环刀法测量[23]。由于野外传统的环刀法采样实测工作量大, 100—500 cm的容重通过土壤容重传递函数模型(Pedotrafer function)来预测和估算。选取国内外关于该区域已建立的经典容重传递函数模型, 带入本研究各样地0—100 cm的相关数据检验其预测精度[24], 并进一步建立各样地的最佳容重传递函数模型, 通过逐步回归分析创建回归方程, 对基础模型进行校正优化, 得到最终的各样地容重传递函数模型。
土壤有机碳密度(SOC density, SOCD)是指单位面积一定土体中土壤有机碳质量, 其计算公式为[25]:

|
(3) |
式中, SOCDi和SOCi分别代表第i层的平均SOCD(kg/m2);平均有机碳含量(g/kg);BDi为第i层平均土壤容重(g/cm3);Di为第i层的土壤厚度(cm)(本文以每10 cm深度计);θi为第i层中直径大于2 mm的石砾所占的体积百分比(%)。鉴于黄土高原典型的土壤特性, 几乎没有粒径大于2 mm的砾石, θ值可忽略不计。
1.2.3 土壤水分亏缺和有机碳积累效应评价以农地为对照, 3种人工林(刺槐、侧柏和核桃)深层的土壤水分亏缺效应(soil moisture deficit, SMD)和SOC积累效应(soil organic carbon accumulation, SOCA)计算如下[25]:
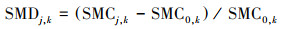
|
(4) |

|
(5) |
式中, SMCj, k、SMC0, k和SOCDj, k和SOCD0, k分别代表第k个土层第j种人工林样地和对照农地的SMC和SOCD。
1.2.4 数据统计及分析根据SMC、SOC和细根参数的土层分布特征, 并结合前人的研究[20, 26], 本研究中将土层分为浅层(0—70 cm)和深层(70—500 cm)进行分析。采用单因素方差分析法(one-way ANOVA)和最小差异显著法(LSD)对3种人工林各根系指标进行统计分析(P < 0.05), 采用Pearson相关系数对细根、SMC和SOCD进行相关性分析。所有统计分析用SPSS 18.0进行。采用Origin 9.0软件绘图。
2 结果与分析 2.1 人工林细根特征参数分布4种人工植被整个土层(0—500 cm)细根特征参数分布如图 1所示。细根特征参数在浅层(0—70 cm)和深层(70—500 cm)有显著差异(P < 0.05)。4种人工植被土壤浅层细根累计生物量达到整个土层的52%—71%, 其FRLD值范围为658.78—1308.27 m/m3, FRMD值范围为46.60—418.34 g/m3;在深土层FRLD值范围为265.57—409.10 m/m3, FRMD值范围为13.12—62.00 g/m3, 3种人工林FRLD和FRMD显著大于农地(P < 0.05), 但彼此间差异不显著。FRAD具有较为相似的分布规律, 其浅层范围为1.41—3.96 m2/m3;深土层为1.84—4.06 m2/m3, 仅在农地和核桃样地有显著差异(P < 0.05);各样地细根直径主要集中在1 mm左右, 农地细根直径显著小于3种人工林细根直径(P<0.05)。

|
| 图 1 4种人工植被0—500 cm剖面细根特征参数分布 Fig. 1 Fine root parameters within the 0—500 cm soil layers under the four vegetation types |
4种植被土壤水分(SMC)在整个土层的分布有较大差异(图 2)。在土壤浅层, 各样地SMC随土壤深度增加表现为增加趋势, 且变异性较大;4种人工植被深土层平均SMC为9.8%—16.4%, 各样地变异性相对较小。以农地为对照, 3种人工林SMC均存在不同程度的亏缺(图 2)。3种人工林在剖面的亏缺状况与其SMC趋势基本一致。3种人工林浅层水分亏缺(SMD)随土壤深度增加呈增加趋势;在深土层, 刺槐和侧柏在70—300 cm土层SMD持续且较为稳定, 而核桃SMD呈减少趋势;300—500 cm土层刺槐和核桃SMD呈增加趋势, 而侧柏SMD呈减少趋势。3种人工林整个土层的SMD效应如图 2所示。就整个土层来看, 各样地浅层SMD均显著高于深层(P < 0.05)。在土壤浅层, 其SMD值表现为:刺槐(-0.36)高于侧柏(-1.00), 核桃(-1.23)次之, 各样地间无显著差异(P>0.05)。在深土层(70—500 cm), 其SMD值表现为:侧柏(-5.37)显著高于核桃(-9.08), 刺槐(-10.73)次之。总体来说, 刺槐水分亏缺最为严重, 核桃次之, 侧柏最轻。

|
| 图 2 4种人工植被0—500 cm剖面土壤水分含量(SMC)分布及亏缺特征(SMD) Fig. 2 Soil moisture content (SMC) and its deficit (SMD) within the 0—500 cm soil layers under the four vegetation types 图中阴影部分面积为各样地与空白对照相比的土壤水分亏缺值, 图c中各样地横坐标统一为10 |
4种人工植被浅层平均有机碳密度(SOCD)范围在0.82—1.58 kg/m2, 占整个土层的14.0%—24.0%, 农地和核桃显著高于刺槐和侧柏(P < 0.05)。4种人工植被深层平均SOCD范围在0.99—1.26 kg/m2, 各样地间无显著差异(P>0.05)。以农地为对照, 3种人工林在0—500 cm的有机碳积累(SOCA)效应表现不同。在土壤浅层, 仅核桃SOCA为正值, 在土壤深层, 刺槐和侧柏的SOCA随土层深度的增加呈现增加趋势, 而核桃的SOCA先减少后增加, 400 cm以下土层, 3种人工林SOCA趋于正值且逐渐趋于稳定。就整个土层来看, 除刺槐外, 各样地深层SOCA显著高于浅层(P < 0.05)(图 3)。在土壤浅层表现为:核桃(1.47)显著高于刺槐(-1.46)和侧柏(-1.64), 在深土层表现为:核桃(4.95)和侧柏(3.55)显著高于刺槐(-0.66)。总体来看, 核桃和侧柏SOCA累计为正值, 具有累积效应, 而刺槐累计为负值, 具有亏缺效应。

|
| 图 3 4种人工植被0—500 cm剖面有机碳密度(SOCD)分布及其积累效应(SOCA) Fig. 3 Soil organic carbon density (SOCD) and accumulation (SOCA) within the 0—500 cm soil layers under the four vegetation types 图中阴影部分面积为各样地与空白对照相比的土壤有机碳亏缺值, 图c中各样地横坐标统一为10 |
Pearson相关性分析表明(表 2), 在整个土层, 各样地SMC、SOCD与细根三者有显著相关关系, 但在不同土层各人工植被有所差异。在土壤浅层, FRLD、FRMD、FRAD与SMC显著负相关, 与SOCD显著正相关(P < 0.05);细根消弱系数β与SMC显著正相关, 与SOCD显著负相关(P < 0.05);D与SMC和SOCD无显著差异(P>0.05);SMC与SOCD显著负相关(P < 0.05)。在土壤深层, 各样地FRLD、FRMD、FRAD与SOCD有显著正相关性(P < 0.05);而与SMC的正相关性仅在刺槐样地显著, SMC与SOCD显著正相关。
| 变量 Variable |
0—70 cm土层Soil layers | 70—500 cm土层Soil layers | 0—500 cm土层Soil layers | |||||||||||
| 农地 | 刺槐 | 侧柏 | 核桃 | 农地 | 刺槐 | 侧柏 | 核桃 | 农地 | 刺槐 | 侧柏 | 核桃 | |||
| SMC与SOCD | ||||||||||||||
| SOCD | -0.95** | -0.78* | -0.94** | -0.97** | -0.22 | 0.64** | -0.30 | 0.53** | -0.39* | 0.57** | -0.27 | 0.54** | ||
| SMC与细根特征参数 | ||||||||||||||
| FRLD | -0.98** | -0.67 | -0.97** | -0.12 | 0.28 | 0.87** | 0.27 | -0.10 | -0.23 | 0.82** | 0.16 | 0.40* | ||
| FRMD | -0.60 | -0.85* | -0.94** | -0.24 | 0.11 | 0.75** | 0.02 | -0.15 | -0.10 | 0.86** | 0.11 | 0.50** | ||
| FRAD | -0.39 | -0.61 | -0.99** | 0.34 | -0.24 | 0.23 | 0.01 | 0.11 | -0.22 | 0.57** | 0.09 | 0.51** | ||
| D | 0.55 | 0.81* | 0.16 | 0.47 | -0.11 | -0.61** | -0.23 | 0.25 | 0.05 | 0.55** | 0.10 | -0.23 | ||
| β | 0.91** | 0.79* | 0.88** | 0.94** | -0.31 | -0.62** | -0.28 | -0.37 | 0.37* | -0.43* | 0.07 | -0.25 | ||
| SOCD与细根特征参数 | ||||||||||||||
| FRLD | 0.95** | 0.51 | 0.94** | 0.25 | 0.66** | 0.59** | -0.36 | 0.18 | 0.94** | 0.75** | 0.615** | 0.55** | ||
| FRMD | 0.73 | 0.65 | 0.86* | 0.33 | 0.72** | 0.54** | -0.39 | 0.27 | 0.79** | 0.77** | 0.61** | 0.69** | ||
| FRAD | 0.29 | 0.35 | 0.96** | -0.21 | 0.59** | 0.45* | -0.45* | 0.39 | 0.10 | 0.57** | 0.57** | 0.48** | ||
| D | -0.62 | -0.55 | -0.09 | -0.47 | -0.57** | -0.51 | -0.02 | 0.62* | 0.12 | 0.19 | 0.26 | 0.17 | ||
| β | -0.89** | -0.85* | -0.98** | -0.89** | -0.40 | -0.35 | 0.36 | -0.73** | -0.92** | -0.88** | -0.77** | -0.87** | ||
| *, P < 0.05; **, P < 0.01; SMC:土壤含水量Soil moisture content; SOCD:土壤有机碳密度Soil organic carbon density; FRLD:根长密度Fine root length density; FRMD:根质量密度Fine root mass density; FRAD:根表面积密度, Fine root surface area density; D:根直径, Fine root diameter; β:根系消弱系数Root extinction coefficient | ||||||||||||||
在本研究中, 3种人工林深土层细根特征值显著高于农地, 且随土层深度增加有不同程度下降。Bennett等[27]对西部红杉的研究也有类似的结果, 他们认为人工林根系大的分布空间造成了明显优势的细根密度特征值。3种人工林土壤深层细根生物量占细根总量表现为:侧柏(28.9%)#60;刺槐(43.2%)#60;核桃(44.0%), 说明侧柏细根的表聚现象优于刺槐和核桃。荐圣淇等[28]的研究表明刺槐的细根主要分布在0—80 cm土层范围内。王迪海等[29]在安塞县的研究认为, 0 — 200 cm土层是树木细根表面积的主要分布层, 有80%以上的细根表面积分布。这些研究结果与本研究相似。成向荣等[30]的研究显示, 泾川和安塞0—200 cm土层刺槐平均细根FRAD值为2.37和1.71 m2/m3, 侧柏为3.01和2.56 m2/m3, 与本研究(2.01 m2/m3)有所差别, 这可能是由于气候和土壤资源有效性的差异造成的。多数细根直径较小, 主要集中在1 mm左右, 随深度增加根量分布越少且直径相对较小, 因此深层细根可能主要以吸收水分为主[31]。
3.2 人工林细根对土壤水分的影响黄土高原地区气候干旱, 降雨补给十分有限, 深层渗漏很难发生[32], 因此植被根系分布及其吸水能力的差异是造成土壤水分差异的主要原因[33]。在本研究中3种人工林均呈现不同程度的水分亏缺, 其中刺槐亏缺最为严重, 核桃和侧柏次之。在土层深度上, 各样地深层土壤水分亏缺显著高于浅层。Gao等[25]在天河流域的研究表明, 刺槐的水分亏缺程度最大, 柠条次之, 侧柏相对较轻, 且各样地200 cm以下亏缺明显, 与本研究结果类似。这与植物根系的分布特征有直接关系。侧柏深层的细根分布较刺槐和核桃少, 因此其深层水分亏缺较少, 刺槐和核桃则相反。细根对土壤水分的利用在植被类型间有所差异, 且在不同深度差异显著。在土壤浅层, 植物主要利用发达的根系吸收土壤水分, 因此上层细根特征值较大。细根的高密度分布对土壤水分需求也逐渐增大, 加之降水补充不足, 已造成土壤浅层一定程度水分亏缺, 从而导致根系向深扎, 来获取更深层次的土壤水分来维持自身需要, 因此相比农地来说, 人工林深层细根密度值较大。由于根系强烈的吸水作用且多年降水难以补给, 土壤水库已失去调节功能, 因此导致深层土壤水分亏缺严重[34]。土壤水分无法满足细根生长需求, 细根特征密度就会下降[35]。但有许多研究表明[19, 36], 尽管深层细根分布较少, 其对水分的吸收作用仍较为强烈。相关性分析也进一步证实, 细根与土壤水分的相关关系仅在刺槐有显著性, 这可能与不同人工林根系类型对水分的利用程度有关。
3.3 人工林细根对土壤有机碳的影响在本研究中, 3种人工林浅层平均SOCD为0.82—1.58 kg/m2。黄政等[37]在陇南地区的研究中, 侧柏林地浅层(0—50 cm)平均SOCD为2.79 kg/m2, 与本研究有所差别。这主要是因为人工林表层SOC的主要来源为枯枝落叶, 而较本地区来说, 陇南地区高降水量更加有利于有机物质的转化和积累[38]。从土壤剖面来看, 本研究中, 3种人工林深层SOCD占整个土层的77%—86%;且各样地深层SOCD积累效应显著高于浅层。相关性分析也进一步表明, 3种人工林细根与土壤有机碳存在显著正相关关系。张帅等[39]对子午岭人工油松林的研究发现, 深层(100—200 cm)土壤有机碳含量占整个剖面(0—200 cm)的27%—42%, 人工油松林提高了深层有机碳储量, 与本研究结果相似。这证实了尽管深层细根分布有限, 但对于土壤有机碳的积累仍有较大贡献[40]。相比农地来说, 3种人工林中侧柏和核桃SOCD表现为正向积累作用, 而刺槐则相反。这主要是因为不同植被细根分布的差异性主导着有机碳的积累水平[41]。本研究中, 3种人工林细根分布深度及生物量远高于农地。一方面, 植物在深土层的细根分布增加了植被地下生物量、根系分泌物(有机酸、氨基酸和其他有机物质)以及根系脱落物[42], 其分解腐殖化过程大大提高了深层SOC的含量;另一方面, 细根的分布改善了土壤团聚体结构, 从而有利于SOC的积累[43]。而刺槐在深层存在严重的水分亏缺, 水分胁迫降低了有机物质的归还[44], 可能影响到有机碳的积累。
4 结论(1) 3种人工林细根主要集中分布在土壤浅层(0—70 cm), 占细根总量约为56%—71%, 其细根特征值显著高于农地。
(2) 3种人工林深层(70—500 cm)土壤水分亏缺显著高于浅层;与农地相比, 3种人工林在深剖面均有不同程度的水分亏缺, 其中刺槐亏缺最为严重, 核桃次之, 侧柏再次之。
(3) 3种人工林土壤深层有机碳密度贡献较大, 占整个土层(0—500 cm)约为77%—86%;总体来看, 在0—500 cm土层, 3种人工林中侧柏和核桃有机碳积累效应表现为正向积累作用, 刺槐为负向亏缺作用。
(4) 在不同土层, 人工林细根对土壤水碳的贡献有所差异。在土壤浅层, 细根与土壤水分和有机碳密度三者有显著相关关系;在深土层, 细根与有机碳密度有显著相关性, 与土壤水分的相关性仅在刺槐样地较显著。
| [1] |
Fu B J, Liu Y, Lu Y H, He C S, Zeng Y, Wu B F. Assessing the soil erosion control service of ecosystems change in the Loess Plateau of China. Ecological Complexity, 2011, 8(4): 284-293. |
| [2] |
Huang M B, Zhang L, Gallichand J. Runoff respoes to afforestation in a watershed of the Loess Plateau, China. Hydrological Processes, 2003, 17(13): 2599-2609. DOI:10.1002/hyp.1281 |
| [3] |
Li Y Y, Shao M A. Change of soil physical properties under long-term natural vegetation restoration in the Loess Plateau of China. Journal of Arid Environments, 2006, 64(1): 77-96. |
| [4] |
Matamala R, Gonzàlez-Meler M A, Jastrow J D, Norby R J, Schlesinger W H. Impacts of fine root turnover on forest NPP and soil C sequestration potential. Science, 2003, 302(5649): 1385-1387. |
| [5] |
Wang Y Q, Shao M A, Shao H B. A preliminary investigation of the dynamic characteristics of dried soil layers on the Loess Plateau of China. Journal of Hydrology, 2010, 381(1/2): 9-17. |
| [6] |
赵路红, 李昌珍, 康迪, 任成杰, 韩新辉, 佟小刚, 冯永忠. 黄土丘陵区植被恢复对土壤可溶性氮组分的影响. 生态学报, 2017, 37(10): 3533-3542. |
| [7] |
杨磊, 卫伟, 莫保儒, 陈利顶. 半干旱黄土丘陵区不同人工植被恢复土壤水分的相对亏缺. 生态学报, 2011, 31(11): 3060-3068. |
| [8] |
Liu Y X, Zhao W W, Zhang X, Fang X N. Soil water storage changes within deep profiles under introduced shrubs during the growing season: evidence from semiarid Loess Plateau, China. Water, 2016, 8(10): 475-491. |
| [9] |
杨景成, 韩兴国, 黄建辉, 潘庆民. 土地利用变化对陆地生态系统碳贮量的影响. 应用生态学报, 2003, 14(8): 1385-1390. |
| [10] |
邱甜甜, 刘国彬, 王国梁, 孙利鹏, 姚旭. 人工油松林不同生长阶段深层土壤有机碳和活性碳的差异及其影响因素. 自然资源学报, 2016, 31(8): 1399-1409. |
| [11] |
王征, 刘国彬, 许明祥. 黄土丘陵区植被恢复对深层土壤有机碳的影响. 生态学报, 2010, 30(14): 3947-3952. |
| [12] |
Jackson R B, Mooney H A, Schulze E D. A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(14): 7362-7366. |
| [13] |
赵忠, 成向荣, 薛文鹏, 王迪海, 袁志发. 黄土高原不同水分生态区刺槐细根垂直分布的差异. 林业科学, 2006, 42(11): 1-7. |
| [14] |
杨秀云, 韩有志, 武小钢. 华北落叶松林细根生物量对土壤水分、氮营养空间异质性改变的响应. 植物生态学报, 2012, 36(9): 965-972. |
| [15] |
韩凤朋, 郑纪勇, 张兴昌. 黄土退耕坡地植物根系分布特征及其对土壤养分的影响. 农业工程学报, 2009, 25(2): 50-55. |
| [16] |
燕辉, 刘广全, 李红生. 青杨人工林根系生物量、表面积和根长密度变化. 应用生态学报, 2010, 21(11): 2763-2768. |
| [17] |
杨文航, 任庆水, 李昌晓, 宋虹, 袁中勋, 马文超, 崔云风, 王朝英. 三峡库区消落带落羽杉与立柳林土壤微生物生物量碳氮磷动态变化. 生态学报, 2019, 39(5): 1496-1506. |
| [18] |
Jia X X, Shao M A, Zhu Y J, Luo Y. Soil moisture decline due to afforestation across the Loess Plateau, China. Journal of Hydrology, 2017, 546: 113-122. |
| [19] |
邹俊亮, 郭胜利, 李泽, 张芳, 张彦军, 周小刚, 南雅芳. 小流域土壤有机碳的分布和积累及土壤水分的影响. 自然资源学报, 2012, 27(3): 430-439. DOI:10.11849/zrzyxb.2012.03.009 |
| [20] |
Fang X N, Zhao W W, Wang L X, Feng Q, Ding J Y, Liu Y X, Zhang X. Variations of deep soil moisture under different vegetation types and influencing factors in a watershed of the Loess Plateau, China. Hydrology and Earth System Sciences, 2016, 20(8): 3309-3323. |
| [21] |
张良德, 徐学选, 胡伟, 李星. 黄土丘陵区燕沟流域人工刺槐林的细根空间分布特征. 林业科学, 2011, 47(11): 31-36. |
| [22] |
Gale M R, Grigal D F. Vertical root distributio of northern tree species in relation to successional status. Canadian Journal of Forest Research, 1987, 17(8): 829-834. DOI:10.1139/x87-131 |
| [23] |
中国科学院南京土壤研究所. 土壤理化分析. 上海:上海科学技术出版社, 1978, 196-211. |
| [24] |
易小波, 邵明安, 赵春雷, 张晨成. 黄土高原南北样带不同土层土壤容重变异分析与模拟. 农业机械学报, 2017, 48(4): 198-205. |
| [25] |
Gao X D, Li H C, Zhao X N, Ma W, Wu P T. Identifying a suitable revegetation technique for soil restoration on water-limited and degraded land: Coidering both deep soil moisture deficit and soil organic carbon sequestration. Geoderma, 2018, 319: 61-69. |
| [26] |
韩仕峰, 李玉山, 张孝中, 史竹叶. 黄土高原地区土壤水分区域动态特征. 中国科学院水利部西北水土保持研究所集刊, 1989(1): 161-167. |
| [27] |
Bennett J N, Andrew B, Prescott C E. Vertical fine root distributio of western redcedar, western hemlock, and salal in old-growth cedar-hemlock forests on northern Vancouver Island. Canadian Journal of Forest Research, 2002, 32(7): 1208-1216. DOI:10.1139/x02-034 |
| [28] |
荐圣淇, 赵传燕, 方书敏, 余凯. 陇中黄土高原主要造林树种细根生物量分布. 应用生态学报, 2014, 25(7): 1905-1911. |
| [29] |
王迪海, 赵忠, 李剑. 土壤水分对黄土高原主要造林树种细根表面积季节动态的影响. 植物生态学报, 2010, 34(7): 819-826. |
| [30] |
成向荣, 赵忠, 郭满才, 王迪海, 袁志发. 刺槐人工林细根垂直分布模型的研究. 林业科学, 2006, 42(6): 40-48. |
| [31] |
王珺, 刘茂松, 盛晟, 徐驰, 刘小恺, 王汉杰. 干旱区植物群落土壤水盐及根系生物量的空间分布格局. 生态学报, 2008, 28(9): 4120-4127. |
| [32] |
索立柱, 黄明斌, 段良霞, 张永坤. 黄土高原不同土地利用类型土壤含水量的地带性与影响因素. 生态学报, 2017, 37(6): 2045-2053. |
| [33] |
Wang Y Q, Shao M A, Zhang C C, Liu Z P, Zou J L, Xiao J F. Soil organic carbon in deep profiles under Chinese continental monsoon climate and its relations with land uses. Ecological Engineering, 2015, 82: 361-367. |
| [34] |
于洋, 卫伟, 陈利顶, 冯天骄, 杨磊, 张涵丹. 黄土丘陵区坡面整地和植被耦合下的土壤水分特征. 生态学报, 2016, 36(11): 3441-3449. |
| [35] |
李陆生, 赵西宁, 高晓东, 吴普特, 李虹辰, 凌强, 孙文浩. 黄土丘陵区不同树龄旱作枣园细根空间分布特征. 农业工程学报, 2015, 31(20): 140-146. |
| [36] |
胡小宁, 赵忠, 袁志发, 王迪海, 郭满才, 李剑. 黄土高原刺槐林细根生长与土壤水分的耦合关系. 林业科学, 2010, 46(12): 30-35. |
| [37] |
黄政.甘肃陇南山地人工植被土壤有机碳密度及固碳潜力研究[D].兰州: 兰州大学, 2018.
|
| [38] |
佟小刚, 黄绍敏, 徐明岗, 卢昌艾, 张文菊. 长期不同施肥模式对潮土有机碳组分的影响. 植物营养与肥料学报, 2009, 15(4): 831-836. |
| [39] |
张帅, 许明祥, 张亚锋, 王超华, 陈盖. 黄土丘陵区土地利用变化对深层土壤有机碳储量的影响. 环境科学学报, 2014, 34(12): 3094-3101. |
| [40] |
常恩浩, 李鹏, 肖列, 徐国策, 赵宾华, 苏远逸, 冯朝红. 黄土丘陵沟壑区植被恢复演替过程中根系行为特征. 生态学报, 2019, 39(6): 2090-2100. |
| [41] |
Trumbore S E, Davidson E A, De Camargo P B, Nepstad D C, Martinelli L A. Belowground cycling of carbon in forests and pastures of eastern Amazonia. Global Biogeochemical Cycles, 1995, 9(4): 515-528. |
| [42] |
兰志龙, 赵英, 张建国, 李会杰, 司炳成, 焦瑞, Khan M N, Sial T A. 陕北黄土丘陵区不同土地利用方式下土壤碳剖面分布特征. 环境科学, 2018, 39(1): 339-347. |
| [43] |
Poirier V, Roumet C, Munson A D. The root of the matter: Linking root traits and soil organic matter stabilization processes. Soil Biology and Biochemistry, 2018, 120: 246-259. |
| [44] |
Bowden R D, Newkirk K M, Rullo G M. Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions. Soil Biology and Biochemistry, 1998, 30(12): 1591-1597. DOI:10.1016/s0038-0717(97)00228-9 |