 2019, Vol. 39
2019, Vol. 39文章信息
- 郭建忠, 陈作志, 田永军, 张魁, 许友伟, 徐姗楠, 李纯厚
- GUO Jianzhong, CHEN Zuozhi, TIAN Yongjun, ZHANG Kui, XU Youwei, XU Shannan, LI Chunhou
- 胶州湾海域鱼类群落种类组成及多样性
- Species composition and diversity of fish community in Jiaozhou Bay
- 生态学报. 2019, 39(19): 7002-7013
- Acta Ecologica Sinica. 2019, 39(19): 7002-7013
- http://dx.doi.org/10.5846/stxb201808141735
-
文章历史
- 收稿日期: 2018-08-14
- 网络出版日期: 2019-08-16





2. 中国海洋大学水产学院渔业海洋学实验室, 青岛 266003
2. Laboratory of Fishery and Oceanography, College of Fisheries, Ocean University of China, Qingdao 266003, China
胶州湾是青岛的“母亲河”, 位于山东半岛南部, 黄海之滨, 是我国北方典型的半封闭海湾。湾内形状略呈扇形, 生物资源丰富, 是多种经济鱼类繁衍生息的重要场所[1]。20世纪80年代以来, 随着沿岸经济的迅速发展, 大型桥梁工程、水产养殖、滩涂围垦、海上船舶及过度捕捞等人类活动和自然扰动引起胶州湾生态环境恶化, 面积缩小[2], 近岸水域富营养化[3], 鱼类群落生态特征发生变化, 资源衰退, 种类数降低, 优势种以小型低值底层鱼类为主[4-6]。因此, 开展胶州湾鱼类群落组成和多样性研究, 对了解海域目前鱼类资源状况、资源合理开发与管理以及生态恢复具有重要意义。
20世纪80年代至今, 对胶州湾鱼类资源研究主要集中在群落结构[6]、多样性[7]和生物学[8-10]等方面。由于前人调查年代较早、间隔时间长、调查区域有限、且连续作业航次调查较少, 因此, 为了了解胶州湾鱼类群落结构现状特征及长期以来在自然扰动和人类活动干扰下鱼类资源的历史演变状况, 开展胶州湾海域鱼类资源调查研究是必要的。本文根据2016—2017年4个连续航次调查资料, 对胶州湾鱼类群落种类组成及多样性进行了研究;并结合历史资料, 对比分析了20世纪80年代以来胶州湾鱼类种类组成、多样性、资源量以及主要经济鱼类生物学的变化特征, 旨在为胶州湾海域鱼类物种多样性的保护及渔业资源管理提供科学依据。
1 材料与方法 1.1 调查海域胶州湾位于35°38′N—36°18′N, 120°04′E—120°23′E之间, 东岸为青岛市区, 北岸为红岛, 西南岸为黄岛和薛家岛, 受地理位置、陆地气候、水深以及黄海沿岸水流的影响, 胶州湾属于暖温带季风型气候, 四季分明, 1月份最冷, 平均温度为-1.2℃, 8月份最热, 平均温度为25.5℃;其中, 5—11月多在15℃以上, 年平均气温为12.2℃;盐度较高, 约为31.5—32.4, 且春末夏初盐度较高, 雨季受降雨及地面径流影响, 盐度较低;潮汐为典型半日潮, 湾内波浪以风浪为主[1]。海岸线长度为206.8 km, 南北长33 km, 东西宽28 km, 面积约343.1 km2;沿岸入湾河流较多, 有大沽河、洋河、海泊河、李村河等, 且大沽河水流量最大[11]。北部浅海区为菲律宾蛤仔(Ruditapes philippinarum)底播养殖区, 且菲律宾蛤仔是胶州湾目前的主要捕捞对象, 长期过度捕捞使得渔业资源现状处于衰退状态[12]。如1981—1982年以青鳞小沙丁(Sardinella zunasi)和斑鰶(Konosirus punctatus)和带鱼(Trichiurus lepturus)为优势种群[1], 转变为2011年以方氏云鳚(Enedras fangi Wang et Wang)和六丝钝尾鰕虎鱼(Amblychaeturichthys hexanema)为主[6], 且小型低值鱼类成为胶州湾鱼类优势种群[7]。
1.2 站位与样品采集调查航线按照《海洋调查规范》(GB/T12763—2007)和《海洋监测规范》(GB 17378—2007), 渔业资源按照《海洋渔业资源调查规范》(SC/T 9403—2012)进行调查。由于北部浅海区为菲律宾蛤仔底播养殖区, 禁止拖网捕捞, 因此调查站位设置在胶州湾5 m以深水域。采用分层随机取样(stratifiedrandom sampling)方法设计调查站位, 根据水深和地理位置不同, 在湾内、湾外海域共设置9个站点进行海洋生态和渔业资源调查。分别于2016年1月(冬季)、2016年4月(春季)、2016年11月(秋季)和2017年8月(夏季)利用中国水产科学研究院黄海水产研究所“黄海星”号科学调查船进行4个航次调查, 其中, 选取春季7个(无S3和S8)、夏季8个(无S6)、秋季9个和冬季7个(无S3和S5)拖网作业站位进行渔业资源现状调查(图 1)。所有调查中, 每个站位拖曳1次, 每次拖曳1 h, 平均拖曳速度为3.0 kn。捕获样品在现场进行鉴定和分类, 并对每种物种进行生物学测定。

|
| 图 1 胶州湾海域站位采样示意图 Fig. 1 Map of sampling stations in the Jiaozhou Bay “S”:站位Sampling |
依据《海洋调查规范》(GB/T 12763—2007)规范要求, 以Nelson等[13]、李明德等[14]和刘瑞玉等[1]分类系统对渔获鱼类物种进行鉴定与分类, 并依据前人研究来划分鱼类物种的适温类型[15-16]。
1.4 研究方法鱼类物种的生态优势度用Pinkas等[17]提出的相对重要性指数(Index of Relative Important, IRI)来评价。等级划分依据IRI>1000为优势种;100<IRI<1000为重要种;10<IRI<100为常见种, 1<IRI<10为一般种, IRI<1为少有种[18]:
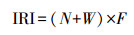
|
(1) |
式中, N为某一鱼类物种尾数占渔获总尾数的百分比;W为某一鱼类种类重量占渔获总重量的百分比;F为某一鱼类种类出现的站位数量占总调查站位数量的百分比。
鱼类群落的生态多样性用Shannon-Wiener多样性指数H′、Margalef种类丰富度指数D′和Pielou均匀度指数J′来分析[19]。由于不同鱼类种间、种内个体差异较大, Wilhm[20]提出用生物量取代个体数量来计算鱼类物种多样性更易接近种类间能量分布, 因此本研究根据渔获鱼类物种生物量来计算鱼类群落的多样性指数H′。

|
(2) |

|
(3) |

|
(4) |
式中, S为鱼类种类数, W为某一渔获鱼类物种的总生物量, Wi为第i站某一渔获鱼类物种的生物量, N为站点渔获鱼类物种的个体数量。
鱼类种类组成的季节更替用种类更替率A来分析[1];各季节鱼类物种相似性指数用Jaccard相似性指数Js表示[21]:

|
(5) |

|
(6) |
式中, C为相邻两季节间种类增加及减少数, N为两季节共有的种类数, a 、b分别为相邻季节的物种数, c为其共有物种数。
1.5 环境数据海洋环境数据测量按《海洋监测规范》(GB17378.5—2007)进行。由船载探测仪获得各站位水深, 萨氏盘测得海水透明度;由YSI Pro Series型多功能水质仪测得海水盐度、表底层水温、溶解氧和pH等物理环境数据;由荧光分光光度计测得叶绿素a值。
1.6 数据处理由于各年代间调查均采用底拖网, 网目差异不大, 主要是拖网时间有所差异, 为了年代间具有可比性, 在进行数据分析前对原始数据进行标准化处理, 每个航次各站位渔获质量和尾数均标准化成拖网时间1 h的渔获值。调查数据经标准化处理校正后, 用ArcGis 10.3软件和SPSS 19.0统计分析软件进行数据处理分析。各年代间优势种计算方法均采用相对重要性指数来评价(IRI>1000为优势种), 具有可比性。优势度、多样性指数、物种更替率计算方法均按照季节来计算, 比较季节间差异;全年多样性指数取各季节的平均值。利用相关检验和生物-环境分析研究影响鱼类群落物种组成及多样性的环境因子。其中, 生物—环境分析采用CANOCO 4.5软件中的典范对应分析(canonical correspondence analysis, CCA)进行分析处理数据, 排序结果用物种-环境因子关系的双序图来表示;单因子相似性分析(analysis of similarities, ANOSIM)用于不同季节群落物种组成及多样性差异的显著性检验。运用非线性多维标度排序(non-metric multidimensional scaling, NMDS)对4个季节鱼类群落结构进行显著性检验, 其中NMDS采用PRIMER5.0软件来进行操作;所得到的二维点图有一定解释意义;其中stress<0.1, 表明排序较好;stress<0.05, 表明排序有很好代表性[22]。
2 结果 2.1 种类组成捕获鱼类46种, 隶属2纲10目30科41属, 以硬骨鱼纲鱼类占绝对优势, 45种, 占总渔获种类数的97.83%;软骨鱼纲鱼类1种, 占总渔获种类数的2.17%。其中, 鲈形目(Perciformes)最多, 其次是鲉形目(Scorpaeniformes)和鲽形目(Pleuronectiformes);其余目种类数均不超过5种, 所占比例均小于10.00%。全年4个季节都出现的鱼类物种有1种, 为许氏平鮋(Sebastes schlegeli), 占种类总数的2.17%。各季节鱼类物种组成存在差异, 种、属和科所占比例均以夏季最高, 秋季最低, 春、冬季次之(表 1)。
| 季节 Season |
目Order | 种Species | 属Genus | 科Family | |||||||
| 数量Number | % | 数量Number | % | 数量Number | % | 数量Number | % | ||||
| 春季Spring | 4 | 40.00 | 18 | 39.13 | 17 | 41.46 | 14 | 46.67 | |||
| 夏季Summer | 6 | 60.00 | 23 | 50.00 | 21 | 51.22 | 16 | 53.33 | |||
| 秋季Autumn | 7 | 70.00 | 16 | 34.78 | 16 | 39.02 | 13 | 43.33 | |||
| 冬季Winter | 6 | 60.00 | 21 | 45.65 | 20 | 48.78 | 15 | 50.00 | |||
| 全年Annual | 10 | 46 | 41 | 30 | |||||||
胶州湾海域鱼类适温性组成以暖温性种占优势, 暖水性种次之, 冷温性种最少, 分别有22种、14种和10种, 占渔获总种类数的47.83%、30.43%和21.74%。季节变化上存在明显差异。春季, 暖温性和冷温性种类均最多, 8种;暖水性种类最少, 2种。夏季, 暖水性种类最多, 11种, 冷温性种类最少, 2种。秋季, 暖温性种类最多, 11种, 冷温性种类最少, 1种。冬季, 暖温性种类最多, 11种, 暖水性种类最少, 2种(图 2)。

|
| 图 2 胶州湾海域鱼类适温性组成 Fig. 2 The thermal nature composition of fishes in the Jiaozhou Bay |
胶州湾海域鱼类群落优势种组成存在明显的季节差异, 如表 2。
| 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | |||||||
| 种名Specie | IRI | 种名Specie | IRI | 种名Specie | IRI | 种名Specie | IRI | |||
| 褐牙鲆 | 3150 | 赤鼻棱鳀 | 8178 | 长丝鰕虎鱼 | 3490 | 褐菖鲉 | 5246 | |||
| Paralichthys olivaceus | Thryssa kammalensis | Cryptocentrus filifer | Sebastiscus marmoratus | |||||||
| 细纹狮子鱼 | 2488 | 长蛇鲻 | 1057 | 矛尾鰕虎鱼 | 3308 | 玉筋鱼 | 934 | |||
| Liparis tanakae | Saurida elongata | Chaeturichthys stigmatias | Ammodytes personatus | |||||||
| 方氏云鳚 | 1538 | 中颌棱鳀 | 876 | 尖海龙 | 1521 | 大泷六线鱼 | 924 | |||
| Enedras fangi Wang et Wang | Thryssa mystax | Syngnathus acus | Hexagrammos otakii | |||||||
| 星突江鲽 | 1041 | 白姑鱼 | 853 | 长蛇鲻 | 1202 | 六丝钝尾鰕虎鱼 | 316 | |||
| Platichthys stellatus | Argyrosomus argentatus | Amblychaeturichthys hexanema | Chaeturichthys hexanema | |||||||
| 矛尾鰕虎鱼 | 1007 | 细条天竺鲷 | 698 | 许氏平鮋 | 314 | 褐牙鲆 | 229 | |||
| Chaeturichthys stigmatias | Apogon lineatus | Sebastes schlegeli | Paralichthys olivaceus | |||||||
| 许氏平鮋 | 413 | 银鲳 | 600 | 黄鮟鱇 | 209 | 许氏平鲉 | 181 | |||
| Sebastes schlegeli | Pampus argenteus | Lophius litulon | Sebastes schlegeli | |||||||
| 鲱 | 345 | 褐菖鲉 | 302 | 花鲈 | 120 | 矛尾鰕虎鱼 | 120 | |||
| Callionymus beniteguri | Sebastiscus marmoratus | Lateolabrax maculatus | Chaeturichthys stigmatias | |||||||
| 纹缟鰕虎鱼 | 220 | 蓝点马鲛 | 187 | 褐牙鲆 | 109 | 鮻 | 106 | |||
| Tridentiger trigonocephalus | Scomberomorrus niphonius | Paralichthys olivaceus | Liza haematocheila | |||||||
| 大泷六线鱼 | 141 | 青鳞小沙丁 | 162 | 赤鼻棱鳀 | 107 | 孔鳐 | 103 | |||
| Hexagrammos otakii | Sardinella zunasi | Thryssa kammalensis | Raja porosa | |||||||
| 长丝鰕虎鱼 | 160 | |||||||||
| Cryptocentrus filifer | ||||||||||
| 大泷六线鱼 | 119 | |||||||||
| Hexagrammos otakii | ||||||||||
春季, 采集到鱼类18种。其中, 优势种5种, 分别为褐牙鲆(Paralichthys olivaceus)、细纹狮子鱼(Liparis tanakae)、方氏云鳚、星突江鲽(Platichthys stellatus)和矛尾鰕虎鱼(Chaeturichthys stigmatias), 占春季种类数的27.78%;重要种4种, 常见种9种, 分别占春季种类数的22.22%和50.00%。
夏季, 采集到鱼类23种。其中, 优势种2种, 分别为赤鼻棱鳀(Thryssa kammalensis)和长蛇鲻(Saurida elongata), 占夏季种类数的8.70%;重要种9种, 常见种8种, 一般种4种, 分别占夏季种类数的39.13%、34.78%和17.39%。
秋季, 采集到鱼类16种。其中, 优势种4种, 分别为长丝鰕虎鱼(Cryptocentrus filifer)、矛尾鰕虎鱼、尖海龙(Syngnathus acus)和长蛇鲻, 占秋季种类数的25.00%;重要种5种, 常见种5种, 一般种2种, 分别占秋季种类数的31.25%、31.25%和12.50%。
冬季, 采集到鱼类21种。其中, 优势种1种, 为褐菖鲉(Sebastiscus marmoratus), 占冬季种类数的4.76%;重要种8种, 常见种8种, 一般种4种, 分别占冬季种类数的38.10%、38.10%和19.04%。
2.3 物种多样性胶州湾海域4个季节的鱼类物种多样性如表 3所示:多样性指数(H′)变化范围为1.668—2.453, 以夏季最高, 春季最低;均匀度指数(J′)变化范围为0.577—0.808, 以秋季最高, 春季最低;丰富度指数(D′)变化范围为2.431—3.123, 以冬季最高, 秋季最低。
| 季节Season | 种类数Species | H′ | J′ | D′ |
| 春季Spring | 18 | 1.668 | 0.577 | 3.041 |
| 夏季Summer | 23 | 2.453 | 0.782 | 2.700 |
| 秋季Autumn | 16 | 2.241 | 0.808 | 2.431 |
| 冬季Winter | 21 | 1.763 | 0.579 | 3.123 |
| H′:多样性指数Diversity index;J′:均匀度指数Uniformity index;D′:丰富度指数Richness index | ||||
从空间分布来看(表 4), 各站位鱼类物种年均多样性指数H′范围为0.891—1.314。H′值最高的3个站是湾外的S8站和湾内的S5、S1站, 分别为1.238、1.314、1.116;H′值最低的3个站是湾内的S3、S7和湾外的S9站, 分别是0.891、0.915、0.902。年均均匀度指数J′范围为0.456—0.699;其中, S1、S3、S4和S5站均大于0.600, 其余各站小于0.600, 且各站位间相差不大。年均丰富度指数D′范围为0.717—1.716;D′值最高的3个站是湾内的S2、S7站和湾外的S8站, 分别为1.523、1.716、1.545;D′值最低的3个站是湾内的S3、S4站和湾外的S9站, 分别为1.037、1.072和0.717。
| 指数 Index |
时间 Time |
S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 |
| 多样性指数 | 春季 | 1.307 | 0.791 | — | 1.246 | 1.155 | 0.874 | 0.420 | — | 1.294 |
| Diversity index | 夏季 | 1.418 | 1.010 | 0.751 | 1.913 | 1.518 | — | 1.547 | 1.513 | 1.472 |
| 秋季 | 0.400 | 1.716 | 1.031 | 0.718 | 1.269 | 0.821 | 1.027 | 0.806 | 0.841 | |
| 冬季 | 1.337 | 0.793 | — | 0.633 | — | 1.196 | 0.666 | 1.396 | 0.000 | |
| 平均 | 1.116 | 1.078 | 0.891 | 1.128 | 1.314 | 0.964 | 0.915 | 1.238 | 0.902 | |
| 均匀度指数 | 春季 | 0.812 | 0.406 | — | 0.695 | 0.502 | 0.543 | 0.191 | — | 0.933 |
| Evenness index | 夏季 | 0.682 | 0.519 | 0.684 | 0.871 | 0.730 | — | 0.645 | 0.609 | 0.670 |
| 秋季 | 0.289 | 0.958 | 0.640 | 0.654 | 0.708 | 0.510 | 0.573 | 0.582 | 0.766 | |
| 冬季 | 0.746 | 0.408 | — | 0.576 | — | 0.743 | 0.414 | 0.606 | 0.000 | |
| 平均 | 0.632 | 0.573 | 0.662 | 0.699 | 0.647 | 0.599 | 0.456 | 0.599 | 0.592 | |
| 丰富度指数 | 春季 | 1.134 | 1.519 | — | 1.253 | 2.552 | 1.335 | 2.269 | — | 0.813 |
| Richness index | 夏季 | 1.898 | 1.519 | 0.690 | 1.510 | 0.889 | — | 2.612 | 1.935 | 1.531 |
| 秋季 | 1.038 | 1.669 | 1.384 | 0.410 | 1.003 | 1.610 | 1.185 | 1.038 | 0.522 | |
| 冬季 | 1.162 | 1.385 | — | 1.116 | — | 0.935 | 0.798 | 1.663 | 0.000 | |
| 平均 | 1.308 | 1.523 | 1.037 | 1.072 | 1.482 | 1.293 | 1.716 | 1.545 | 0.717 | |
| “—”表示未调查; S:站位Sampling | ||||||||||
胶州湾海域鱼类群落种类组成存在明显的季节更替现象。相似性分析(ANOSIM)表明, 各季节间鱼类种类组成存在极显著的差异性(P<0.01)。季节间鱼类物种相似性指数Js的变化范围为0.108—0.560, 以春—冬季最大, 春—夏季最小, 即:春冬季鱼类群落的相似性最高, 春夏季鱼类群落的相似性最低(表 5);季节更替率的变化范围为44.00%—89.19%, 尤其是春夏之交和夏秋之交更替率均较高, 鱼类物种组成更替比较显著。
| 季节Season | 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter |
| 春季Spring | 0.108 | 0.214 | 0.560 | |
| 夏季Summer | 0.147 | 0.128 | ||
| 秋季Autumn | 0.276 | |||
| 冬季Winter |
胶州湾海域各季节鱼类群落存在明显的差异性, 如图 3所示。其中, 春、秋、冬三季节可分为两大种群, 夏季分为三大种群。各季节胁强系数(stress)分别为0.08、0.04、0.07和0.01, 4个季节排序结果胁强系数均小于0.1, 即:排序结果具有一定解释意义且较好, 其中, 夏冬两季节排序结果具有很好的代表性。

|
| 图 3 胶州湾非参数多变量标序(NMDS)图 Fig. 3 Ordination of non-metric multidimensional scaling (NMDS) in Jiaozhou Bay |
鱼类群落物种与环境因子的CCA排序结果表明(图 4), 各季节环境因子对鱼类群落种类组成的影响作用存在显著差异(P<0.05), 环境因子与第一、二排序轴间的相关系数如表 6所示。从整体上看, 影响胶州湾鱼类群落种类组成的主要环境因素是温度、盐度、水深和pH值。春季, 第一、二排序轴物种与环境关系累积百分比为74.1%, pH、溶解氧(DO)和水深是影响鱼类群落种类组成的重要因子, 其相关系数分别为0.625, 0.100和-0.396。夏季, 第一、二排序轴线物种与环境关系累积百分比为62.6%, 温度、盐度和叶绿素a是影响鱼类群落种类组成的重要因子, 其相关系数分别为-0.392、0.270和-0.169。秋季, 第一、二排序轴物种与环境关系累积百分比为53.6%, pH和叶绿素a是影响鱼类群落种类组成的重要因子, 其相关系数分别为-0.557和-0.433。冬季, 第一、二排序轴物种与环境关系累积百分比为81.5%, 盐度, 叶绿素a和DO是影响鱼类群落种类组成的重要因子, 其相关系数分别为-0.998、0.988和0.808。

|
| 图 4 胶州湾鱼类群落物种-环境因子典范对应分析排序图 Fig. 4 Ordination diagram of canonical correspondence analysis (CCA) based on fish community species and environmental factors in the Jiaozhou Bay area Enedr:方氏云鳚Enedras fangi Wang et Wang;Enche:长绵鳚Enchelyopus elongatus;Sebas:许氏平鮋Sebastes schlegeli;Tride:纹缟鰕虎鱼Tridentiger trigonocephalus;Chaet-s:矛尾鰕虎鱼Chaeturichthys stigmatias;Platy:鲬Platycephalus indicus;Paral:褐牙鲆Paralichthys olivaceus;Hexag:大泷六线鱼Hexagrammos otakii;Triae:钟馗鰕虎鱼Triaenopogon barbatus;Ambly:六丝钝尾鰕虎鱼Amblychaeturichthys hexanema;Karei:石鲽Kareius bicoloratus;Argyr:白姑鱼Argyrosomus argentatus;Crypt:长丝鰕虎鱼Cryptocentrus filifer;Pampu:银鲳Pampus argenteus;Odont:红狼牙鰕虎鱼Odontamblyopus rubicundus;Cteno:中华栉孔鰕虎鱼Ctenotrypauchen chinensis;Ammod:玉筋鱼Ammodytes personatus;Raja:孔鳐Raja porosa;Scomb:蓝点马鲛Scomberomorrus niphonius;Calli-v:瓦氏Callionymus valenciennei;Upene:条尾绯鲤Upeneus bensasi;Konos:斑鰶Konosirus punctatus;Scomb:日本鲭Scomber japonicus;Sphyr:油妤Sphyraena pinguis;Liza:鮻Liza haematocheila;Apogo:细条天竺鲷Apogon lineatus;Sardi:青鳞小沙丁Sardinella zunasi;Hippo:日本海马Hippocampus japonicus;Cynog:长吻红舌鳎Cynoglossus lighti;Acent:普氏栉虾虎鱼Acentrogobius pflaumii;Thrys:中颌棱鳀Thryssa mystax;Sebas:褐菖鲉Sebastiscus marmoratus;Lateo:花鲈Lateolabrax maculatus;Apogo:细条天竺鱼Apogonichthys lineatus;Conge:星康吉鳗Conger myriaster;Lophi:黄鮟鱇Lophius litulon;Callio-b:鲱Callionymus beniteguri;Thrys-k:赤鼻棱鳀Thryssa kammalensis;Minou:虎鲉Minous monodactylus;Lipar:细纹狮子鱼Liparis tanakae;Plati:星突江鲽Platichthys stellatus;Syngn:尖海龙Syngnathus acus;Cheli:绿鳍鱼Chelidonichthys kumu;Veras:圆斑星鲽Verasper variegatus;Cynog:短吻红舌鳎Cynoglossus joyneri;Sauri:长蛇鲻Saurida elongata |
| 环境因子 Environmental factors |
春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | |||||||
| CCA1 | CCA2 | CCA1 | CCA2 | CCA1 | CCA2 | CCA1 | CCA2 | ||||
| 水深Depth | -0.396 | 0.334 | -0.159 | 0.534 | -0.367 | 0.329 | -0.760 | 0.190 | |||
| 水温Temperature | 0.038 | -0.446 | -0.392 | -0.449 | 0.026 | 0.500 | -0.787 | 0.175 | |||
| 盐度Salinity | 0.183 | 0.162 | 0.270 | 0.427 | 0.103 | 0.410 | -0.998 | 0.005 | |||
| 叶绿素a Chlorophyll a | -0.063 | -0.315 | -0.169 | -0.548 | -0.433 | -0.409 | 0.988 | 0.102 | |||
| 酸碱度pH | 0.625 | -0.313 | 0.138 | 0.450 | -0.557 | -0.001 | 0.570 | 0.243 | |||
| 溶解氧Dissolved oxygen | 0.100 | 0.540 | -0.045 | 0.033 | -0.379 | -0.043 | 0.808 | 0.364 | |||
| CCA:典范对应分析canonical correspondence analysis | |||||||||||
物种多样性的差异性与鱼类生存环境[23]、洄游特性[24]、繁殖和人为捕捞密切相关[25], 且水深对鱼类分布具有限制作用[26]。胶州湾鱼类多样性与环境因子相关性表明(表 7), 水温和pH对该海域物种多样性具有极显著相关性(P<0.01), 其他环境因子则无明显的相关性(P>0.05)。均匀度指数J′的季节变化不大, 说明鱼类群落的均匀度受生境条件改变的影响较小。夏季, 水温和盐度均最高(27.03℃, 29.74), pH值最低(7.88), 营养盐丰富, 浮游动植物大量繁殖为鱼类带来丰富营养饵料, 促进鱼类繁衍和生长;此外, 春、夏季为胶州湾鱼类主要生殖季节[1], 且6—8月为黄海休渔期, 鱼卵和幼鱼等鱼类资源得到保护;加上本次调查是在禁渔期刚结束后就展开调查, 受捕捞影响较小, 致使渔获种类最多(23种), 个体数量最多(3460尾), 生物量较大(12.24 kg), 物种多样性指数最大。冬季, 水温较低(5.50℃), 捕捞压力较小, 且受季节洄游影响, 暖水性和暖温性季节洄游鱼类游向湾外越冬, 种类数减少(21种), 鱼类群落组成主要以地域性的暖温性和冷温性鱼类为主(表 2), 但由于渔获生物量最大(23.62 kg), 渔获个体数量相对较少(604尾), 致使鱼类丰富度最大, 物种多样性相对较低。此外, 海域底质类型也影响鱼类多样性, 且黏土底质对胶州湾鱼类群落的分布具有显著影响[6]。
| 环境因子 Environmental factors |
H′ | J′ | D′ | |||||
| r | P | r | P | r | P | |||
| 水深Depth | -0.107 | 0.626 | -0.251 | 0.248 | -0.051 | 0.817 | ||
| 水温Temperature | 0.540 | 0.008** | 0.371 | 0.081 | 0.211 | 0.334 | ||
| 盐度Salinity | 0.020 | 0.926 | -0.078 | 0.723 | 0.077 | 0.728 | ||
| 叶绿素a Chlorophyll a | -0.134 | 0.543 | 0.082 | 0.711 | -0.104 | 0.637 | ||
| 酸碱度pH | -0.581 | 0.004** | -0.352 | 0.100 | -0.202 | 0.354 | ||
| 溶解氧Dissolved oxygen | -0.118 | 0.591 | -0.188 | 0.389 | 0.051 | 0.819 | ||
| r表示环境因子与物种多样性的相关系数;*:P<0.05, 表示显著差异;**:P<0.01, 表示极显著差异 | ||||||||
本次调查与1981—1982年、2003—2004年、2011年航次相比, 种类数呈显著下降趋势, 分别下降了59.29%、20.69%和19.30%;优势种由1981—1982年以青鳞小沙丁和斑鰶为主[1], 转变为2011年以方氏云鳚和六丝钝尾鰕虎鱼为主[6], 演变到本次以赤鼻棱鳀、褐菖鲉和褐牙鲆为主, 更替显著, 且暖温性种占优势(表 8)。鱼类多样性水平低于2003—2004年和2011年, 呈明显的下降趋势(图 5)。可见, 胶州湾海域鱼类群落种类数及优势种组成发生变化。同时, 各年代优势种的季节变化也存在明显的更替现象, 表 9。1981—1982年, 平均更替率为41.60%, 其中冬春季更替率最大(61.50%), 夏秋季更替率最小(35.00%)[1];2011年, 平均更替率为54.69%, 其中春夏季更替率最大(64.58%), 夏秋季更替率最小(42.86%)[6];2016—2017, 平均更替率为72.72%, 其中春夏季更替率最大(89.19%), 冬春更替率最小(44.00%);可知, 胶州湾鱼类物种更替率呈逐步增加趋势, 季节更替率也发生明显变化。此外, 主要大型经济鱼类物种减少。如鲈鱼(Lateolabrax maculatus)体长范围、质量百分比和尾数百分比分别由1981—1982年的100—600 mm、2.00%、0.83%减少到2016—2017年的180—290 mm、1.57%、0.08%;银鲳(Pampus argenteus)体长范围、质量百分比和尾数百分比分别由1981—1982年的100—300 mm、4.00%、0.83%减少到2016—2017年的95—145 mm、2.66%、0.67%;长绵鳚(Enchelyopus elongatus)体长范围、质量百分比和尾数百分比分别由1981—1982年的150—400 mm、6.00%、0.83%减少到2016—2017年的295 mm、0.52%、0.04%;带鱼体长范围、质量百分比和尾数百分比分别由1981—1982年的80—300 mm、4.00%、3.00%减少到2016—2017年的未捕捞到一尾。此外, 鱼类资源量也发生明显变化。本次调查鱼类平均资源密度(1593.16 kg/km2)低于1981—1982年鱼类平均资源密度(10857 kg/km2), 约下降了85.33%。
| 年份 Year |
种类数 Species |
优势种 Dominant species |
适温性 Moderate temperature |
| 1981—1982 | 113 | 青鳞小沙丁鱼(Sardinella zunasi) | 暖温种 |
| 斑鰶(Konosirus punctatus) | 暖温种 | ||
| 赤鼻棱鳀(Thryssa kammalensis) | 暖温种 | ||
| 中颌棱鳀(Thryssa mystax) | 暖水种 | ||
| 细条天竺鲷(Apogon lineatus) | 暖水种 | ||
| 带鱼(Trichiurus lepturus) | 暖水种 | ||
| 2003—2004 | 58 | 小黄鱼(Larimichthys polyactis) | 暖温种 |
| 皮氏叫姑鱼(Johnius belangerii) | 暖水种 | ||
| 玉筋鱼(Ammodytes personatus) | 冷温种 | ||
| 方氏云鳚(Enedras fangi Wang et Wang) | 冷温种 | ||
| 2011 | 57 | 方氏云鳚(Enedras fangi Wang et Wang) | 冷温种 |
| 六丝钝尾鰕虎鱼(Amblychaeturichthys hexanema) | 暖温种 | ||
| 2016—2017 | 46 | 赤鼻棱鳀(Thryssa kammalensis) | 暖温种 |
| 褐菖鲉(Sebastiscus marmoratus) | 暖温种 | ||
| 褐牙鲆(Paralichthys olivaceus) | 暖温种 |

|
| 图 5 胶州湾海域鱼类物种多样性指数的年际变化 Fig. 5 Inter-annual variability of fish species diversity index in the Jiaozhou Bay H′:多样性指数Diversity index;J′:均匀度指数Uniformity index;D′:丰富度指数Richness index |
| 年份Year | 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter |
| 1981—1982 | 牙鲆 | 绿鳍马面魨 | 斑鰶 | 梭鱼 |
| Paralichihys olivaceus | Thamnaconus modestus | Konosirus punctatus | Liza so-iuy | |
| 青鳞小沙丁鱼 | 长蛇鲻 | 长绵鳚 | ||
| Sardinella zunasi | Saurida elongala | Enchelyopus elongaius | ||
| 2003—2004 | 方氏云鳚 | 小黄鱼 | 方氏云鳚 | |
| Enedras fangi Wang et Wang | Larimichthys polyactis | Enedras fangi Wang et Wang | ||
| 玉筋鱼 | 皮氏叫姑鱼 | 玉筋鱼 | ||
| Ammodytes personatus | Johnius belangerii | Ammodytes personatus | ||
| 六丝钝尾鰕虎鱼 | ||||
| Amblychaeturichthys hexanema | ||||
| 2011 | 方氏云鳚 | 赤鼻棱鳀 | 六丝钝尾鰕虎鱼 | 方氏云鳚 |
| Enedras fangi Wang et Wang | Thryssa kammalensis | Amblychaeturichthys hexanema | Enedras fangi Wang et Wang | |
| 六丝钝尾鰕虎鱼 | 皮氏叫姑鱼 | 普氏缰鰕虎鱼 | 李氏 | |
| Amblychaeturichthys hexanema | Johnius belangerii | Acentrogobius pflaumii | Repomucenus richardsonii | |
| 细纹狮子鱼 | 斑鰶 | 方氏云鳚 | 六丝钝尾鰕虎鱼 | |
| Liparis tanakae | Konosirus punctatus | Enedras fangi Wang et Wang | Amblychaeturichthys hexanema | |
| 尖海龙 | 细条天竺鲷 | |||
| Syngnathus acus | Apogon lineatus | |||
| 长吻红舌鰨 | ||||
| Cynoglossus lighti | ||||
| 2016—2017 | 褐牙鲆 | 赤鼻棱鳀 | 长丝鰕虎鱼 | 褐菖鲉 |
| Paralichthys olivaceus | Thryssa kammalensis | Cryptocentrus filifer | Sebastiscus marmoratus | |
| 细纹狮子鱼 | 长蛇鲻 | 矛尾鰕虎鱼 | ||
| Liparis tanakae | Saurida elongala | Chaeturichthys stigmatias | ||
| 方氏云鳚 | 尖海龙 | |||
| Enedras fangi Wang et Wang | Syngnathus acus | |||
| 星突江鲽 | ||||
| Platichthys stellatus | ||||
| 矛尾鰕虎鱼 | ||||
| Chaeturichthys stigmatias |
30多年来, 受人类活动及自然扰动的影响, 胶州湾鱼类种类数减少, 物种多样性下降, 优势种更替显著, 鱼类群落结构趋向简单化。主要原因有以下几个方面:第一, 受人类捕捞影响, 尤其是选择性捕捞, 导致个体质量大的经济鱼类减少、多样性降低, 鱼类组成以个体质量小、未成熟的低值鱼类为主[27]。据统计, 2000—2016年胶州湾鱼类资源捕捞量下降了74.73%[28], 说明受人类捕捞影响, 鱼类资源量衰竭, 尤其是优质鱼类剧减, 鱼类群落结构发生改变。第二, 菲律宾蛤仔等贝类的大规模养殖。据统计, 贝类养殖面积占胶州湾总养殖面积的71.6%, 其中菲律宾蛤仔养殖面积占贝类养殖面积的93.2%、全湾养殖面积的66.7%[29];1994—2008年菲律宾蛤仔养殖面积和产量分别增加了约95%和99.63%[30], 近年来养殖产量已达30多万吨, 且菲律宾蛤仔为底栖动物主要优势种[31], 其生物扰动影响营养盐在沉积物-水界面的交换, 加剧水体中Si限制[32], 大规模养殖造成鱼类栖息地环境和底栖鱼类底质的破坏, 影响鱼类产卵繁育、底层鱼类群落结构以及定居种的生长[7]。第三, 鱼类栖息地遭到破坏。围填海和炸山等人类活动破坏了胶州湾海底地形和沿岸地貌, 且围填海使胶州湾海域面积不断缩小, 与1863年相比缩小了38.60%, 导致海湾纳潮量降低, 海洋自净能力降低, “赤潮”和“浒苔”频发, 致使栖息地受到破坏, 渔业资源衰退, 多样性水平降低[33]。第四, 海域生态环境破坏严重。沿岸生活污水及工业废水大量排入, 加上船舶溢油, 湾内中度污染和轻度污染海域约占胶州湾总面积的3/5, 富营养化污染加剧, 水质质量下降, 生物多样性减少, 鱼类群落结构发生改变[34]。
今后应加大胶州湾海域生态环境的监测与保护, 做到生态保护和资源开发的可持续发展。首先, 实施生态修复工程。政府应积极开展实施退田还海、滩涂岸线整治等生态修复建设工程, 加强滨海湿地养护、入湾河口修复, 在滩涂种植碱蓬、柽柳等耐盐碱植物进行生态修复, 恢复原始自然风貌, 研发新的生态补偿管理技术, 为胶州湾生态修复提供技术支撑;其次, 实施渔业资源保育工程。如完善休渔管理制度, 优化改革捕捞方式及网具类型, 实施限额捕捞, 开展增殖放流, 促进渔业资源的恢复和生态系统的平衡;此外, 应该在坚持“生态优先”的理念下, 建设海洋牧场, 开展海上观光旅游、垂钓、海底潜水采捕等休闲渔业为代表的海洋第三产业, 实现三产融合。
致谢: 中国科学院海洋研究所海洋生态与环境科学重点实验室宋金明研究员、袁华茂博士、中国水产科学研究院黄海水产研究所袁伟、孙坚强老师等帮助采集样品;中国海洋大学水产学院张弛老师帮助标本鉴定,特此致谢。| [1] |
刘瑞玉. 胶州湾生态学和生物资源. 北京: 科学出版社, 1992: 271-295.
|
| [2] |
李京梅, 刘铁鹰. 基于生境等价分析法的胶州湾围填海造地生态损害评估. 生态学报, 2012, 32(22): 7146-7155. |
| [3] |
杨东方, 高振会, 马媛, 孙培艳, 杨应斌. 胶州湾环境变化对海洋生物资源的影响. 海洋环境科学, 2006, 25(4): 39-42. DOI:10.3969/j.issn.1007-6336.2006.04.011 |
| [4] |
Xue Y, Ren Y, Xu B, Mei C, Chen X, Zan X. Length-weight relationships of fish species caught by bottom trawl in Jiaozhou Bay, China. Journal of Applied Ichthyology, 2011, 27(3): 949-954. DOI:10.1111/j.1439-0426.2010.01607.x |
| [5] |
曾晓起, 朴成华, 姜伟, 刘群. 胶州湾及其邻近水域渔业生物多样性的调查研究. 中国海洋大学学报, 2004, 34(6): 977-982. |
| [6] |
翟璐, 韩东燕, 傅道军, 张崇良, 薛莹. 胶州湾及其邻近海域鱼类群落结构及与环境因子的关系. 中国水产科学, 2014, 21(4): 810-821. |
| [7] |
徐宾铎, 曾慧慧, 薛莹, 纪毓鹏, 任一平. 胶州湾近岸浅水区鱼类群落结构及多样性. 生态学报, 2013, 33(10): 3074-3082. |
| [8] |
高权新, 谢明媚, 彭士明, 张晨捷, 施兆鸿. 急性温度胁迫对银鲳幼鱼代谢酶、离子酶活性及血清离子浓度的影响. 南方水产科学, 2016, 12(2): 59-66. DOI:10.3969/j.issn.2095-0780.2016.02.009 |
| [9] |
韩东燕, 薛莹, 纪毓鹏, 麻秋云. 胶州湾六丝钝尾虾虎鱼的摄食生态特征. 应用生态学报, 2013, 24(5): 1446-1452. |
| [10] |
韩真, 肖永双, 高天翔. 中国近海9个小黄鱼群体的形态学比较研究. 南方水产科学, 2012, 8(3): 25-33. DOI:10.3969/j.issn.2095-0780.2012.03.004 |
| [11] |
马立杰, 杨曦光, 祁雅莉, 刘艳霞, 张金枝. 胶州湾海域面积变化及原因探讨. 地理科学, 2014, 34(3): 365-369. |
| [12] |
马孟磊, 陈作志, 许友伟, 张魁, 袁伟, 徐姗楠. 基于Ecopath模型的胶州湾生态系统结构和能量流动分析. 生态学杂志, 2018, 37(2): 462-470. |
| [13] |
Day J J. Fishes of the world, 4th Edition. Fish and Fisheries, 2006, 7(4): 334.
|
| [14] |
李明德. 鱼类分类学. 北京: 海洋出版社, 1998: 28-47.
|
| [15] |
农业部水产局, 农业部黄海区渔业指挥部. 黄、渤海区渔业资源调查和区划. 北京: 海洋出版社, 1990.
|
| [16] |
张春霖. 黄渤海鱼类调查报告. 北京: 科学出版社, 1955.
|
| [17] |
Pinkas L, Iverson I L K. Food habits of albacore, bluefin tuna and bonito in California waters. California Department of Fish and Game:Fish Bulletin, 1971, 152: 1-105. |
| [18] |
王雪辉, 杜飞雁, 邱永松, 李纯厚, 孙典荣, 贾晓平. 1980-2007年大亚湾鱼类物种多样性、区系特征和数量变化. 应用生态学报, 2010, 21(9): 2403-2410. |
| [19] |
Ludwig J A, Reynolds J F. Statistical Ecology: A Primer in Methods and Computing. New York: Wiley-Interscience, 1988.
|
| [20] |
Wilhm J L. Use of biomass units in Shannon's formula. Ecology, 1968, 49(1): 153-156. DOI:10.2307/1933573 |
| [21] |
Jaccard P. The distribution of the flora in the alpine zone 1. New Phytologist, 1912, 11(2): 37-50. DOI:10.1111/j.1469-8137.1912.tb05611.x |
| [22] |
Khalaf M A, Kochzius M. Changes in trophic community structure of shore fishes at an industrial site in the Gulf of Aqaba, Red Sea. Marine Ecology Progress Series, 2002, 239: 287-299. DOI:10.3354/meps239287 |
| [23] |
袁梦, 汤勇, 徐姗楠, 陈作志, 杨玉滔, 江艳娥. 珠江口南沙海域秋季渔业资源群落结构特征. 南方水产科学, 2017, 13(2): 18-25. DOI:10.3969/j.issn.2095-0780.2017.02.003 |
| [24] |
袁华荣, 陈丕茂, 秦传新, 黎小国, 周艳波, 冯雪, 余景, 舒黎明, 唐振朝, 佟飞. 南海柘林湾鱼类群落结构季节变动的研究. 南方水产科学, 2017, 13(2): 26-35. DOI:10.3969/j.issn.2095-0780.2017.02.004 |
| [25] |
李圣法, 程家骅, 李长松, 李建生. 东海中部鱼类群落多样性的季节变化. 海洋渔业, 2005, 27(2): 113-119. DOI:10.3969/j.issn.1004-2490.2005.02.005 |
| [26] |
武智, 李新辉, 李捷, 陈方灿, 朱书礼. 红水河岩滩水库鱼类资源声学评估. 南方水产科学, 2017, 13(3): 20-25. DOI:10.3969/j.issn.2095-0780.2017.03.003 |
| [27] |
Fenberg P B, Roy K. Ecological and evolutionary consequences of size-selective harvesting:how much do we know?, , 17(1):209-220. Molecular Ecology, 2008, 17(1): 209-220. DOI:10.1111/j.1365-294X.2007.03522.x |
| [28] |
刘岐涛. 2017青岛统计年鉴. 北京: 中国统计出版社, 2017.
|
| [29] |
张明亮.胶州湾贝类养殖容量研究与分析[D].青岛: 国家海洋局第一海洋研究所, 2008. http://cdmd.cnki.com.cn/Article/CDMD-85301-2008180320.htm
|
| [30] |
郭永禄.胶州湾底播增殖菲律宾蛤仔(Ruditapes philippinarum)渔业生物学研究[D].青岛: 中国海洋大学, 2005. http://cdmd.cnki.com.cn/Article/CDMD-10423-2005139623.htm
|
| [31] |
于海燕, 李新正, 李宝泉, 王金宝, 王洪法. 胶州湾大型底栖动物生物多样性现状. 生态学报, 2006, 26(2): 416-422. DOI:10.3321/j.issn:1000-0933.2006.02.014 |
| [32] |
邓可, 刘素美, 张桂玲, 陆小兰, 张经. 菲律宾蛤仔养殖对胶州湾沉积物-水界面生源要素迁移的影响. 环境科学, 2012, 33(3): 782-793. |
| [33] |
郭臣.胶州湾围填海造陆生态补偿机制研究[D].青岛: 中国海洋大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10423-1012505599.htm
|
| [34] |
张学庆, 刘津梁, 王翠. 胶州湾随机动力条件下的船舶溢油污染概率研究. 应用海洋学学报, 2014, 33(3): 379-384. DOI:10.3969/J.ISSN.2095-4972.2014.03.013 |